Коэффициент Редфилда
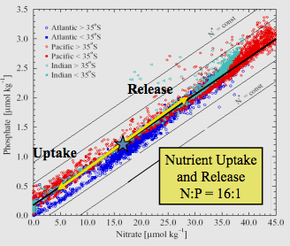
Отношение Редфилда или стехиометрия Редфилда — это постоянное атомное соотношение углерода , азота и фосфора, обнаруженное в морском фитопланктоне и во всех глубоких океанах.
Термин назван в честь американского океанографа Альфреда Редфилда , который в 1934 году впервые описал относительно постоянное соотношение питательных веществ в образцах морской биомассы, собранных во время нескольких рейсов на борту исследовательского судна «Атлантис» , и эмпирически установил, что это соотношение равно C:N:P = 106:16:1. [1] Хотя отклонения от канонического соотношения 106:16:1 были обнаружены в зависимости от вида фитопланктона и района исследования, соотношение Редфилда остается важным ориентиром для океанографов, изучающих ограничение питательных веществ. В документе 2014 года, обобщающем большой набор данных по измерениям питательных веществ во всех основных регионах океана с 1970 по 2010 год, сообщается, что глобальное медианное значение C:N:P составляет 163:22:1. [2]
Открытие
[ редактировать ]| Часть серии о |
| Углеродный цикл |
|---|
 |
В своей статье 1934 года Альфред Редфилд проанализировал данные по нитратам и фосфатам для Атлантического , Индийского , Тихого океанов и Баренцева моря . [1] Будучи физиологом из Гарварда , Редфилд участвовал в нескольких рейсах на борту исследовательского судна «Атлантис» , анализируя данные о содержании C, N и P в морском планктоне, а также ссылался на данные, собранные другими исследователями еще в 1898 году.
Анализ эмпирических данных, проведенный Редфилдом, привел к тому, что он обнаружил, что в трех океанах и Баренцевом море морская вода имеет атомное соотношение N:P около 20:1 (позже скорректированное до 16:1) и очень похоже на среднее значение. N:P фитопланктона.
Чтобы объяснить это явление, Редфилд первоначально предложил два взаимоисключающих механизма:
I) Состав N:P в планктоне приближается к составу N:P морской воды. В частности, виды фитопланктона с разными потребностями в азоте и фосфоре конкурируют в одной и той же среде и отражают состав питательных веществ морской воды. [1]
II) Равновесие между морской водой и планктонными питательными веществами поддерживается посредством биотических механизмов обратной связи. [1] [3] Редфилд предложил сценарий, подобный термостату, в котором деятельность азотфиксаторов и денитрификаторов поддерживает соотношение нитратов и фосфатов в морской воде близкое к потребностям в протоплазме. [4] Учитывая, что в то время мало что было известно о составе «протоплазмы» или общем составе фитопланктона, Редфилд не пытался объяснить, почему соотношение N:P должно составлять примерно 16:1.
В 1958 году, почти через четверть века после первого открытия этих соотношений, Редфилд склонялся к последнему механизму в своей рукописи «Биологический контроль химических факторов в окружающей среде». [3] Редфилд предположил, что соотношение азота и фосфора в планктоне приводит к тому, что в мировом океане наблюдается удивительно похожее соотношение растворенных нитратов и фосфатов (16:1). Он рассмотрел, как циклы не только N и P, но также C и O могут взаимодействовать, приводя к такому совпадению.
Объяснение
[ редактировать ]Редфилд обнаружил поразительное соответствие между химией глубин океана и химией живых существ, таких как фитопланктон на поверхности океана. Оба имеют соотношение N:P примерно 16:1 в пересчете на атомы. Когда питательные вещества не являются лимитирующими , молярное соотношение элементов C:N:P в большинстве фитопланктона составляет 106:16:1. Редфилд считал, что не случайно то, что химический состав обширных океанов идеально соответствует потребностям живых организмов.
Лабораторные эксперименты в контролируемых химических условиях показали, что биомасса фитопланктона будет соответствовать коэффициенту Редфилда, даже если уровни питательных веществ в окружающей среде превышают их, что позволяет предположить, что экологическая адаптация к соотношениям питательных веществ в океане не является единственным регулирующим механизмом (в отличие от одного из механизмов, первоначально предложенных Редфилдом). ). [5] Однако последующее моделирование механизмов обратной связи, в частности потоков нитрат-фосфорного взаимодействия, подтверждает предложенный им механизм биотического равновесия с обратной связью, хотя эти результаты опровергаются ограничениями в нашем нынешнем понимании потоков питательных веществ. [6]
В океане большую часть биомассы составляет планктон, богатый азотом. Многие из этих планктонов потребляются другой планктонной биомассой, имеющей аналогичный химический состав. Это приводит к аналогичному соотношению N:P в среднем для всего планктона во всем мировом океане, которое, как установлено эмпирически, составляет примерно 16:1. Когда эти организмы погружаются в недра океана, их биомасса потребляется бактериями, которые в аэробных условиях окисляют органическое вещество с образованием растворенных неорганических питательных веществ, главным образом углекислого газа , нитратов и фосфатов.
То, что соотношение нитратов и фосфатов во внутренней части всех основных океанских бассейнов очень похоже, возможно, связано с временем пребывания этих элементов в океане относительно времени циркуляции океана: примерно 100 000 лет для фосфора и 2000 лет для азота. . [7] Тот факт, что времена пребывания этих элементов превышают времена смешения океанов (~1000 лет) [8] может привести к тому, что соотношение нитратов и фосфатов во внутренней части океана останется довольно однородным. Было показано, что фитопланктон играет ключевую роль в поддержании этого соотношения. Когда органическое вещество опускается, нитраты и фосфаты выбрасываются в океан в результате реминерализации. Микроорганизмы преимущественно потребляют кислород в нитратах, а не в фосфатах, что приводит к образованию более глубоких океанических вод с соотношением N:P менее 16:1. Оттуда океанские течения поднимают питательные вещества на поверхность, где фитопланктон поглощает избыток фосфора и поддерживает соотношение N:P 16:1, потребляя N 2 посредством фиксации азота. [9] Хотя такие аргументы потенциально могут объяснить, почему эти соотношения довольно постоянны, они не отвечают на вопрос, почему соотношение N:P составляет почти 16, а не какое-то другое число.
Использование
[ редактировать ]Исследования, в результате которых было установлено это соотношение, стали фундаментальной особенностью понимания биогеохимических циклов океанов и одним из ключевых принципов биогеохимии. Коэффициент Редфилда играет важную роль в оценке потоков углерода и питательных веществ в моделях глобальной циркуляции . Они также помогают определить, какие питательные вещества являются лимитирующими в локализованной системе, если такое питательное вещество существует. Это соотношение также можно использовать для понимания формирования цветения фитопланктона и последующей гипоксии путем сравнения соотношения между различными регионами, например, сравнения отношения Редфилда реки Миссисипи с соотношением северной части Мексиканского залива. [10] Контроль N:P может стать средством устойчивого управления резервуарами. [11] Возможно даже, что коэффициент Редфилда применим к наземным растениям, почвам и почвенной микробной биомассе, что может указывать на ограничение ресурсов в наземных экосистемах. [12] В исследовании 2007 года было обнаружено, что почва и микробная биомасса имеют постоянное соотношение C:N:P 186:13:1 и 60:7:1 соответственно в среднем в глобальном масштабе. [12]
Отклонения от канонического соотношения Редфилда
[ редактировать ]Коэффициент Редфилда первоначально был получен эмпирически на основе измерений элементного состава планктона, а также содержания нитратов и фосфатов в морской воде, собранной на нескольких станциях в Атлантическом океане . Позже это было подтверждено сотнями независимых измерений растворенных нитратов и фосфатов. Однако состав отдельных видов фитопланктона, выращенных в условиях ограничения по азоту или фосфору, показывает, что соотношение N:P может варьироваться от 6:1 до 60:1. Понимая эту проблему, Редфилд никогда не пытался объяснить ее, за исключением того, что отметил, что соотношение N:P неорганических питательных веществ во внутренней части океана было средним с ожидаемой небольшой изменчивостью.
Хотя соотношение Редфилда удивительно стабильно в глубоком океане, было широко показано, что фитопланктон может иметь большие различия в составе C:N:P, и их жизненная стратегия играет роль в соотношении C:N:P. Эта изменчивость заставила некоторых исследователей предположить, что коэффициент Редфилда, возможно, является общим средним показателем в современном океане, а не фундаментальной особенностью фитопланктона. [13] хотя также утверждалось, что это связано с гомеостатическим соотношением белка и рРНК , которое фундаментально присутствует как у прокариот, так и у эукариот, что способствует тому, что он является наиболее распространенным составом. [14] Существует несколько возможных объяснений наблюдаемой изменчивости соотношений C:N:P. Скорость роста клетки влияет на ее состав и, следовательно, на ее стехиометрию. [15] Кроме того, когда фосфора не хватает, сообщества фитопланктона могут снизить содержание фосфора, повышая соотношение N:P. [16] Кроме того, накопление и количество мертвого фитопланктона и детрита могут влиять на доступность определенных источников пищи, что, в свою очередь, влияет на состав клетки. [17] Также было показано, что в некоторых экосистемах соотношение Редфилда значительно варьируется в зависимости от доминирующих таксонов фитопланктона, присутствующих в экосистеме, даже в системах с обильным содержанием питательных веществ. Следовательно, коэффициент Редфилда для конкретной системы может служить показателем структуры планктонного сообщества. [18]
Несмотря на сообщения о том, что элементный состав таких организмов , как морской фитопланктон в океаническом регионе, не соответствует каноническому соотношению Редфилда, фундаментальная концепция этого соотношения остается актуальной и полезной.
Расширенное соотношение Редфилда
[ редактировать ]Некоторые считают, что существуют и другие элементы, такие как калий , сера , цинк , медь и железо , которые также важны для химии океана . [19]
В частности, железу (Fe) придавалось большое значение, поскольку ранние биологи-океанографы предположили, что железо также может быть ограничивающим фактором для первичного производства в океане. [20] С тех пор эксперименты доказали, что железо является ограничивающим фактором для первичного производства. Раствор, богатый железом, добавлен на 64 км. 2 площади, что привело к увеличению первичной продукции фитопланктона. [21] В результате был разработан расширенный коэффициент Редфилда, включающий его в баланс. Это новое стехиометрическое соотношение гласит, что соотношение должно составлять 106 C:16 N:1 P:0,1-0,001 Fe. Большой разброс содержания Fe является результатом значительного препятствия со стороны кораблей и научного оборудования, загрязняющих любые пробы, собранные в море, избытком Fe. [22] Именно это загрязнение привело к появлению первых данных, свидетельствующих о том, что концентрации железа высоки и не являются ограничивающим фактором первичной морской продукции.
Диатомовым водорослям , помимо других питательных веществ, необходима кремниевая кислота для создания биогенного кремнезема в панцирях (клеточных стенках). В результате этого для диатомовых водорослей было предложено соотношение питательных веществ Редфилда-Бжезинского, составляющее C:Si:N:P = 106:15:16:1. [23] Было показано, что, выходя за рамки самого первичного производства, кислород, потребляемый аэробным дыханием биомассы фитопланктона, подчиняется предсказуемой пропорции по отношению к другим элементам. O 2 :C составило 138:106. Измеренное соотношение [6]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Редфилд, AC (1934). «О пропорциях органических производных в морской воде и их связи с составом планктона» (PDF) . Мемориал Джеймса Джонстона, том : 176 . Проверено 1 марта 2019 г.
- ^ Мартини, Адам С.; Вругт, Джаспер А.; Ломас, Майкл В. (9 декабря 2014 г.). «Концентрация и соотношение твердых частиц органического углерода, азота и фосфора в мировом океане» . Научные данные . 1 (1): 140048. doi : 10.1038/sdata.2014.48 . ПМК 4421931 . ПМИД 25977799 .
- ^ Перейти обратно: а б РЕДФИЛД, АЛЬФРЕД К. (1958). «Биологический контроль химических факторов окружающей среды». Американский учёный . 46 (3): 205–221, 230А. ISSN 0003-0996 . JSTOR 27827150 . ПМИД 24545739 .
- ^ Грубер, Николас; Сармьенто, Хорхе Л. (1997). «Глобальные закономерности фиксации и денитрификации морского азота» . Глобальные биогеохимические циклы . 11 (2): 235–266. Бибкод : 1997GBioC..11..235G . дои : 10.1029/97GB00077 . ISSN 1944-9224 .
- ^ Гольдман, Джоэл К. (1986). «О скорости роста фитопланктона и соотношении частиц C:N:P при слабом освещении1» . Лимнология и океанография . 31 (6): 1358–1363. Бибкод : 1986LimOc..31.1358G . дои : 10.4319/lo.1986.31.6.1358 . ISSN 1939-5590 .
- ^ Перейти обратно: а б Лентон, Тимоти М.; Уотсон, Эндрю Дж. (2000). «Возврат к Редфилду: 1. Регулирование содержания нитратов, фосфатов и кислорода в океане» . Глобальные биогеохимические циклы . 14 (1): 225–248. Бибкод : 2000GBioC..14..225L . дои : 10.1029/1999GB900065 . ISSN 1944-9224 .
- ^ «Проект «Зеленый океан» Азотный цикл» . Архивировано из оригинала 02 апреля 2015 г. Проверено 27 марта 2015 г.
- ^ «Группа химических сенсоров» . 20 августа 2015 г.
- ^ Сигман, Дэниел М.; Хейн, Матис П.; Хауг, Джеральд Х. (июль 2010 г.). «Полярный океан и ледниковые циклы концентрации CO2 в атмосфере» . Природа . 466 (7302): 47–55. Бибкод : 2010Natur.466...47S . дои : 10.1038/nature09149 . ISSN 0028-0836 . ПМИД 20596012 . S2CID 4424883 .
- ^ Доддс, Уолтер (2006). «Питательные вещества и «мертвая зона»: связь между соотношением питательных веществ и растворенным кислородом в северной части Мексиканского залива». Границы в экологии и окружающей среде . 4 (4): 211–217. doi : 10.1890/1540-9295(2006)004[0211:NATDZT]2.0.CO;2 .
- ^ Левич, АП (1 февраля 1996 г.). «Роль соотношения азота и фосфора в выборе доминирования фитопланктона цианобактериями или зелеными водорослями и его применение для управления водоемами». Журнал здоровья водной экосистемы . 5 (1): 55–61. дои : 10.1007/BF00691729 . ISSN 1573-5141 . S2CID 84469052 .
- ^ Перейти обратно: а б Кливленд, Кори К.; Липцин, Дэниел (1 сентября 2007 г.). «Стехиометрия C:N:P в почве: существует ли «коэффициент Редфилда» для микробной биомассы?» . Биогеохимия . 85 (3): 235–252. дои : 10.1007/s10533-007-9132-0 . ISSN 1573-515X . S2CID 51898417 .
- ^ Арриго, КР (2005). «Морские микроорганизмы и глобальные круговороты питательных веществ». Природа . 437 (7057): 349–55. Бибкод : 2005Natur.437..349A . дои : 10.1038/nature04159 . ПМИД 16163345 . S2CID 62781480 .
- ^ Лоладзе, Ираклий; Элзер, Джеймс Дж. (2011). «Причина соотношения азота и фосфора Редфилда лежит в гомеостатическом соотношении белка и рРНК». Экологические письма . 14 (3): 244–250. дои : 10.1111/j.1461-0248.2010.01577.x . ISSN 1461-0248 . ПМИД 21244593 .
- ^ Клаусмайер К., Личман Э., Дофрен Т. и др. Оптимальная азотно-фосфорная стехиометрия фитопланктона. Природа 429, 171–174 (2004). https://doi.org/10.1038/nature02454
- ^ Гэлбрейт, Эрик Д.; Мартини, Адам С. (2015). «Простой механизм зависимости от питательных веществ для прогнозирования стехиометрии морских экосистем» . Труды Национальной академии наук . 112 (27): 8199–8204. Бибкод : 2015PNAS..112.8199G . дои : 10.1073/pnas.1423917112 . ПМК 4500256 . ПМИД 26056296 .
- ^ Мартини, Адам С.; Фам, Чау Т.А.; Примо, Франсуа В.; Вругт, Джаспер А.; Мур, Дж. Кейт; Левин, Саймон А.; Ломас, Майкл В. (17 марта 2013 г.). «Сильные широтные закономерности в соотношении элементов морского планктона и органического вещества» . Природа Геонауки . 6 (4): 279–283. Бибкод : 2013NatGe...6..279M . дои : 10.1038/ngeo1757 . ISSN 1752-0894 . S2CID 5677709 .
- ^ Арриго, Кевин Р.; Данбар, Роберт Б.; Лизотт, Майкл П.; Робинсон, Д.Х. (2002). «Таксоноспецифичные различия в сокращении C/P и N/P для фитопланктона в море Росса, Антарктида». Письма о геофизических исследованиях . 29 (19): 44–1–44-4. Бибкод : 2002GeoRL..29.1938A . дои : 10.1029/2002GL015277 . ISSN 1944-8007 . S2CID 129704137 .
- ^ Беннер, Р; Пакульски, доктор юридических наук; Маккарти, М; Хеджес, Дж.И.; Хэтчер, П.Г. (1992). «Объемные химические характеристики растворенного органического вещества в океане». Наука . 255 (5051): 1561–4. Бибкод : 1992Sci...255.1561B . дои : 10.1126/science.255.5051.1561 . ПМИД 17820170 . S2CID 28604425 .
- ^ Тортелл, PD; Мальдонадо, Монтана; Грейнджер, Дж; Прайс, Нью-Мексико (1999). «Морские бактерии и биогеохимический круговорот железа в океанах» . Микробиология Экология . 29 (1): 1. doi : 10.1111/j.1574-6941.1999.tb00593.x .
- ^ Мур, CM; Миллс, ММ; Арриго, КР; Берман-Франк, И.; Бопп, Л.; Бойд, П.В.; Гэлбрейт, Эд; Гейдер, Р.Дж.; Гье, К.; Жаккар, СЛ; Джикеллс, Т.Д.; Ла Рош, Дж.; Лентон, ТМ; Маховальд, Нью-Мексико; Мараньон, Э. (сентябрь 2013 г.). «Процессы и закономерности ограничения питательных веществ океана» . Природа Геонауки . 6 (9): 701–710. Бибкод : 2013NatGe...6..701M . дои : 10.1038/ngeo1765 . ISSN 1752-0894 . S2CID 249514 .
- ^ Брокер, WS; Пэн, Т; Бенг, З. (1982). Трейсеры в море . Геологическая обсерватория Ламонт-Доэрти. ISBN 9780961751104 .
- ^ Бжезинский, Марк А. (1985). «СООТНОШЕНИЕ Si:C:N МОРСКИХ ДИАТОМОВ: МЕЖВИДОВАЯ ИЗМЕНЧИВОСТЬ И ВЛИЯНИЕ НЕКОТОРЫХ ОКРУЖАЮЩИХ ПЕРЕМЕННЫХ1». Журнал психологии . 21 (3): 347–357. дои : 10.1111/j.0022-3646.1985.00347.x . ISSN 1529-8817 . S2CID 86568341 .
Внешние ссылки
[ редактировать ]- Джонсон, Закари. «Биогеохимия IV» (PDF) . Школа наук и технологий об океане и Земле Гавайского университета.
- Ленц, Дженнифер (2010). «Стехиометрия питательных веществ — соотношения Редфилда» (PDF) . Школа побережья и окружающей среды ЛГУ.
- П.Г. Фальковский; К.С. Дэвис (2004). «МОРСКАЯ БИОГЕОХИМИЯ: О ОТНОШЕНИЯХ РЕДПОЛЯ» . Неделя науки. Природа .
| Базы данных органов управления : Национальные |
|---|