Твердые органические вещества

| Часть серии о |
| Углеродный цикл |
|---|
 |
Твердые органические вещества (ПОМ) — это часть общего органического вещества, определяемая с практической точки зрения как та, которая не проходит через поры фильтра, размер которых обычно варьируется от 0,053 миллиметра (53 мкм) до 2 миллиметров. [3]
Частичный органический углерод (POC) – это тесно связанный термин, который часто используется как синоним POM. POC относится конкретно к массе углерода в твердых частицах органического материала, тогда как POM относится к общей массе твердых частиц органического вещества. Помимо углерода, ПОМ включает в себя массу других элементов органического вещества, таких как азот, кислород и водород. В этом смысле ПОК является компонентом ПОМ, и его обычно примерно в два раза больше, чем ПОК. [4] Многие утверждения, которые можно сделать о POM, в равной степени применимы и к POC, и многое из того, что сказано в этой статье о POM, в равной степени можно было бы сказать и о POC.
Твердые органические вещества иногда называют взвешенными органическими веществами, макроорганическими веществами или органическими веществами крупной фракции. При выделении проб земли путем просеивания или фильтрации в эту фракцию входят частично разложившийся детрит и растительный материал, пыльца и другие материалы. [5] [6] При просеивании для определения содержания ПОМ консистенция имеет решающее значение, поскольку размер отдельных фракций будет зависеть от силы перемешивания. [7]
ПОМ легко разлагается, выполняет множество функций почвы и поставляет наземный материал в водоемы. Он является источником питания как для почвенных , так и для водных организмов и обеспечивает питательными веществами растения . В водоемах ПОМ может существенно способствовать повышению мутности, ограничивая фотную глубину, что может подавлять первичную продуктивность. ПОМ также улучшает структуру почвы , что приводит к увеличению инфильтрации воды , аэрации и устойчивости к эрозии . [5] [8] Практика управления почвой , такая как обработка почвы и внесение компоста / навоза , изменяет содержание ПОВ в почве и воде. [5] [6]
Обзор
[ редактировать ]Твердый органический углерод (POC) с практической точки зрения определяется как весь горючий некарбонатный углерод , который может быть собран фильтром . Океанографическое сообщество исторически использовало различные фильтры и размеры пор, чаще всего 0,7, 0,8 или 1,0 мкм фильтры из стекла или кварцевого волокна с размером пор . Биомасса . живого зоопланктона намеренно исключается из ПОУ путем использования предварительного фильтра или специально сконструированных приемников проб, отпугивающих плавающие организмы [9] Субмикронные частицы, включая большинство морских прокариот , диаметром 0,2–0,8 мкм, часто не улавливаются, но их следует рассматривать как часть ПОУ, а не растворенного органического углерода (РОУ), размер которого обычно определяется как <0,2 мкм. [10] [9] Обычно считается, что ПОУ содержит взвешенные и тонущие частицы размером ≥ 0,2 мкм, что, следовательно, включает биомассу живых микробных клеток, детритный материал, включая мертвые клетки, фекальные гранулы , другой агрегированный материал и органическое вещество земного происхождения. В некоторых исследованиях POC дополнительно разделяется по оперативному принципу в зависимости от скорости или размера его погружения. [11] с частицами размером ≥ 51 мкм, иногда приравниваемыми к тонущей фракции. [12] И DOC, и POC играют важную роль в углеродном цикле , но POC является основным путем, по которому органический углерод , производимый фитопланктоном, экспортируется – в основном путем гравитационного осаждения – с поверхности в глубокие слои океана и, в конечном итоге, в отложения , и, таким образом, является ключевым Компонент биологического насоса . [13] [14] [15] [16] [17] [9]
Наземные экосистемы
[ редактировать ]Органическое вещество почвы
[ редактировать ]Органическое вещество почвы – это все, что находится в почве биологического происхождения. Углерод является его ключевым компонентом, составляющим около 58% по весу. Простую оценку общего органического вещества можно получить путем измерения органического углерода в почве. Живые организмы (включая корни) составляют около 15% от общего количества органического вещества в почве. Они имеют решающее значение для функционирования цикла углерода в почве . Далее речь идет об остальных 85% органического вещества почвы – неживой составляющей. [18]
Как показано ниже, неживые органические вещества в почвах можно разделить на четыре отдельные категории на основе размера, поведения и стойкости. [19] Эти категории расположены в порядке убывания способности к разложению. Каждый из них по-разному способствует здоровью почвы. [19] [18]
| Органическое вещество почвы |
| ||||||||||||||||||
| (неживой) |
Растворенное органическое вещество (РОВ): органическое вещество, растворяющееся в почвенной воде. Он состоит из относительно простых органических соединений (например, органических кислот, сахаров и аминокислот), которые легко разлагаются. Срок оборота менее 12 месяцев. Сюда включены экссудаты корней растений (слизи и десны). [18]
Частичное органическое вещество (POM): органическое вещество, сохраняющее свою первоначальную клеточную структуру. [18] и обсуждается далее в следующем разделе.
Гумус : обычно представляет собой наибольшую долю органических веществ в почве, его доля составляет от 45 до 75%. Обычно он прилипает к почвенным минералам и играет важную роль в структурировании почвы. Гумус является конечным продуктом деятельности почвенных организмов, он химически сложен и не имеет узнаваемых характеристик своего происхождения. Гумус имеет очень маленькие размеры и большую площадь поверхности по отношению к его весу. Он удерживает питательные вещества, обладает высокой водоудерживающей способностью и значительной катионообменной способностью , буферизует изменение pH и может удерживать катионы. Гумус разлагается довольно медленно и существует в почве десятилетиями. [18]
Устойчивое органическое вещество: имеет высокое содержание углерода и включает древесный уголь, обугленные растительные материалы, графит и уголь. Сроки оборота велики и исчисляются сотнями лет. Он не является биологически активным, но положительно влияет на структурные свойства почвы, включая водоудерживающую способность, катионообменную способность и термические свойства. [18]
Роль ПОМ в почвах
[ редактировать ]Твердые органические вещества (ПОВ) включают в себя устойчиво разлагающийся растительный опад и фекалии животных, а также детрит, образующийся в результате деятельности микроорганизмов. Большая часть его постоянно подвергается разложению микроорганизмами (при достаточной влажности) и обычно имеет срок годности менее 10 лет. Для оборота менее активных частей может потребоваться от 15 до 100 лет. Там, где частицы органического вещества все еще находятся на поверхности почвы и относительно свежие, они перехватывают энергию дождевых капель и защищают физические поверхности почвы от повреждений. В процессе разложения твердые органические вещества обеспечивают большую часть энергии, необходимой почвенным организмам, а также обеспечивают устойчивое выделение питательных веществ в почвенную среду. [18]
Разложение ПОМ обеспечивает энергию и питательные вещества. Питательные вещества, не усваиваемые почвенными организмами, могут быть доступны для потребления растениями. [6] Количество питательных веществ, высвобождаемых ( минерализованных ) при разложении, зависит от биологических и химических характеристик ПОМ, таких как соотношение C:N . [6] Помимо высвобождения питательных веществ, редуценты, колонизирующие ПОМ, играют роль в улучшении структуры почвы. [20] Грибной мицелий опутывает частицы почвы и выделяет в почву липкие, похожие на цемент полисахариды; в конечном итоге образуя почвенные агрегаты [20]
На содержание РОМ в почве влияют органические вещества и деятельность разлагателей почвы. Добавление органических материалов, таких как навоз или растительные остатки , обычно приводит к увеличению содержания ПОВ. [6] Альтернативно, повторная обработка или нарушение почвы увеличивает скорость разложения, подвергая почвенные организмы воздействию кислорода и органических субстратов ; в конечном итоге, истощение ПОМ. Снижение содержания ПОМ наблюдается при преобразовании естественных лугов в сельскохозяйственные угодья. [5] Температура и влажность почвы также влияют на скорость разложения ПОМ. [6] Поскольку ПОМ является легкодоступным (лабильным) источником питательных веществ в почве, вносит вклад в структуру почвы и очень чувствителен к управлению почвой, его часто используют в качестве индикатора для измерения качества почвы . [8]
Пресноводные экосистемы
[ редактировать ]На плохо обрабатываемых почвах, особенно на склонах, эрозия и перенос почвенных отложений, богатых ПОМ, могут привести к загрязнению водоемов. [8] Поскольку ПОМ является источником энергии и питательных веществ, быстрое накопление органических веществ в воде может привести к эвтрофикации . [20] Взвешенные органические материалы также могут служить потенциальным переносчиком загрязнения воды фекальными бактериями , токсичными металлами или органическими соединениями.
Морские экосистемы
[ редактировать ]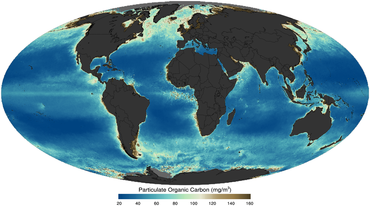
как снято со спутника в 2011 году


Жизнь и твердые органические вещества в океане фундаментально сформировали планету. На самом базовом уровне твердые органические вещества можно определить как живые, так и неживые вещества биологического происхождения размером ≥0,2 мкм в диаметре, включая все, что угодно, от небольших бактерий (размером 0,2 мкм) до синих китов (20 м размером). [22] Органическое вещество играет решающую роль в регулировании глобальных морских биогеохимических циклов и событий, начиная с Великого события окисления в ранней истории Земли. [23] к улавливанию атмосферного углекислого газа в глубинах океана. [24] Таким образом, понимание распределения, характеристик, динамики и изменений во времени твердых частиц в океане имеет основополагающее значение для понимания и прогнозирования морской экосистемы, от динамики пищевой сети до глобальных биогеохимических циклов. [25] [26]
Измерение ПОМ
[ редактировать ]Оптические измерения частиц становятся важным методом для понимания углеродного цикла океана, включая вклад в оценку их нисходящего потока, который связывает углекислый газ в глубоком море. Оптические инструменты можно использовать с кораблей или устанавливать на автономных платформах, обеспечивая гораздо больший пространственный и временной охват частиц в мезопелагической зоне океана, чем традиционные методы, такие как отстойники . Технологии получения изображений частиц значительно продвинулись за последние два десятилетия, но количественный перевод этих огромных наборов данных в биогеохимические свойства остается проблемой. В частности, необходимы достижения для обеспечения оптимального перевода отображаемых объектов в содержание углерода и скорости погружения. Кроме того, разные устройства часто измеряют разные оптические свойства, что приводит к трудностям при сравнении результатов. [25]
Первичная продукция океана
[ редактировать ]Первичную морскую продукцию можно разделить на новую продукцию из аллохтонных питательных веществ, поступивших в эвфотическую зону , и регенерированную продукцию из рециркуляции питательных веществ в поверхностных водах. Общая новая продукция в океане примерно равна опускающемуся потоку твердых частиц органического вещества в глубины океана, примерно 4 × 10 9 тонн углерода ежегодно. [27]
Модель тонущих океанических частиц
[ редактировать ]Тонущие океанические частицы имеют широкий диапазон формы, пористости, балласта и других характеристик. Модель, показанная на диаграмме справа, пытается уловить некоторые из преобладающих особенностей, которые влияют на форму профиля опускающегося потока (красная линия). [21] Опускание органических частиц, образующихся в верхних освещенных солнцем слоях океана, образует важную часть океанического биологического насоса, который влияет на секвестрацию углерода и пополнение запасов питательных веществ в мезопелагическом океане. Частицы, выпадающие из верхних слоев океана, подвергаются реминерализации бактериями, колонизированными на их поверхности и внутри, что приводит к ослаблению опускающегося потока органического вещества с глубиной. Диаграмма иллюстрирует механистическую модель зависящего от глубины потока массы тонущих частиц, состоящего из ряда тонущих реминерализующих частиц. [21]
Морской снег различается по форме, размеру и характеру: от отдельных клеток до комков и агрегатов, большая часть которых быстро колонизируется и потребляется гетеротрофными бактериями, что способствует ослаблению опускающегося потока с глубиной. [21]
Скорость погружения
[ редактировать ]Диапазон зарегистрированных скоростей погружения частиц в океанах варьируется от отрицательных (частицы всплывают к поверхности) [28] [29] до нескольких км в день (как в случае сальповых фекальных гранул) [30] При рассмотрении скорости погружения отдельной частицы первое приближение можно получить из закона Стокса (первоначально полученного для сферических, непористых частиц и ламинарного потока) в сочетании с приближением Уайта: [31] которые предполагают, что скорость погружения увеличивается линейно с увеличением избыточной плотности (отличия от плотности воды) и квадрата диаметра частиц (т.е. линейно с площадью частиц). Основываясь на этих ожиданиях, многие исследования пытались связать скорость погружения в первую очередь с размером, который, как было показано, является полезным показателем для частиц, образующихся в контролируемых средах (например, в роликовых резервуарах). [32] [33] [34] Однако сильные связи наблюдались только тогда, когда все частицы были созданы с использованием одного и того же сообщества воды и планктона. [35] Когда частицы были созданы различными планктонными сообществами, сам по себе размер был плохим предиктором (например, Diercks and Asper, 1997), что убедительно подтверждает мнение о том, что плотность и форма частиц широко варьируются в зависимости от исходного материала. [35] [25]
Упаковка и пористость в значительной степени влияют на определение скорости погружения. С одной стороны, добавление в агрегаты балластных материалов, таких как панцири диатомей, может привести к увеличению скоростей погружения за счет увеличения избыточной плотности. С другой стороны, добавление балластных минеральных частиц к популяциям морских частиц часто приводит к образованию более мелких и более плотно упакованных агрегатов, которые тонут медленнее из-за своего меньшего размера. [36] [37] Было показано, что частицы, богатые слизью, плавают, несмотря на относительно большие размеры. [28] [38] тогда как было показано, что нефтесодержащие или пластмассосодержащие агрегаты быстро тонут, несмотря на присутствие веществ с избыточной плотностью, меньшей, чем у морской воды. [39] [40] В естественной среде частицы образуются с помощью разных механизмов, разными организмами и в различных условиях окружающей среды, которые влияют на агрегацию (например, соленость, pH, минералы), балластировку (например, отложение пыли, отложения; [35] [34] van der Jagt et al., 2018) и поведение при опускании (например, вязкость; [41] ). Следовательно, универсальное преобразование размера в скорость погружения невозможно. [42] [25]
Роль в нижней водной пищевой сети
[ редактировать ]Наряду с растворенными органическими веществами ПОМ управляет нижней водной пищевой сетью, обеспечивая энергию в виде углеводов, сахаров и других полимеров, которые могут разлагаться. ПОВ в водоемах образуется из наземных источников (например, органического вещества почвы, листового опада), подводной или плавающей водной растительности или автохтонного образования водорослей (живых или детритных). Каждый источник ПОМ имеет свой собственный химический состав, который влияет на его лабильность или доступность для пищевой сети. ПОМ, полученное из водорослей, считается наиболее лабильным, но появляется все больше свидетельств того, что ПОМ наземного происхождения может дополнять рацион таких микроорганизмов, как зоопланктон, когда первичная продуктивность ограничена. [43] [44]
Биологический углеродный насос
[ редактировать ]Динамика пула твердых частиц органического углерода (POC) в океане занимает центральное место в морском углеродном цикле . POC является связующим звеном между первичной продукцией поверхности, глубиной океана и отложениями. Скорость разложения ПОУ в темном океане может повлиять на концентрацию CO 2 в атмосфере . Таким образом, центральным направлением исследований морской органической геохимии является улучшение понимания распределения, состава и круговорота ВОУ. За последние несколько десятилетий произошли улучшения в аналитических методах, которые значительно расширили возможности измерения, как с точки зрения структурного разнообразия и изотопного состава органических соединений, так и с точки зрения дополнительных молекулярных омических исследований . [9]


Как показано на схеме, фитопланктон фиксирует углекислый газ в эвфотической зоне, используя солнечную энергию, и производит ВОУ. ВОУ, образующиеся в эвфотической зоне, перерабатываются морскими микроорганизмами (микробами), зоопланктоном и их потребителями в органические агрегаты ( морской снег ), которые затем экспортируются в мезопелагическую (глубина 200–1000 м) и батипелагическую зоны погружения и вертикальной миграции путем зоопланктон и рыбы. [46] [47] [48]
Биологический углеродный насос описывает совокупность биогеохимических процессов, связанных с производством, погружением и реминерализацией органического углерода в океане. [49] [50] Короче говоря, фотосинтез микроорганизмов в верхних десятках метров водного столба фиксирует неорганический углерод (любую химическую разновидность растворенного углекислого газа) в биомассу . Когда эта биомасса опускается в глубины океана, часть ее питает метаболизм живущих там организмов, включая глубоководных рыб и донные организмы. [48] Зоопланктон играет решающую роль в формировании потока частиц посредством поглощения и фрагментации частиц. [51] [52] [53] [54] [55] [56] производство быстротонущего фекального материала [48] [30] и активная вертикальная миграция. [57] [58] [59] [25]
Помимо важности «экспортируемого» органического углерода как источника пищи для глубоководных организмов океана, биологический углеродный насос выполняет ценную экосистемную функцию: экспортируемый органический углерод ежегодно переносит в глубины океана около 5–20 Гт углерода. [60] где его часть (~0,2–0,5 Гт С) [61] изолирован на несколько тысячелетий. Таким образом, биологический углеродный насос по величине аналогичен нынешним выбросам углерода от ископаемого топлива (~ 10 Гт С в год). Любые изменения его величины, вызванные глобальным потеплением, могут иметь прямые последствия как для глубоководных организмов, так и для концентрации углекислого газа в атмосфере. [62] [47] [25]
Величина и эффективность (количество связываемого углерода по отношению к первичному производству) биологического углеродного насоса, а, следовательно, и накопление углерода в океане, частично определяются количеством экспортируемого органического вещества и скоростью его реминерализации (т. е. скоростью, с которой тонущее органическое вещество перерабатывается и дышит в районе мезопелагической зоны . [62] [63] [64] Особенно важными параметрами, определяющими, насколько быстро тонет частица, являются размер и состав частиц. [65] [63] сколько материала он содержит, [66] и какие организмы могут его найти и использовать. [67] [68] [69] [25]
Тонущими частицами могут быть фитопланктон, зоопланктон, детрит, фекальные гранулы или их смесь. [70] [71] [48] Их размер варьируется от нескольких микрометров до нескольких сантиметров, а частицы диаметром >0,5 мм называются морским снегом . [72] Обычно считается, что частицы в жидкости тонут, если их плотность превышает плотность окружающей жидкости, т. е. когда избыточная плотность превышает ноль. Таким образом, более крупные отдельные клетки фитопланктона могут способствовать потокам осадочных пород. Например, было показано, что крупные клетки диатомей и цепочки диатомей диаметром> 5 мкм тонут со скоростью до нескольких 10 метров в день, хотя это возможно только из-за тяжелого балласта кремнеземной панциря . [73] [74] И размер, и плотность влияют на скорость погружения частиц; например, для скоростей погружения, соответствующих закону Стокса , удвоение размера частицы увеличивает скорость погружения в 4 раза. [75] [73] Однако высокопористая природа многих морских частиц означает, что они не подчиняются закону Стокса, поскольку небольшие изменения плотности частиц (т.е. компактности) могут оказать большое влияние на скорость их погружения. [63] Крупные тонущие частицы обычно бывают двух типов: (1) агрегаты, образованные из ряда первичных частиц, включая фитопланктон, бактерии, фекальные гранулы , живые простейшие , зоопланктон и мусор, и (2) фекальные гранулы зоопланктона , которые могут доминировать в потоках частиц. и тонуть со скоростью, превышающей 1000 мд. −1 . [48] [25]
Знание размера, распространенности, структуры и состава (например, содержания углерода) оседающих частиц важно, поскольку эти характеристики накладывают фундаментальные ограничения на биогеохимический круговорот углерода. Например, ожидается, что изменения климата будут способствовать изменению видового состава таким образом, что изменятся элементный состав твердых частиц, размер клеток и траектория углерода через пищевую сеть , что повлияет на долю биомассы, экспортируемой на глубину. [76] Таким образом, любые вызванные климатом изменения в структуре или функциях сообществ фитопланктона, вероятно, изменят эффективность биологического углеродного насоса, что приведет к обратной связи со скоростью изменения климата. [77] [78] [25]
Гипотеза биолюминесцентного шунта
[ редактировать ]
Потребление биолюминесцентных ЧОК рыбами может привести к выделению биолюминесцентных фекальных гранул (переупаковка), которые также могут производиться с использованием небиолюминесцентных ЧОК, если кишечник рыбы уже заряжен биолюминесцентными бактериями. [80]
На диаграмме справа тонущий POC движется вниз, а за ним следует химический шлейф. [81] Простые белые стрелки обозначают поток углерода. Панель (а) представляет классический взгляд на небиолюминесцентную частицу. Длина шлейфа определяется шкалой сбоку. [82] Панель (b) представляет случай светящейся частицы в гипотезе биолюминесцентного шунта. Биолюминесцентные бактерии представлены агрегированными на частице. Их световое излучение показано в виде голубоватого облака вокруг него. Синие пунктирные стрелки обозначают визуальное обнаружение и движение к частице организмов-потребителей. Увеличение визуального обнаружения позволяет лучше обнаруживать высшие трофические уровни, что потенциально может привести к фрагментации тонущих ПОУ во взвешенные ПОУ из-за неаккуратного кормления. [80]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Монрой, П., Эрнандес-Гарсия, Э., Росси, В. и Лопес, К. (2017) «Моделирование динамического опускания биогенных частиц в океаническом потоке». Нелинейные процессы в геофизике , 24 (2): 293–305. дои : 10.5194/npg-24-293-2017 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 3.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 3.0 . - ^ Саймон М., Гроссарт Х., Швейцер Б. и Плауг Х. (2002) «Микробная экология органических агрегатов в водных экосистемах». Водная микробная экология , 28 : 175–211. дои : 10.3354/ame028175 .
- ^ Камбарделла, Калифорния; Эллиотт, ET (1991). «Изменение органических веществ в твердых частицах почвы в процессе возделывания пастбищ». Журнал Американского общества почвоведения . 56 (3): 777–783. дои : 10.2136/sssaj1992.03615995005600030017x .
- ^ Муди, К.С. и Уорролл, Ф. (2017) «Моделирование скорости разложения DOC с использованием состава DOM и гидроклиматических переменных». Журнал геофизических исследований: Biogeosciences , 122 (5): 1175–1191. дои : 10.1002/2016JG003493 .
- ^ Jump up to: а б с д Брейди, Северная Каролина; Вейль, Р.Р. (2007). Природа и свойства почв (11-е изд.). Река Аппер-Сэддл, Нью-Джерси: Prentice-Hall Inc.
- ^ Jump up to: а б с д и ж Грегорич, Э.Г.; Беар, Миннесота; Макким, УФ; Скьемстад, Дж. О. (2006). «Химические и биологические характеристики физически незакомплексованного органического вещества». Журнал Американского общества почвоведения . 70 (3): 975–985. Бибкод : 2006SSASJ..70..975G . дои : 10.2136/sssaj2005.0116 .
- ^ Картер, MR (1993). Отбор проб почвы и методы анализа . ЦРК Пресс.
- ^ Jump up to: а б с «Твердые органические вещества» . Качество почвы для здоровья окружающей среды . НРКС.
- ^ Jump up to: а б с д и Харбуш, Дж.Дж., Клоуз, Х.Г., Ван Муй, Б.А., Арности, К., Смиттенберг, Р.Х., Ле Муань, Ф.А., Молленхауэр, Г., Шольц-Бетчер, Б., Обрет, И., Кох, Б.П. и Беккер, К. (2020) «Деконструирование твердых частиц органического углерода: молекулярный и химический состав твердых частиц органического углерода в океане». Границы морской науки , 7 : 518. два : 10.3389/fmars.2020.00518 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Вагнер, Саша; Шубоц, Флоренция; Кайзер, Карл; Халлманн, Кристиан; Васка, Ханнелор; Россель, Памела Э.; Хансман, Роберта; Элверт, Маркус; Мидделбург, Джек Дж.; Энгель, Аня; Блаттманн, Томас М.; Катала, Тереза С.; Леннарц, Синикка Т.; Гомес-Саес, Гонсало В.; Пантоха-Гутьеррес, Сильвио; Бао, Руй; Гали, Валье (2020). «Предсказывающий ДОМ: современный взгляд на будущее океанического растворенного органического углерода» . Границы морской науки . 7 . дои : 10.3389/fmars.2020.00341 . hdl : 21.11116/0000-0006-6F5D-7 .
- ^ Райли, Дж.С.; Сандерс, Р.; Марсей, К.; Ле Муань, КВС; Ахтерберг, Е.П.; Поултон, Эй Джей (2012). «Относительный вклад быстрых и медленно тонущих частиц в экспорт углерода из океана» . Глобальные биогеохимические циклы . 26 (1): н/д. Бибкод : 2012GBioC..26.1026R . дои : 10.1029/2011GB004085 . S2CID 41836211 .
- ^ Лам, Фиби Дж.; Дони, Скотт С.; Бишоп, Джеймс КБ (2011). «Динамический биологический насос океана: выводы из глобальной компиляции профилей концентрации твердых частиц органического углерода, CaCO3 и опала из мезопелагических слоев» . Глобальные биогеохимические циклы . 25 (3): н/д. Бибкод : 2011GBioC..25.3009L . дои : 10.1029/2010GB003868 .
- ^ Эппли, Ричард В.; Петерсон, Брюс Дж. (1979). «Поток твердых частиц органического вещества и новая продукция планктона в глубоком океане». Природа . 282 (5740): 677–680. Бибкод : 1979Natur.282..677E . дои : 10.1038/282677a0 . S2CID 42385900 .
- ^ Волк, Тайлер; Хофферт, Мартин И. (2013). «Углеродные насосы океана: анализ относительной силы и эффективности изменений CO2 в атмосфере, вызванных океаном». Углеродный цикл и атмосферный CO2: естественные вариации от архейского периода до настоящего времени . Серия геофизических монографий. стр. 99–110. дои : 10.1029/gm032p0099 . ISBN 9781118664322 .
- ^ Бойд, П.В.; Трулл, ТВ (2007). «Понимание экспорта биогенных частиц в океанических водах: есть ли консенсус?» . Прогресс в океанографии . 72 (4): 276–312. Бибкод : 2007Proce..72..276B . дои : 10.1016/j.pocean.2006.10.007 .
- ^ Каван, Эль; Ле Муань, КВС; Поултон, Эй Джей; Тарлинг, Джорджия; Уорд, П.; Дэниелс, CJ; Фрагозо, генеральный директор; Сандерс, Р.Дж. (2015). «Ослабление потока твердых частиц органического углерода в море Скоша в Южном океане контролируется фекальными гранулами зоопланктона» . Письма о геофизических исследованиях . 42 (3): 821–830. Бибкод : 2015GeoRL..42..821C . дои : 10.1002/2014GL062744 . S2CID 53508151 .
- ^ Ле Муань, Фредерик AC (2019). «Пути нисходящего транспорта органического углерода океаническим биологическим углеродным насосом» . Границы морской науки . 6 . дои : 10.3389/fmars.2019.00634 .
- ^ Jump up to: а б с д и ж г Почва: формы и функции Викторианские ресурсы в Интернете . Обновлено 23 марта 2020 г. Текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б Бэлдок Дж. А. и Скьемстад Дж. О. (1999) «Почвенный органический углерод/почвенное органическое вещество в почве». В книге К. И. Певерилла, Л. А. Спарроу и Д. Д. Рейтера (ред.) Анализ почвы: руководство по интерпретации , страницы 159–170, Организация Содружества по научным и промышленным исследованиям, Мельбурн. ISBN 9780643063761
- ^ Jump up to: а б с Шесть, Дж.; Боссайт, Х.; Дегрыз, С; Денеф, К. (2004). «История исследований связи между (микро) агрегатами, почвенной биотой и динамикой органического вещества почвы». Исследования почвы и обработки почвы . 79 (1): 7–31. Бибкод : 2004STilR..79....7S . дои : 10.1016/j.still.2004.03.008 .
- ^ Jump up to: а б с д Оманд М.М., Говиндараджан Р., Хе Дж. и Махадеван А. (2020) «Поток твердых частиц органического вещества в океанах: чувствительность к характеристикам частиц». Научные отчеты , 10 (1): 1–16. два : 10.1038/s41598-020-60424-5 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Бланшар, Дж. Л., Хенеган, Р. Ф., Эверетт, Дж. Д., Требилко, Р. и Ричардсон, А. Дж. (2017) «От бактерий до китов: использование спектров функциональных размеров для моделирования морских экосистем. Тенденции в экологии и эволюции, 32 (3), стр.174-186. дои : 10.1016/j.tree.2016.12.003 .
- ^ Голландия, HD (2006) «Насыщение кислородом атмосферы и океанов». Философские труды Королевского общества B: Биологические науки , 361 (1470): 903–915. два : 10.1098/rstb.2006.1838 .
- ^ Хайнце, К., Мейер, С., Горис, Н., Андерсон, Л., Стейнфельдт, Р., Чанг, Н., Кере, К.Л. и Баккер, округ Колумбия (2015) «Поглотитель углерода в океане – воздействие, уязвимости и вызовы». Динамика системы Земли , 6 (1): 327–358. дои : 10.5194/esd-6-327-2015 .
- ^ Jump up to: а б с д и ж г час я Гиринг, С.Л., Каван, Э.Л., Базедов, С.Л., Бриггс, Н., Берд, А.Б., Дэррок, Л.Дж., Гуиди, Л., Иррисон, Дж.О., Иверсен, М.Х., Кико, Р. и Линдси, DJ (2020) " Тонущие органические частицы в океане — оценки потоков с помощью оптических устройств, находящихся на месте». два : 10.3389/fmars.2019.00834 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Конте, Морин Х ; Ральф, Нейт; Росс, Эдит Х (2001). «Сезонная и межгодовая изменчивость потоков частиц в глубоком океане на участке Программы океанических потоков (OFP)/Бермудских атлантических временных рядов (BATS) в западной части Саргассова моря возле Бермудских островов» . Глубоководные исследования, часть II: Актуальные исследования в океанографии . 48 (8–9): 1471–1505. Бибкод : 2001DSRII..48.1471C . дои : 10.1016/S0967-0645(00)00150-8 .
- ^ Эппли, Р.В. и Петерсон, Б.Дж. (1979) «Поток твердых частиц органического вещества и новое производство планктона в глубоком океане». Природа , 282 (5740): 677–680. два : 10.1038/282677a0 .
- ^ Jump up to: а б Азецу-Скотт, Кумико; Пассов, Ута (2004). «Восходящие морские частицы: значение прозрачных экзополимерных частиц (ТЭП) в верхних слоях океана» (PDF) . Лимнология и океанография . 49 (3): 741–748. Бибкод : 2004LimOc..49..741A . дои : 10.4319/lo.2004.49.3.0741 . S2CID 32205017 .
- ^ Акунья, JL; Лопес-Альварес, М.; Ногейра, Э.; Гонсалес-Табоада, Ф. (2010). «Флотация диатомей в начале весеннего цветения фитопланктона: эксперимент in situ» . Серия «Прогресс в области морской экологии» . 400 : 115–125. Бибкод : 2010MEPS..400..115A . дои : 10.3354/meps08405 . hdl : 10651/7011 .
- ^ Jump up to: а б Иверсен М.Х., Пахомов Е.А., Хант Б.П., Ван дер Ягт Х., Вольф-Гладроу Д. и Клаас К. (2017) «Грузила или плавающие поплавки? Вклад гранул сальпы в экспортный поток во время большого цветения событие в Южном океане». Глубоководные исследования, часть II: Актуальные исследования в океанографии , 138 : 116–125. дои : 10.1016/j.dsr2.2016.12.004 .
- ^ Уайт, Фрэнк М. (2006). Поток вязкой жидкости . МакГроу-Хилл. ISBN 9780071244930 .
- ^ Гердес, Астрид; Иверсен, Мортен Х.; Гроссарт, Ганс-Петер; Пассов, Ута; Ульрих, Матиас С. (2011). «Диатомовые бактерии необходимы для агрегации Thalassiosira weissflogii» . Журнал ISME . 5 (3): 436–445. Бибкод : 2011ISMEJ...5..436G . дои : 10.1038/ismej.2010.145 . ПМК 3105730 . ПМИД 20827289 .
- ^ Иверсен, МХ; Плауг, Х. (2013). «Влияние температуры на скорость дыхания углерода и скорость погружения скоплений диатомей - потенциальные последствия для процессов экспорта из глубокого океана» . Биогеонауки . 10 (6): 4073–4085. Бибкод : 2013BGeo...10.4073I . дои : 10.5194/bg-10-4073-2013 .
- ^ Jump up to: а б Иверсен, Мортен Х.; Роберт, Майя Л. (2015). «Балластирующее воздействие смектита на образование агрегатов и их экспорт из природного планктонного сообщества». Морская химия . 175 : 18–27. Бибкод : 2015МартЧ.175...18I . дои : 10.1016/j.marchem.2015.04.009 .
- ^ Jump up to: а б с Иверсен, Мортен Хвитфельдт; Новальд, Николас; Плауг, Хелле; Джексон, Джордж А.; Фишер, Герхард (2010). «Профили высокого разрешения вертикального вывоза твердых частиц у мыса Блан, Мавритания: процессы деградации и эффекты балласта». Глубоководные исследования. Часть I: Статьи океанографических исследований . 57 (6): 771–784. Бибкод : 2010DSRI...57..771I . дои : 10.1016/j.dsr.2010.03.007 .
- ^ Хамм, Кристиан Э. (2002). «Интерактивная агрегация и седиментация диатомей и глинистого литогенного материала» . Лимнология и океанография . 47 (6): 1790–1795. Бибкод : 2002LimOc..47.1790H . дои : 10.4319/lo.2002.47.6.1790 .
- ^ Пассов, Ута; де ла Роша, Кристина Л.; Фэрфилд, Кейтлин; Шмидт, Катрин (2014). «Агрегация как функция минеральных частиц» . Лимнология и океанография . 59 (2): 532–547. Бибкод : 2014LimOc..59..532P . дои : 10.4319/lo.2014.59.2.0532 .
- ^ Бочданский, Александр Б.; Клаус, Мелисса А.; Херндль, Герхард Дж. (2016). «Короли драконов морских глубин: морские частицы заметно отклоняются от общепринятого спектра размеров» . Научные отчеты . 6 : 22633. Бибкод : 2016NatSR...622633B . дои : 10.1038/srep22633 . ПМЦ 4778057 . ПМИД 26940454 .
- ^ Лонг, Марк; Морисо, Бриваэла; Галлинари, Морган; Ламберт, Кристоф; Юве, Арно; Раффрэ, Жан; Судан, Филипп (2015). «Взаимодействие между микропластиком и агрегатами фитопланктона: влияние на их судьбу» . Морская химия . 175 : 39–46. Бибкод : 2015МартЧ.175...39Л . дои : 10.1016/j.marchem.2015.04.003 .
- ^ Пассоу, У.; Свит, Дж.; Фрэнсис, С.; Сюй, К.; Диссанаяке, Алабама; Лин, ГГ; Санчи, PH; Куигг, А. (2019). «Включение нефти в диатомовые агрегаты» . Серия «Прогресс в области морской экологии» . 612 : 65–86. Бибкод : 2019MEPS..612...65P . дои : 10.3354/meps12881 .
- ^ Таучер, Дж.; Бах, LT; Рибеселл, У.; Ошлис, А. (2014). «Влияние вязкости на поток морских частиц: механизм обратной связи, связанный с климатом» (PDF) . Глобальные биогеохимические циклы . 28 (4): 415–422. Бибкод : 2014GBioC..28..415T . дои : 10.1002/2013GB004728 .
- ^ Жуанде, Мари-Поль; Трулл, Томас В.; Гуиди, Лайонел; Пишераль, Марк; Эберсбах, Фридерика; Стемманн, Ларс; Блен, Стефан (2011). «Оптические изображения мезопелагических частиц указывают на глубокий поток углерода под естественным, обогащенным железом цветением в Южном океане» . Лимнология и океанография . 56 (3): 1130–1140. Бибкод : 2011LimOc..56.1130J . дои : 10.4319/lo.2011.56.3.1130 .
- ^ Вейдель, Брайан; Соломон, Кристофер Т.; Пейс, Майкл Л.; Китчелл, Джим; Карпентер, Стивен Р.; Коул, Джонатан Дж. (1 февраля 2011 г.). «Убедительные доказательства существования наземного существования зоопланктона в небольших озерах на основе стабильных изотопов углерода, азота и водорода» . Труды Национальной академии наук . 108 (5): 1975–1980. Бибкод : 2011PNAS..108.1975C . дои : 10.1073/pnas.1012807108 . ISSN 0027-8424 . ПМК 3033307 . ПМИД 21245299 .
- ^ Канкаала, Паула; Страндберг, Урсула; Киммо К. Кахилайнен; Аалто, Санни Л.; Галлоуэй, Аарон МЫ; Тайпале, Сами Дж. (11 августа 2016 г.). «Наземные углеводы поддерживают пресноводный зоопланктон во время дефицита фитопланктона» . Научные отчеты . 6 : 30897. Бибкод : 2016NatSR...630897T . дои : 10.1038/srep30897 . ISSN 2045-2322 . ПМК 4980614 . ПМИД 27510848 .
- ^ Хенли, Сиан Ф.; Каван, Эмма Л.; Фосетт, Сара Э.; Керр, Родриго; Монтейро, Тьяго; Шеррелл, Роберт М.; Боуи, Эндрю Р.; Бойд, Филип В.; Барнс, Дэвид К.А.; Шлосс, Ирен Р.; Маршалл, Таня; Флинн, Ракель; Смит, Шантель (2020). «Изменение биогеохимии Южного океана и его экосистемные последствия» . Границы морской науки . 7 . дои : 10.3389/fmars.2020.00581 . hdl : 11336/128446 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Басу, С. и Макки, К.Р. (2018) «Фитопланктон как ключевые медиаторы биологического углеродного насоса: их реакция на изменение климата». Устойчивое развитие , 10 (3): 869. два : 10.3390/su10030869 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б Пассоу, У. и Карлсон, Калифорния (2012) «Биологический насос в мире с высоким содержанием CO2». Серия достижений морской экологии , 470 : 249–271. два : 10.3354/meps09985 .
- ^ Jump up to: а б с д и Тернер, Джефферсон Т. (2015). «Фекальные гранулы зоопланктона, морской снег, фитодетрит и биологический насос океана». Прогресс в океанографии . 130 : 205–248. Бибкод : 2015Proce.130..205T . дои : 10.1016/j.pocean.2014.08.005 .
- ^ Волк, Т. и Хофферт, М.И. (1985) «Океанские углеродные насосы: анализ относительной силы и эффективности изменений атмосферного CO2, вызванных океаном. В: Углеродный цикл и атмосферный CO2: естественные вариации от архея до настоящего времени» , страницы 99–110. , Калифорнийский университет. ISBN 9780875900605 .
- ^ Гиринг, С.Л. и Хамфрис, член парламента (2018) «Биологический насос». В: Энциклопедия геохимии , У. Уайт (ред.) Чам: Спрингер, страницы 1–6. дои : 10.1007/978-3-319-39193-9_154-1 .
- ^ Уэйт, А.М., Сафи, К.А., Холл, Дж.А. и Ноддер, С.Д. (2000) «Массовое осаждение пикопланктона, встроенного в органические агрегаты». Лимнология и океанография , 45 (1): 87–97. дои : 10.4319/lo.2000.45.1.0087 .
- ^ Иверсен, М.Х. и Поулсен, Л.К. (2007) «Копрорексия, копрофагия и копрохалия у копепод Calanus helgolandicus , Pseudocalanus elongatus и Oithona similis ». Серия достижений морской экологии , 350 : 79–89. два : 10.3354/meps07095 .
- ^ Поулсен, Л.К. и Иверсен, М.Х. (2008) «Деградация фекальных гранул копепод: ключевая роль протозоопланктона». Серия достижений морской экологии , 367 : 1–13. два : 10.3354/meps07611 .
- ^ Иверсен, М.Х., Новальд, Н., Плауг, Х., Джексон, Г.А. и Фишер, Г. (2010) «Профили высокого разрешения вертикального экспорта твердых частиц у мыса Блан, Мавритания: процессы деградации и эффекты балласта». Глубоководные исследования, часть I: Статьи океанографических исследований , 57 (6): 771–784. дои : 10.1016/j.dsr.2010.03.007 .
- ^ Гиринг С.Л., Сандерс Р., Лэмпитт Р.С., Андерсон Т.Р., Тамбурини К., Бутриф М., Зубков М.В., Марсей К.М., Хенсон С.А., Соу К. и Кук К. ( 2014) «Сверка углеродного баланса в сумеречной зоне океана». Природа , 507 (7493): 480–483. дои : 10.1038/nature13123 .
- ^ Свенсен К., Мората Н. и Рейгстад М. (2014) «Увеличенная деградация фекальных гранул веслоногих раков в результате совместного действия динофлагеллят и Centropages hamatus ». Серия достижений морской экологии , 516 : 61–70. два : 10.3354/meps10976 .
- ^ Стейнберг, Д.К., Карлсон, Калифорния, Бейтс, Н.Р., Голдтуэйт, С.А., Мадин, Л.П. и Майклс, А.Ф. (2000) «Вертикальная миграция зоопланктона и активный перенос растворенного органического и неорганического углерода в Саргассовом море». Глубоководные исследования, часть I: Статьи океанографических исследований , 47 (1): 137–158. два : 10.1016/S0967-0637(99)00052-7 .
- ^ Йонасдоттир, С.Х., Виссер, А.В., Ричардсон, К. и Хит, М.Р. (2015) «Сезонный липидный насос копепод способствует секвестрации углерода в глубокой части Северной Атлантики». Труды Национальной академии наук , 112 (39): 12122–12126. дои : 10.1073/pnas.1512110112 .
- ^ Кико Р., Биастох А., Брандт П., Краватт С., Хаусс Х., Хуммельс Р., Крист И., Марин Ф., МакДоннелл А.М., Ошлис А. и Пичерал, М. (2017) «Биологическое и физическое влияние на морской снегопад на экваторе». Nature Geoscience , 10 (11): 852–858. дои : 10.1038/ngeo3042 .
- ^ Хенсон, С.А., Сандерс, Р., Мэдсен, Э., Моррис, П.Дж., Ле Муань, Ф. и Куартли, Г.Д. (2011) «Уменьшенная оценка силы биологического углеродного насоса океана». Письма о геофизических исследованиях , 38 (4). дои : 10.1029/2011GL046735 .
- ^ Гуиди Л., Лежандр Л., Рейгондо Г., Уитц Дж., Стемманн Л. и Хенсон С.А. (2015) «Новый взгляд на реминерализацию углерода океана для оценки глубоководной секвестрации». Глобальные биогеохимические циклы , 29 (7): 1044–1059. два : 10.1002/2014GB005063 .
- ^ Jump up to: а б Квон, Э.Ю., Примо, Ф. и Сармьенто, Дж.Л. (2009) «Влияние глубины реминерализации на углеродный баланс воздуха и моря». Nature Geoscience , 2 (9): 630–635. два : 10.1038/ngeo612 .
- ^ Jump up to: а б с Иверсен М. и Плауг Х. (2010) «Балластные минералы и опускающийся поток углерода в океане: скорость дыхания углерода и скорость погружения агрегатов морского снега». Биогеонауки , 7 : 2613–2624. дои : 10.5194/bg-7-2613-2010 .
- ^ Рейгондо, Г., Гуиди, Л., Богранд, Г., Хенсон, С.А., Кубби, П., Маккензи, Б.Р., Саттон, Т.Т., Фиорони, М. и Мори, О. (2018) «Глобальные биогеохимические провинции мезопелагическая зона». Журнал биогеографии , 45 (2): 500–514. два : 10.1111/jbi.13149 .
- ^ Плауг, Х., Иверсен, М.Х., Коски, М. и Буитенхейс, Э.Т. (2008) «Производство, скорость дыхания кислорода и скорость погружения фекальных гранул копепод: прямые измерения балласта опалом и кальцитом». Лимнология и океанография , 53 (2): 469–476. дои : 10.4319/lo.2008.53.2.0469 .
- ^ Плауг, Х., Иверсен, М.Х. и Фишер, Г. (2008) «Балласт, скорость погружения и кажущаяся диффузия в морском снеге и фекальных гранулах зоопланктона: последствия для оборота субстрата прикрепленными бактериями». Лимнология и океанография , 53 (5): 1878–1886. дои : 10.4319/lo.2008.53.5.1878 .
- ^ Кьёрбо Т., Саиз Э. и Виссер А. (1999) «Восприятие гидродинамического сигнала веслоногим раком Acartia тонна ». Серия достижений морской экологии , 179 : 97–111. два : 10.3354/meps179097 .
- ^ Виссер, AW (2001) «Гидромеханические сигналы в планктоне». Серия достижений морской экологии , 222 : 1–24. два : 10.3354/meps222001 .
- ^ Виссер, А.В. и Джексон, Джорджия (2004) «Характеристики химического шлейфа за тонущей частицей в турбулентном столбе воды». Серия достижений морской экологии , 283 : 55–71. два : 10.3354/meps283055 .
- ^ Саймон, М.; Гроссарт, HP; Швейцер, Б.; Плауг, Х. (2002). «Микробная экология органических агрегатов водных экосистем» . Водная микробная экология . 28 : 175–211. дои : 10.3354/ame028175 .
- ^ Тернер, Дж. Т. (2002). «Фекальные гранулы зоопланктона, морской снег и тонущее цветение фитопланктона» . Водная микробная экология . 27 : 57–102. дои : 10.3354/ame027057 .
- ^ Олдридж, Элис Л.; Сильвер, Мэри В. (1988). «Характеристика, динамика и значение морского снега». Прогресс в океанографии . 20 (1): 41–82. Бибкод : 1988Proce..20...41A . дои : 10.1016/0079-6611(88)90053-5 .
- ^ Jump up to: а б Уэйт, А.; Фишер, А.; Томпсон, Пенсильвания; Харрисон, Пи Джей (1997). «Зависимость скорости погружения от объема клеток проливает свет на механизмы контроля скорости погружения морских диатомовых водорослей» . Серия «Прогресс в области морской экологии» . 157 : 97–108. Бибкод : 1997MEPS..157...97W . дои : 10.3354/meps157097 .
- ^ Миклас, Кевин А.; Денни, Марк В. (2010). «Скорость погружения диатомей: улучшенные прогнозы и понимание модифицированного закона Стокса» . Лимнология и океанография . 55 (6): 2513–2525. Бибкод : 2010LimOc..55.2513M . дои : 10.4319/lo.2010.55.6.2513 .
- ^ Мур, Дж. Кейт; Вильяреал, Трейси А. (1996). «Взаимосвязь размера и скорости всплытия у морских диатомовых водорослей с положительной плавучестью» . Лимнология и океанография . 41 (7): 1514–1520. Бибкод : 1996LimOc..41.1514M . дои : 10.4319/lo.1996.41.7.1514 .
- ^ Финкель З.В.; Бердалл, Дж.; Флинн, К.Дж.; Куигг, А.; Рис, ТАВ; Рэйвен, Дж.А. (2010). «Фитопланктон в меняющемся мире: размер клеток и элементарная стехиометрия» . Журнал исследований планктона . 32 : 119–137. дои : 10.1093/plankt/fbp098 .
- ^ Матир, Ричард Дж.; Херст, Энтони К. (1999). «Отзывы об изменении климата о будущем поглощении CO2 океаном». Теллус Б: Химическая и физическая метеорология . 51 (3): 722–733. Бибкод : 1999TellB..51..722M . дои : 10.3402/tellusb.v51i3.16472 .
- ^ Ле Кер, К.; Роденбек, К.; Буитенхейс, ET; Конвей, Ти Джей; Лангенфельдс, Р.; Гомес, А.; Лабушань, К.; Рамонет, М.; Наказава, Т.; Мецль, Н.; Джиллетт, Н.; Хейманн, М. (2007). «Насыщение поглотителя CO2 Южного океана из-за недавнего изменения климата» . Наука . 316 (5832): 1735–1738. Бибкод : 2007Sci...316.1735L . дои : 10.1126/science.1136188 . ПМИД 17510327 . S2CID 34642281 .
- ^ Азам, Фарук; Лонг, Ричард А. (2001). «Морские снежные микрокосмы». Природа . 414 (6863): 495–498. дои : 10.1038/35107174 . ПМИД 11734832 . S2CID 5091015 .
- ^ Jump up to: а б с Танет, Лиза; Мартини, Северин; Казалот, Лори; Тамбурини, Кристиан (2020). «Обзоры и синтезы: Бактериальная биолюминесценция – экология и влияние на биологический углеродный насос» . Биогеонауки . 17 (14): 3757–3778. Бибкод : 2020BGeo...17.3757T . дои : 10.5194/bg-17-3757-2020 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Кьёрбо, Томас (2011). «Как питается зоопланктон: механизмы, особенности и компромиссы». Биологические обзоры . 86 (2): 311–339. дои : 10.1111/j.1469-185X.2010.00148.x . ПМИД 20682007 . S2CID 25218654 .
- ^ Кьёрбо, Томас; Джексон, Джордж А. (2001). «Морской снег, шлейфы органических растворов и оптимальное хемосенсорное поведение бактерий» . Лимнология и океанография . 46 (6): 1309–1318. Бибкод : 2001LimOc..46.1309K . дои : 10.4319/lo.2001.46.6.1309 . S2CID 86713938 .