Биогенный кремнезем
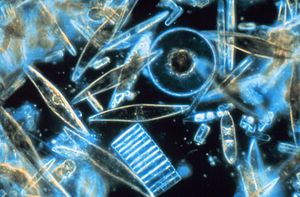
Биогенный кремнезем (bSi), также называемый опалом , биогенным опалом или аморфным опаловым кремнеземом , образует один из наиболее распространенных биогенных минералов . Например, микроскопические частицы кремнезема, называемые фитолитами, можно найти в травах и других растениях.
Кремнезем представляет собой аморфный оксид металлоида, образующийся в результате сложных неорганической полимеризации процессов . Это отличается от других основных биогенных минералов, включая карбонат и фосфат , которые встречаются в природе в виде кристаллических ионно-ковалентных твердых веществ (например, солей которых ), осаждение диктуется равновесием растворимости . [1] С химической точки зрения bSi представляет собой гидратированный кремнезем (SiO 2 · n H 2 O), который необходим многим растениям и животным.
Диатомовые водоросли как в пресной, так и в соленой воде извлекают из воды растворенный кремнезем и используют его в качестве компонента своих клеточных стенок. Аналогичным образом некоторые голопланктонные простейшие ( радилярии ), некоторые губки и некоторые растения ( фитолиты листьев ) используют кремний в качестве структурного материала. Известно, что кремний необходим цыплятам и крысам для роста и развития скелета. человека Кремний содержится в соединительных тканях , костях , зубах , коже , глазах , железах и органах .
Кремнезем в морской среде
[ редактировать ]Силикат , или кремниевая кислота (H 4 SiO 4 ), является важным питательным веществом в океане. В отличие от других основных питательных веществ, таких как фосфат , нитрат или аммоний , которые необходимы почти всему морскому планктону , силикат является важным химическим компонентом очень специфической биоты , включая диатомовые водоросли , радиолярии , силикофлагелляты и кремнистые губки . Эти организмы извлекают растворенный силикат из поверхностных вод открытого океана для накопления дисперсного кремнезема (SiO 2 ) или опалина в скелетных структурах (т.е. твердых частях биоты). [2] [3] Некоторые из наиболее распространенных кремниевых структур, наблюдаемых на поверхности клеток организмов, секретирующих кремнезем, включают: спикулы, чешуйки, твердые пластинки, гранулы, панцири и другие сложные геометрические формы, в зависимости от рассматриваемого вида. [4]
Морские источники кремнезема
[ редактировать ]Можно выделить пять основных источников растворенного кремнезема в морской среде: [3]
- Речной приток растворенного кремнезема в океаны: 4,2 ± 0,8 × 10. 14 г SiO 2 года −1
- Подводный вулканизм и связанные с ним гидротермальные эманации: 1,9 ± 1,0 × 10. 14 г SiO 2 года −1
- Ледниковое выветривание : 2 × 10 12 г SiO 2 года −1
- Низкотемпературное подводное выветривание океанических базальтов
- Некоторое количество кремнезема может также выходить из обогащенных кремнеземом поровых вод пелагических отложений на морском дне.
После гибели организма часть кремниевого скелетного материала растворяется , оседая в толще воды , обогащая глубокие воды растворенным кремнеземом. [3] Некоторые из кремнистых чешуек также могут сохраняться с течением времени в виде микроокаменелостей в глубоководных отложениях , открывая окно в современные и древние планктона / протистов сообщества . Этот биологический процесс, по крайней мере, с раннего палеозоя , регулировал баланс кремнезема в океане. [4]
Радиолярии ( кембрий / ордовик - голоцен ), диатомовые водоросли ( мел - голоцен ) и кремнефлагелляты ( мел - голоцен ) вносят основной вклад в глобальный биогенный цикл кремнезема на протяжении всего геологического времени . Диатомовые водоросли составляют 43% первичной продукции океана и ответственны за большую часть добычи кремнезема из океанских вод в современном океане на протяжении большей части последних пятидесяти миллионов лет. Напротив, океаны юрского и более древнего возраста характеризовались радиоляриями как основными типами, использующими кремнезем. [2] В настоящее время радиолярии являются вторыми (после диатомей) крупными продуцентами взвешенного аморфного кремнезема в водах океана. Их распространение колеблется от Арктики до Антарктики , наиболее распространено в экваториальной зоне. В экваториальных водах Тихого океана , например, можно наблюдать около 16 000 особей на кубический метр. [4]
Кремнеземный цикл
[ редактировать ]В последнее десятилетие кремниевый цикл все больше привлекает научное внимание по нескольким причинам:
Во-первых , широко распространено мнение, что в современном морском цикле кремнезема доминируют диатомовые водоросли, которые фиксируют и экспортируют твердые частицы (включая органический углерод ) из эвфотической зоны в глубокие глубины океана посредством процесса, известного как биологический насос . В результате диатомовые водоросли и другие организмы, выделяющие кремнезем, играют решающую роль в глобальном углеродном цикле и обладают способностью влиять на в атмосфере концентрацию CO 2 в различных временных масштабах, связывая CO 2 в океане. Эта связь между биогенным кремнеземом и органическим углеродом, а также значительно более высокий потенциал сохранности биогенных кремнистых соединений по сравнению с органическим углеродом делают записи накопления опалов очень интересными для палеоокеанографии и палеоклиматологии .
Во-вторых , накопление биогенного кремнезема на морском дне содержит много информации о том, где в океане происходило экспортное производство в масштабах времени от сотен до миллионов лет. По этой причине записи об отложениях опалов предоставляют ценную информацию о крупномасштабных океанографических реорганизациях в геологическом прошлом, а также о палеопродуктивности.
В-третьих , среднее время пребывания силикатов в океане составляет примерно 10 000–15 000 лет. Такое относительно короткое время пребывания делает концентрации и потоки океанических силикатов чувствительными к ледниковым / межледниковым возмущениям и, таким образом, является отличным показателем для оценки изменений климата. [3] [5]
Все чаще изотопные соотношения кислорода ( O 18 :THE 16 ) и кремний (Si 30 :И 28 ) анализируются на основе биогенного кремнезема, сохранившегося в озерных и морских отложениях, с целью получения данных о прошлых изменениях климата и круговороте питательных веществ (De La Rocha, 2006; Leng and Barker, 2006). Это особенно ценный подход, учитывая роль диатомовых водорослей в глобальном круговороте углерода. Кроме того, изотопный анализ BSi полезен для отслеживания прошлых изменений климата в таких регионах, как Южный океан мало биогенных карбонатов , где сохранилось .
Морские кварцевые раковины
[ редактировать ]Кремнистый ил
[ редактировать ]Остатки диатомей и других организмов, использующих кремнезем, встречаются в виде опаловых отложений в пелагических глубоководных отложениях. Пелагические отложения , содержащие значительные количества кремнистых биогенных остатков, принято называть кремнистым илом . Кремнистые илы особенно распространены в современном океане в высоких широтах северного и южного полушарий. Яркой особенностью распространения кремнистого ила является то, что ок. Пояс шириной 200 км, протянувшийся через Южный океан . Некоторые экваториальные районы апвеллинга , где питательные вещества обильны и продуктивность высока, также характеризуются местными кремнистыми илами. [2]
Кремнистые илы состоят преимущественно из остатков диатомей и радиолярий, но могут включать и другие кремнистые организмы, например силикофлагелляты и губок спикулы . Диатомовые илы встречаются преимущественно в высокоширотных районах и вдоль некоторых окраин континентов, тогда как радиоляриевые илы более характерны для экваториальных областей. Кремнистые илы видоизменяются и превращаются при погребении в слоистые кремни . [2]
Отложения Южного океана
[ редактировать ]Отложения Южного океана являются основным поглотителем биогенного кремнезема (50-75% общего содержания в океане 4,5 × 10 14 г SiO 2 года −1 ; DeMaster, 1981), но является лишь незначительным поглотителем органического углерода (<1% океанического 2 × 10 14 г органического C год −1 ). Столь относительно высокие темпы накопления биогенного кремнезема в отложениях Южного океана (преимущественно под Полярным фронтом) по отношению к органическому углероду (60:1 по массе) обусловлены преимущественным сохранением биогенного кремнезема в водной толще Антарктики.
В отличие от того, что считалось ранее, эти высокие темпы накопления биогенного кремнезема не являются результатом высоких темпов первичной продукции . Биологическое производство в Южном океане сильно ограничено из-за низкого уровня радиации в сочетании с глубокими перемешанными слоями и/или ограниченным количеством микроэлементов , таких как железо . [6]
Это предпочтительное сохранение биогенного кремнезема по сравнению с органическим углеродом проявляется в постоянно увеличивающемся соотношении кремнезема/органического углерода в зависимости от глубины в толще воды. Около тридцати пяти процентов биогенного кремнезема, образующегося в эвфотической зоне, переживает растворение в поверхностном слое; тогда как только 4% органического углерода избегает микробного разложения в этих приповерхностных водах.
Следовательно, во время осаждения через толщу воды происходит значительное разделение органического углерода и кремнезема. Накопление биогенного кремнезема на морском дне составляет 12% поверхностной продукции, тогда как скорость накопления органического углерода на морском дне составляет лишь <0,5% поверхностной продукции. В результате на полярные отложения приходится большая часть накопления биогенного кремнезема в океане, но лишь небольшое количество осадочного потока органического углерода. [6]
Влияние океанической циркуляции на поглотители кремнезема
[ редактировать ]Крупномасштабная океаническая циркуляция оказывает прямое влияние на отложение опалов . Циркуляции в Тихом океане (характеризуются бедными питательными веществами поверхностными водами и глубокими водами, богатыми питательными веществами) и Атлантическом океане способствуют производству / сохранению кремнезема и карбоната соответственно. Например, отношения Si/N и Si/P увеличиваются от Атлантики к Тихому и Южному океану, отдавая предпочтение производителям опалов, а не карбонатов . Следовательно, современная конфигурация крупномасштабной океанической циркуляции привела к локализации основных зон захоронения опалов в экваториальной части Тихого океана, в системах апвеллинга восточных пограничных течений и, что наиболее важно, в Южном океане. [5]
Тихий и Южный океаны
[ редактировать ]В водах современного Тихого и Южного океана обычно наблюдается увеличение отношения Si/N на средней глубине, что приводит к увеличению экспорта опалов (~увеличению производства опалов). В Южном океане и северной части Тихого океана эта зависимость между экспортом опала и соотношением Si/N меняется с линейной на экспоненциальную для отношений Si/N, превышающих 2. Это постепенное увеличение важности силиката (Si) по сравнению с азотом (N) привело к огромные последствия для биологического производства океана. Изменение соотношения питательных веществ способствует выбору диатомовых водорослей в качестве основных производителей по сравнению с другими (например, кальцифицирующими) организмами. Например, эксперименты на микромире показали, что диатомовые водоросли являются суперконкурентами DSi и доминируют над другими производителями с DSi выше 2 мкМ. Следовательно, будет предпочтительнее экспорт опала, а не карбоната, что приведет к увеличению производства опала. Южный океан и северная часть Тихого океана также демонстрируют максимальное соотношение биогенных силикатов к органическим потокам C и, таким образом, заключаются в обогащении биогенными силикатами по сравнению с C. органический экспортный поток. Такое совокупное увеличение сохранности и экспорта опалов делает Южный океан сегодня самым важным поглотителем для DSi. [5]
Атлантический океан
[ редактировать ]В Атлантическом океане промежуточные и глубокие воды характеризуются меньшим содержанием DSi по сравнению с современными Тихим и Южным океаном. Эта более низкая разница в DSi между бассейнами приводит к снижению потенциала сохранности опала в Атлантике по сравнению с его аналогами в Тихом и Южном океане. В водах, обедненных Атлантическим DSi, наблюдается тенденция к образованию относительно менее окремненных организмов, что сильно влияет на сохранность их панцирей . Этот механизм лучше всего иллюстрируется при сравнении систем апвеллинга Перу и северо-западной Африки. Соотношение растворение /производство при атлантическом апвеллинге намного выше, чем при тихоокеанском апвеллинге. Это связано с тем, что воды прибрежного апвеллинга значительно богаче DSi у берегов Перу, чем у северо-западной Африки. [5]
Морской биогенный баланс кремнезема
[ редактировать ]Реки и подводные гидротермальные излучения поставляют 6,1 × 10 14 г SiO 2 года −1 к морской среде. Примерно две трети этого поступления кремнезема хранится на окраинах континентов и в глубоководных отложениях. Кремнистые глубоководные отложения, расположенные под Антарктической конвергенцией ( зоной конвергенции ), содержат около 25% кремнезема, поступающего в океаны (т.е. 1,6 × 10 14 г SiO 2 года −1 ) и, следовательно, образуют один из крупнейших поглотителей кремнезема на Земле. Самые высокие скорости накопления биогенного кремнезема в этом районе наблюдаются в Южной Атлантике, их значения достигают 53 см·тысяч лет. −1 в течение последних 18 000 лет. Кроме того, обширное накопление биогенного кремнезема зарегистрировано в глубоководных отложениях Берингова моря , Охотского моря и субарктической северной части Тихого океана . Суммарные скорости накопления биогенного кремнезема в этих регионах составляют около 0,6 × 10 14 г SiO 2 года −1 , что эквивалентно 10% растворенного кремнезема, поступающего в океаны.
Континентальные окраинные районы апвеллинга, такие как Калифорнийский залив , побережье Перу и Чили, характеризуются одними из самых высоких темпов накопления биогенного кремнезема в мире. Например, скорость накопления биогенного кремнезема составляет 69 г SiO 2 /см. 2 /kyr были зарегистрированы в Калифорнийском заливе. Из-за ограниченного по латерали характера этих зон быстрого биогенного накопления кремнезема только на области апвеллинга приходится около 5% растворенного кремнезема, поступающего в океаны. Наконец, в обширных глубоководных отложениях Атлантического, Индийского и Тихого океанов наблюдаются чрезвычайно низкие темпы накопления биогенного кремнезема, что делает эти океаны незначительными для глобального баланса морского кремнезема. [7]
Производство биогенного кремнезема
[ редактировать ]
Среднесуточный курс BSi сильно зависит от региона:
- Прибрежный апвеллинг : 46 ммоль.м. −2 .д −1
- Субарктическая часть Тихого океана: 18 ммоль·м. −2 .д −1
- Южный океан: 3–38 ммоль·м. −2 .д −1
- срединно-океанические круговороты : 0,2–1,6 ммоль·м. −2 .д −1
Аналогичным образом, интегрированное годовое производство BSi сильно зависит от региона:
- Прибрежный апвеллинг: 3 × 10 12 моль −1
- Субарктическая часть Тихого океана: 8 × 10 12 моль −1
- Южный океан: 17–37 × 10 12 моль −1
- срединно-океанические круговороты: 26 × 10 12 моль −1
Производство BSi контролируется:
- Однако наличие растворенного кремнезема , константа полунасыщения Kμ для роста, ограниченного кремнием, ниже, чем Ks для поглощения кремния.
- Наличие света: нет требований к прямому освещению; поглощение кремния на 2-кратной глубине фотосинтеза ; Поглощение кремния продолжается ночью, но клетки должны активно расти.
- Доступность микроэлементов .
Растворение биогенного кремнезема
[ редактировать ]Растворение BSi контролируется:
- Термодинамика растворимости : Температура (от 0 до 25 °C – 50- кратное увеличение).
- Скорость погружения : Структура пищевой сети — травоядные, фекальные гранулы, выброшенные кормовые структуры. Агрегация — быстрый транспорт.
- Бактериальная деградация ( органического матрикса Бидл и Азам, 1999).
Биогенная консервация кремнезема
[ редактировать ]Сохранность BSi измеряется:
- Скорость седиментации, в основном отстойники (Хондзё);
- бентоса Скорость реминерализации («рециклинг»), камера донного потока (Берельсон);
- Концентрация BSi в отложениях, химическое выщелачивание в щелочном растворе , зависит от конкретного участка, необходимо различать литогенный и биогенный Si, дифракция рентгеновских лучей .
Сохранность BSi контролируется:
- Скорость седиментации;
- Концентрация растворенного кремнезема в поровой воде : насыщение 1100 мкмоль/л;
- Поверхностные покрытия: растворенный Al изменяет растворимость осажденных частиц биогенного кремнезема, растворенный кремнезем также может осаждаться с Al в виде глины или покрытий Al-Si.
Опалиновый кремнезем на Марсе
[ редактировать ]В кратере Гусева на Марсе марсоход Spirit случайно обнаружил опаловый кремнезем. Одно из его колес ранее было обездвижено и, таким образом, фактически врезалось в марсианский реголит , пока тот тащился за перемещающимся марсоходом. Более поздний анализ показал, что кремнезем свидетельствует о гидротермальных условиях. [8]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Корадин Т., Лопес П.Дж. (2003). «Создание биогенного кремнезема: простая химия или тонкая биология?» ХимБиоХим 3: 1-9.
- ^ Jump up to: а б с д Боггс, С. (2005). «Принципы седиментологии и стратиграфии (4-е издание)». Pearson Education, Inc, 662 стр.
- ^ Jump up to: а б с д ДеМастер, ди-джей (1981). «Поставка и накопление кремнезема в морской среде». Geochimica et Cosmochimica Acta 45: 1715–1732.
- ^ Jump up to: а б с Эрлих и др. (2010). «Современные взгляды на десилицирование: растворение биокремнезема и абиотического кремнезема в естественной и искусственной среде». хим. Откр. 110: 4656-4689.
- ^ Jump up to: а б с д Кортезе Г., Жерсонд Р. (2004). «Сдвиги осадконакопления опалов в Мировом океане за последние 15 млн лет». Письма о Земле и планетарной науке 224: 509–527.
- ^ Jump up to: а б ДеМастер, Д. (1992). «Круговорот и накопление биогенного кремнезема и органических веществ в высоких широтах: море Росса». Океанография 5(3): 147-153.
- ^ ДеМастер, ди-джей (2002). «Накопление и круговорот биогенного кремнезема в Южном океане: новый взгляд на баланс морского кремнезема». Глубоководные исследования, часть II 49: 3155-3167
- ^ [1] Рафф, SW и др. (2011). «Характеристики, распределение, происхождение и значение опалового кремнезема, наблюдаемого марсоходом Spirit в кратере Гусева на Марсе». Дж. Геофиз. Рез., 116, E00F23.
- Бжезинский, Массачусетс (1985). «Соотношение Si:C:N морских диатомей: межвидовая изменчивость и влияние некоторых переменных окружающей среды». Журнал Phycology 21 (3): 347-357.
- Де Ла Роча, CL (2006). «Основные данные о палеоэкологических условиях на основе опала». Глобальные биогеохимические циклы 20. дои : 10.1029/2005GB002664 .
- Дагдейл, Р.К. и Ф.П. Вилкерсон (1998). «Силикатное регулирование новой продукции в экваториальном тихоокеанском апвеллинге». Природа 391(6664): 270.
- Дагдейл, Р.К., Ф.П. Вилкерсон и др. (1995). «Роль силикатного насоса в стимулировании нового производства». Глубоководные исследования I 42 (5): 697-719.
- Ленг, М.Дж. и Баркер, Пенсильвания (2006). «Обзор изотопного состава кислорода озерного диатомового кремнезема для реконструкции палеоклимата». Обзоры наук о Земле 75:5-27.
- Рагено О., П. Трегер и др. (2000). «Обзор цикла кремния в современном океане: недавний прогресс и недостающие пробелы в применении биогенного опала в качестве показателя палеопродуктивности». Глобальные и планетарные изменения 26: 317-365.
- Такеда, С. (1998). «Влияние доступности железа на соотношение потребления питательных веществ диатомовыми водорослями в океанических водах». Природа 393: 774-777.
- Вернер, Д. (1977). Биология диатомей. Беркли и Лос-Анджелес, Калифорнийский университет Press.