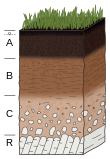
Дыхание почвы
| Часть серии о |
| Углеродный цикл |
|---|
 |
Под дыханием почвы понимается выработка углекислого газа при дыхании почвенных организмов . Сюда входит дыхание корней растений , ризосферы , микробов и фауны .
Дыхание почвы – это ключевой экосистемный процесс, который высвобождает углерод из почвы в форме CO 2 . CO 2 усваивается растениями из атмосферы и превращается в органические соединения в процессе фотосинтеза . Растения используют эти органические соединения для создания структурных компонентов или вдыхают их для высвобождения энергии. Когда дыхание растений происходит под землей, в корнях, оно дополняет дыхание почвы. С течением времени структурные компоненты растений потребляются гетеротрофами . При этом гетеротрофном потреблении выделяется CO 2 , и когда этот CO 2 выделяется подземными организмами, это считается дыханием почвы.
Количество дыхания почвы, происходящее в экосистеме, контролируется несколькими факторами. Температура, влажность, содержание питательных веществ и уровень кислорода в почве могут вызывать чрезвычайно разную скорость дыхания. Частоту дыхания можно измерить различными методами. Для разделения исходных компонентов, в данном случае типа пути фотосинтеза ( С3 / С4 ), дышащих растительных структур можно использовать и другие методы.
На скорость дыхания почвы может в значительной степени влиять деятельность человека. Это связано с тем, что люди обладают способностью изменять различные факторы, контролирующие дыхание почвы, и уже многие годы меняют их. Глобальное изменение климата состоит из множества изменяющихся факторов, включая повышение уровня CO 2 в атмосфере , повышение температуры и изменение характера осадков . Все эти факторы могут повлиять на скорость глобального дыхания почвы. Увеличение азотных внесения удобрений людьми также потенциально может повлиять на показатели по всей планете .
Дыхание почвы и его скорость в экосистемах чрезвычайно важны для понимания. Это связано с тем, что дыхание почвы играет большую роль в глобальном круговороте углерода , а также в других круговоротах питательных веществ . Дыхание растительных структур выделяет не только CO 2 , но и другие питательные вещества в этих структурах, например азот. Дыхание почвы также связано с положительной обратной связью с глобальным изменением климата. Положительная обратная связь – это когда изменение в системе вызывает реакцию в том же направлении, что и изменение. Таким образом, изменение климата может повлиять на скорость дыхания почвы, а затем отреагировать усилением изменения климата.
Источники углекислого газа в почве
[ редактировать ]
Любое клеточное дыхание высвобождает энергию, воду и CO 2 из органических соединений. Любое дыхание, происходящее под землей, считается дыханием почвы. Дыхание корней растений, бактерий, грибов и почвенных животных приводит к выделению CO 2 в почву, как описано ниже.
Цикл трикарбоновых кислот (ТСА)
[ редактировать ]Цикл трикарбоновых кислот (ТСА) – или цикл лимонной кислоты – является важным этапом клеточного дыхания. шестиуглеродный сахар В цикле ТСА окисляется . [1] образуются CO 2 и H 2 В результате этого окисления из сахара O. Растения, грибы, животные и бактерии используют этот цикл для преобразования органических соединений в энергию. Именно так происходит большая часть дыхания почвы на самом базовом уровне. Поскольку для возникновения этого процесса требуется кислород, его называют аэробным дыханием.
Ферментация
[ редактировать ]Ферментация — это еще один процесс, в котором клетки получают энергию из органических соединений. В этом метаболическом пути энергия получается из соединений углерода без использования кислорода. Продуктами этой реакции являются углекислый газ и обычно либо этиловый спирт , либо молочная кислота . [2] Из-за недостатка кислорода этот путь называют анаэробным дыханием . Это важный источник CO 2 при дыхании почвы в заболоченных экосистемах с недостатком кислорода, например, в торфяных болотах и водно-болотных угодьях . Однако большая часть CO 2 высвобождается из почвы в результате дыхания, и один из наиболее важных аспектов подземного дыхания происходит в корнях растений.
Корневое дыхание
[ редактировать ]Растения дышат некоторыми соединениями углерода, образующимися в результате фотосинтеза. Когда это дыхание происходит в корнях, оно дополняет дыхание почвы. На корневое дыхание приходится примерно половина всего дыхания почвы. Однако эти значения могут варьироваться от 10 до 90% в зависимости от доминирующих типов растений в экосистеме и условий, которым подвергаются растения. Таким образом, количество CO 2 , выделяемого при дыхании корней, определяется биомассой корней и удельной скоростью дыхания корней. [3] Непосредственно рядом с корнем находится область, известная как ризосфера, которая также играет важную роль в дыхании почвы.
Ризосферное дыхание
[ редактировать ]Ризосфера . – это зона, непосредственно примыкающая к поверхности корня с соседней почвой В этой зоне происходит тесное взаимодействие растения и микроорганизмов. Корни непрерывно выделяют вещества или экссудат в почву. Эти экссудаты включают сахара, аминокислоты , витамины , длинноцепочечные углеводы , ферменты и лизаты , которые высвобождаются при разрушении клеток корня. Количество углерода, теряемого с экссудатами, значительно варьируется в зависимости от вида растений. Показано, что до 20% углерода, полученного в результате фотосинтеза, выделяется в почву в виде корневых выделений. [4] Эти экссудаты разлагаются преимущественно бактериями. Эти бактерии будут вдыхать соединения углерода в цикле ТСА; однако брожение также присутствует. Это связано с недостатком кислорода из-за большего потребления кислорода корнем по сравнению с объемной почвой, почвой, находящейся на большем расстоянии от корня. [5] Другим важным организмом в ризосфере являются грибы, поражающие корни, или микоризы . Эти грибы увеличивают площадь поверхности корня растения и позволяют корню встречать и усваивать большее количество питательных веществ почвы, необходимых для роста растений. В обмен на эту выгоду растение будет передавать сахар грибам. Грибы будут вдыхать эти сахара для получения энергии, тем самым увеличивая дыхание почвы. [6] Грибы, наряду с бактериями и почвенными животными, также играют большую роль в разложении подстилки и органического вещества почвы .
Почвенные животные
[ редактировать ]Почвенные животные питаются популяциями бактерий и грибов, а также заглатывают и расщепляют мусор, чтобы улучшить дыхание почвы. Микрофауна состоит из мельчайших почвенных животных. К ним относятся нематоды и клещи . Эта группа специализируется на почвенных бактериях и грибах. Поглощая эти организмы, углерод, который изначально находился в органических соединениях растений и был включен в бактериальные и грибковые структуры, теперь будет вдыхаться почвенными животными. Мезофауна - это почвенные животные длиной от 0,1 до 2 миллиметров (от 0,0039 до 0,0787 дюйма), которые питаются почвенным мусором. Фекальный материал будет удерживать большее количество влаги и иметь большую площадь поверхности. Это позволит обеспечить новую атаку микроорганизмов и увеличить объем дыхания почвы. Макрофауна — это организмы размером от 2 до 20 миллиметров (от 0,079 до 0,787 дюйма), такие как дождевые черви и термиты . Большая часть макрофауны фрагментирует подстилку, тем самым подвергая большую территорию микробному нападению. Другая макрофауна роет норы или заглатывает мусор, уменьшая объемную плотность почвы, разбивая почвенные агрегаты и увеличивая аэрацию почвы и проникновение воды. [7]
Регуляция дыхания почвы
[ редактировать ]Регулирование производства CO 2 в почве происходит за счет различных абиотических , или неживых, факторов. Температура, влажность почвы и азот влияют на скорость дыхания в почве.
Температура
[ редактировать ]
Температура влияет практически на все стороны процессов дыхания. Температура будет экспоненциально увеличивать дыхание до максимума, после чего дыхание уменьшится до нуля, когда ферментативная активность прерывается. Дыхание корней увеличивается экспоненциально с увеличением температуры в ее низком диапазоне, когда скорость дыхания ограничивается в основном циклом ТСА. При более высоких температурах ограничивающим фактором становится транспорт сахаров и продуктов обмена. При температуре выше 35 °C (95 °F) корневое дыхание начинает полностью прекращаться. [8] Микроорганизмы делятся на три температурные группы; криофилы , мезофилы и термофилы . Криофилы оптимально функционируют при температуре ниже 20 °C (68 °F), мезофилы лучше всего функционируют при температуре от 20 до 40 °C (104 °F), а термофилы оптимально функционируют при температуре выше 40 °C (104 °F). В естественных почвах существует множество различных когорт или групп микроорганизмов. Все эти когорты будут лучше всего функционировать в разных условиях, поэтому дыхание может происходить в очень широком диапазоне. [9] Повышение температуры приводит к увеличению скорости дыхания почвы, пока высокие значения не замедляют микробную функцию. Это та же самая закономерность, которая наблюдается при изменении уровня влажности почвы.
Влажность почвы
[ редактировать ]Влажность почвы – еще один важный фактор, влияющий на дыхание почвы. Дыхание почвы низкое в засушливых условиях и увеличивается до максимума при промежуточном уровне влажности, пока не начинает уменьшаться, когда содержание влаги исключает кислород. Это позволяет преобладать анаэробным условиям и подавлять аэробную микробную активность. Исследования показали, что влажность почвы ограничивает дыхание только в самых низких и самых высоких условиях, при этом для большинства экосистем существует большое плато при промежуточных уровнях влажности почвы. [10] Многие микроорганизмы обладают стратегиями роста и выживания в условиях низкой влажности почвы. В условиях высокой влажности почвы многие бактерии поглощают слишком много воды, что приводит к лизису или разрыву их клеточных мембран. Это может временно снизить скорость дыхания почвы, но лизис бактерий приводит к резкому увеличению ресурсов для многих других бактерий. Такое быстрое увеличение количества доступных лабильных субстратов вызывает кратковременное усиление дыхания почвы. Корневое дыхание будет усиливаться с увеличением влажности почвы, особенно в засушливых экосистемах; однако реакция корневого дыхания отдельных видов на влажность почвы будет широко варьироваться от вида к виду в зависимости от особенностей жизненного цикла. Повышенная влажность почвы угнетает дыхание корней, ограничивая доступ атмосферного кислорода. За исключением водно-болотных растений, у которых развились особые механизмы корневой аэрации, большинство растений не адаптировано к водно-болотной почвенной среде с низким содержанием кислорода . [11] Эффект подавления дыхания повышенной влажности почвы усиливается, когда дыхание почвы также снижает окислительно-восстановительный потенциал почвы посредством биоэлектрогенеза . [12] Почвенные микробные топливные элементы становятся популярными образовательными инструментами в научных классах.
Азот
[ редактировать ]Азот напрямую влияет на дыхание почвы несколькими способами. Азот должен усваиваться корнями, чтобы способствовать росту и жизни растений. Наиболее доступный азот находится в форме NO3 . − , для проникновения которого в корень требуется 0,4 единицы CO 2 необходимо использовать энергию , поскольку для перемещения его вверх по градиенту концентрации . Попав в корень, NO 3 − должно быть уменьшено до NH 3 . Этот шаг требует больше энергии, что соответствует 2 единицам CO 2 на восстановленную молекулу. У растений с бактериальными симбионтами , фиксирующими атмосферный азот, энергетические затраты растения на приобретение одной молекулы NH 3 из атмосферного N 2 составляют 2,36 CO 2 . [13] Очень важно, чтобы растения поглощали азот из почвы или полагались на симбионтов, которые усвоили его из атмосферы, чтобы обеспечить рост, размножение и долгосрочное выживание.
Другой способ воздействия азота на дыхание почвы – разложение подстилки . Подстилка с высоким содержанием азота считается высококачественной и легче разлагается микроорганизмами, чем подстилка низкого качества. Разложение целлюлозы , прочного структурного соединения растений, также является процессом, ограниченным азотом, и будет усиливаться с добавлением азота в подстилку. [14]
Методы измерения
[ редактировать ]Существуют различные методы измерения скорости дыхания почвы и определения источников. Методы можно разделить на полевые и лабораторные. Наиболее распространенные полевые методы включают использование долговременных автономных систем почвенных потоков для измерений в одном месте в разное время; обследовать системы дыхания почвы для измерения в разных местах и в разное время. Использование соотношений стабильных изотопов может быть использовано как в лабораторных, так и в полевых измерениях.
Дыхание почвы можно измерять отдельно или с добавлением питательных веществ и (углеродных) субстратов, которые служат источниками питания для микроорганизмов. Дыхание почвы без каких-либо добавок питательных веществ и субстратов называется базальным дыханием почвы (БД). С добавлением питательных веществ (часто азота и фосфора) и субстратов (например, сахаров) это называется субстрат-индуцированным дыханием почвы (SIR). При измерениях BR и SIR содержание влаги можно регулировать с помощью воды.
Полевые методы
[ редактировать ]Автономные системы почвенного потока для долгосрочного использования для измерения в одном месте с течением времени
[ редактировать ]
Эти системы производят измерения в одном месте в течение длительных периодов времени. Поскольку они измеряют только в одном месте, обычно используют несколько станций, чтобы уменьшить ошибку измерения, вызванную изменчивостью почвы на небольших расстояниях. Изменчивость почвы можно проверить с помощью приборов для исследования дыхания почвы.
Приборы длительного действия спроектированы таким образом, чтобы место измерения подвергалось максимальному воздействию условий окружающей среды между измерениями.
Виды долгосрочных автономных инструментов
[ редактировать ]Закрытые, нестационарные системы
[ редактировать ]Закрытые системы проводят кратковременные измерения (обычно в течение нескольких минут) в камере, закрытой над почвой. [15] Скорость истечения CO 2 из почвы рассчитывается на основе увеличения CO 2 внутри камеры. в закрытых камерах Поскольку CO 2 продолжает накапливаться, периоды измерений сокращаются до минимума, чтобы добиться заметного линейного увеличения концентрации, избегая чрезмерного накопления CO 2 внутри камеры с течением времени.
об индивидуальном анализе как информация , так и информация о суточном измерении дыхания CO 2 Доступна . Такие системы также часто измеряют температуру почвы, влажность почвы и ФАР ( фотосинтетически активное излучение ). Эти переменные обычно записываются в файл измерений вместе со значениями CO 2 .
Для определения дыхания почвы и наклона увеличения CO 2 исследователи использовали линейный регрессионный анализ, алгоритм Педерсена (2001) и экспоненциальную регрессию . Есть больше опубликованных ссылок на анализ линейной регрессии; однако алгоритм Педерсена и методы экспоненциального регрессионного анализа также имеют своих последователей. Некоторые системы предлагают выбор математических методов. [16]
При использовании линейной регрессии несколько точек данных отображаются на графике, и точки могут быть сопоставлены с уравнением линейной регрессии, которое обеспечит наклон. Этот уклон может определить скорость дыхания почвы по уравнению , где F — скорость дыхания почвы, b — уклон, V — объем камеры, А — площадь поверхности почвы, покрытая камерой. [17] Важно, чтобы измерение не продолжалось в течение более длительного периода времени, поскольку увеличение концентрации CO 2 в камере также приведет к увеличению концентрации CO 2 в пористом верхнем слое профиля почвы. Это увеличение концентрации приведет к недооценке скорости дыхания почвы из-за дополнительного накопления CO 2 в почве. [18]

Открытые устойчивые системы
[ редактировать ]Системы открытого режима предназначены для определения скорости потока почвы при достижении равновесия в измерительной камере. Воздух проходит через камеру, прежде чем камера закрывается и герметизируется. Это удаляет CO 2 из камеры все уровни , не относящиеся к окружающей среде, перед измерением. После закрытия камеры в нее подается свежий воздух с контролируемой и программируемой скоростью потока. Он смешивается с CO 2 из почвы, и через некоторое время достигается равновесие. Исследователь определяет точку равновесия как разницу в измерениях CO 2 между последовательными показаниями за истекший период времени. Во время анализа скорость изменения медленно снижается до тех пор, пока не будет соответствовать критериям скорости изменения клиента или максимально выбранному времени для анализа. Затем определяется поток почвы или скорость изменения, как только внутри камеры достигаются условия равновесия. Скорость и время потока в камере программируются, точно измеряются и используются в расчетах. Эти системы имеют вентиляционные отверстия, предназначенные для предотвращения возможного неприемлемого накопления частичного CO. 2 давление обсуждается в системах закрытого типа. Поскольку движение воздуха внутри камеры может вызвать повышение давления в камере, а внешние ветры могут привести к снижению давления в камере, предусмотрено вентиляционное отверстие, максимально защищенное от ветра.
Открытые системы также не так чувствительны к изменениям структуры почвы или к проблемам сопротивления пограничного слоя на поверхности почвы. Поток воздуха в камере у поверхности почвы предназначен для минимизации явлений сопротивления пограничного слоя.
Системы гибридного режима
[ редактировать ]Существует также гибридная система. Он имеет вентиляционное отверстие, которое спроектировано так, чтобы быть максимально защищенным от ветра и предотвращать возможное неприемлемое частичное повышение давления CO 2 , но в других отношениях оно предназначено для работы как система проектирования закрытого режима.
Исследование систем дыхания почвы – для проверки изменений дыхания CO 2 в разных местах и в разное время.
[ редактировать ]
Это портативные или полупортативные инструменты открытого или закрытого типа. Они измеряют изменчивость дыхания почвы CO 2 в разных местах и в разное время. С помощью этого типа инструмента почвенные воротники, которые можно подключить к геодезическому измерительному прибору, вставляются в землю, и почве дают стабилизироваться в течение определенного периода времени. Установка почвенного воротника временно нарушает почву, создавая артефакты измерения. По этой причине обычно в разных местах вставляют несколько почвенных воротников. Почвенные воротники вставляются достаточно глубоко, чтобы ограничить боковую диффузию CO 2 . После стабилизации почвы исследователь переходит от одного воротника к другому в соответствии с планом эксперимента для измерения дыхания почвы.
Системы исследования дыхания почвы также могут использоваться для определения количества долговременных автономных временных инструментов, которые необходимы для достижения приемлемого уровня погрешности. В разных местах может потребоваться разное количество автономных установок длительного действия из-за большей или меньшей изменчивости дыхания почвы.
Изотопные методы
[ редактировать ]Растения усваивают CO 2 и производят органические соединения, используя один из трех путей фотосинтеза . Двумя наиболее распространенными путями являются процессы C3 и C4 . Растения C 3 лучше всего приспособлены к прохладным и влажным условиям, а растения C 4 хорошо себя чувствуют в жарких и засушливых экосистемах. Из-за разных ферментов фотосинтеза в этих двух путях разные изотопы углерода преимущественно приобретаются . Изотопы — это один и тот же элемент, которые различаются числом нейтронов, в результате чего один изотоп становится тяжелее другого. Два стабильных изотопа углерода: 12 С и 13 C. Путь C3 будет дискриминировать более тяжелый изотоп в большей степени, чем путь C4 . Это сделает растительные структуры, полученные из растений C 4 , более обогащенными более тяжелым изотопом, и, следовательно, корневые экссудаты и опад этих растений также будут более обогащены. Когда углерод в этих структурах вдыхается, CO 2 будет иметь аналогичное соотношение двух изотопов. Исследователи будут выращивать растение C 4 на почве, которая ранее была занята растением C 3 , или наоборот. Путем измерения дыхания почвы и анализа изотопных соотношений CO 2 можно определить, является ли дыхание почвы преимущественно старым или недавно образовавшимся углеродом. Например, кукуруза, растение C 4 , выращивалась на почве, где ранее выращивалась яровая пшеница , растение C 3 . Результаты показали дыхание C 3 SOM в первые 40 дней с постепенным линейным увеличением обогащения тяжелыми изотопами до 70-го дня. Дни после 70 показали замедление обогащения до пика на 100-й день. [19] Анализируя данные о стабильных изотопах углерода, можно определить исходные компоненты вдыхаемого ПОВ, образующегося различными путями фотосинтеза.
Субстратно-индуцированное дыхание в полевых условиях с использованием стабильных изотопов
[ редактировать ]Одна из проблем измерения дыхания почвы в полевых условиях заключается в том, что дыхание микроорганизмов невозможно отличить от дыхания корней растений и почвенных животных. Эту проблему можно решить, используя методы стабильных изотопов. Тростниковый сахар – это сахар C 4 , который может действовать как изотопный индикатор. [20] [21] Тростниковый сахар имеет несколько более высокое содержание 13 С (д 13 C ≈ −10‰), чем эндогенный (природный) углерод в экосистеме C 3 (δ 13 C=от -25 до -28‰). Тростниковый сахар можно распылить на почву в виде раствора, и он проникнет в верхний слой почвы. Только микроорганизмы будут вдыхать добавленный сахар, поскольку корни вдыхают исключительно углеродные продукты, которые усваиваются растением посредством фотосинтеза. Анализ δ 13 С CO 2 долю C 3 (корневого и микробного) и C 4 (микробного дыхания). , выделяющегося из почвы с добавлением тростникового сахара или без него, можно рассчитать [22] [23]
Полевое дыхание с использованием стабильных изотопов можно использовать в качестве инструмента для измерения микробного дыхания на месте, не нарушая микробные сообщества путем смешивания питательных веществ почвы, кислорода и почвенных загрязнителей, которые могут присутствовать. [23]
Реакция на человеческое беспокойство
[ редактировать ]За последние 160 лет люди изменили землепользование и промышленную практику, что изменило климат и глобальные биогеохимические циклы . Эти изменения повлияли на скорость дыхания почвы по всей планете. Кроме того, все более частые экстремальные климатические явления [24] такие как волны тепла (связанные с высокими температурными колебаниями и связанными с ними сильными засухами), сопровождаемые интенсивными дождями, оказывают воздействие на микробные сообщества и физико-химические свойства почвы и могут вызывать изменения в дыхании почвы. [25]
Повышенное содержание углекислого газа
[ редактировать ]Со времени промышленной революции люди выбросили огромное количество CO2 в атмосферу . Эти выбросы значительно увеличились с течением времени и привели к увеличению глобального уровня CO 2 в атмосфере до самого высокого уровня за более чем 750 000 лет. Дыхание почвы усиливается, когда экосистемы подвергаются воздействию повышенного уровня CO 2 . Многочисленные исследования по обогащению CO 2 в свободном воздухе (FACE) были проведены для проверки дыхания почвы в прогнозируемых в будущем условиях повышенного содержания CO 2 . Недавние исследования FACE показали значительное увеличение дыхания почвы из-за увеличения биомассы корней и микробной активности. [26] Было обнаружено, что дыхание почвы увеличивается до 40,6% в лесах сладкой камеди в Теннесси и тополевых лесах в Висконсине в условиях повышенного содержания CO 2 . [27] Весьма вероятно, что уровни CO 2 превысят уровни, используемые в этих экспериментах FACE, к середине этого столетия из-за увеличения использования человеком ископаемого топлива и практики землепользования.
Потепление климата
[ редактировать ]Из-за повышения температуры почвы уровень CO 2 в нашей атмосфере увеличивается, и поэтому средняя средняя температура Земли повышается. Это связано с деятельностью человека, такой как вырубка лесов почвы , оголение и событиями, разрушающими автотрофные процессы. С исчезновением фотосинтезирующих растений, покрывающих и охлаждающих поверхность почвы, инфракрасная энергия проникает в почву, нагревая ее и вызывая рост гетеротрофных бактерий. Гетеротрофы в почве быстро разлагают органическое вещество, структура почвы крошится, в результате чего она растворяется в ручьях и реках в море. Большая часть органических веществ, сметаемых наводнениями, вызванными вырубкой лесов, попадает в устья рек , водно-болотные угодья и, в конечном итоге, в открытый океан. Повышенная мутность поверхностных вод вызывает биологическую потребность в кислороде и гибель большего количества автотрофных организмов. Уровень углекислого газа повышается с усилением дыхания почвенных бактерий после повышения температуры из-за потери почвенного покрова.
Как уже говорилось ранее, температура сильно влияет на скорость дыхания почвы. Это может иметь самое сильное влияние в Арктике . Большие запасы углерода заключены в замерзшей вечной мерзлоте . С повышением температуры эта вечная мерзлота тает и начинают преобладать аэробные условия, тем самым значительно увеличивая скорость дыхания в этой экосистеме. [28]
Изменения количества осадков
[ редактировать ]Ожидается, что из-за изменения температуры и изменения условий океана характер осадков изменится по местоположению, частоте и интенсивности. Ожидаются более крупные и частые штормы, когда океаны смогут передавать больше энергии формирующимся штормовым системам. Это может оказать наибольшее воздействие на ксерические или засушливые экосистемы. Показано, что дыхание почвы в аридных экосистемах динамично меняется в пределах дождевого цикла . Скорость дыхания в сухой почве обычно достигает очень высокого уровня после дождя, а затем постепенно снижается по мере высыхания почвы. [10] При увеличении частоты и интенсивности осадков на территории без предшествующих обильных осадков можно сделать вывод о резком увеличении дыхания почвы.
Азотные удобрения
[ редактировать ]С началом « зеленой революции» в середине прошлого века было произведено огромное количество азотных удобрений, которые были внедрены практически во все системы сельского хозяйства. Это привело к увеличению доступного для растений азота в экосистемах по всему миру из-за сельскохозяйственных стоков и ветровых удобрений . Как обсуждалось ранее, азот может оказывать существенное положительное влияние на уровень и скорость дыхания почвы. Было обнаружено, что увеличение содержания азота в почве увеличивает темновое дыхание растений, стимулирует определенную скорость дыхания корней и увеличивает общую биомассу корней. [29] Это связано с тем, что высокие нормы азота связаны с высокими темпами роста растений. Высокие темпы роста растений приведут к увеличению дыхания и биомассы, выявленным в исследовании. При таком увеличении продуктивности может быть обеспечено увеличение активности почвы и, следовательно, дыхания.
Важность
[ редактировать ]Дыхание почвы играет важную роль в глобальных циклах углерода и питательных веществ, а также является движущей силой изменений климата. Эти роли важны для нашего понимания мира природы и сохранения человека.
Глобальный углеродный цикл
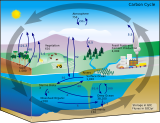
[ редактировать ]Дыхание почвы играет решающую роль в регулировании круговорота углерода на уровне экосистемы и в глобальном масштабе. Ежегодно около 120 петаграмм (Пг) углерода поглощается наземными растениями и такое же количество выбрасывается в атмосферу в результате дыхания экосистемы. Почвы мира содержат до 3150 Пг углерода, из которых 450 Пг приходится на водно-болотные угодья и 400 Пг — в вечномерзлые почвы. Почвы содержат в четыре раза больше углерода, чем атмосфера. [30] Исследователи подсчитали, что на дыхание почвы приходится 77 пг углерода, выбрасываемого в атмосферу каждый год. [31] Этот уровень выбросов превышает выбросы углерода из-за антропогенных источников (56 Пг в год), таких как сжигание ископаемого топлива . Таким образом, небольшое изменение в дыхании почвы может серьезно изменить баланс концентрации CO 2 в атмосфере и запасов углерода в почве . Подобно тому, как дыхание почвы может играть значительную роль в глобальном углеродном цикле, оно также может регулировать глобальный круговорот питательных веществ .
Цикл питательных веществ
[ редактировать ]Основным компонентом дыхания почвы является разложение подстилки, в результате которого выделяется CO 2 в окружающую среду и одновременно иммобилизуются или минерализуются питательные вещества. Во время разложения питательные вещества, такие как азот, иммобилизуются микробами для их собственного роста. Когда эти микробы попадают в организм или умирают, в почву добавляется азот. Азот также минерализуется в результате распада белков и нуклеиновых кислот в подстилке. Этот минерализованный азот также добавляется в почву. Благодаря этим процессам скорость поступления азота в почву связана со скоростью микробного дыхания. Исследования показали, что скорость дыхания почвы связана со скоростью микробного круговорота и минерализацией азота. [5] Изменения глобальных циклов могут в дальнейшем привести к изменению климата планеты.
Изменение климата
[ редактировать ]Как указывалось ранее, CO 2, выделяемый в результате дыхания почвы, является парниковым газом , который будет продолжать улавливать энергию и повышать глобальную среднюю температуру, если концентрации продолжат расти. По мере повышения глобальной температуры будет увеличиваться и скорость дыхания почвы по всему земному шару, что приведет к повышению концентрации CO 2 в атмосфере, что снова приведет к повышению глобальной температуры. Это пример положительной обратной связи. Подсчитано, что повышение температуры на 2 °C приведет к дополнительному выбросу в атмосферу 10 Пг углерода в год в результате дыхания почвы. [32] Это больше, чем текущие антропогенные выбросы углерода. Также существует вероятность того, что это повышение температуры приведет к высвобождению углерода, хранящегося в вечно замерзших почвах, которые сейчас тают. Климатические модели предполагают, что эта положительная обратная связь между дыханием почвы и температурой приведет к уменьшению запасов углерода в почве к середине 21 века. [33]
Краткое содержание
[ редактировать ]Дыхание почвы — это ключевой экосистемный процесс, который высвобождает углерод из почвы в форме углекислого газа. Углерод хранится в почве в виде органического вещества и используется растениями, бактериями, грибами и животными. Когда это дыхание происходит под землей, его называют дыханием почвы. Температура, влажность почвы и азот регулируют скорость превращения углерода в органических соединениях почвы в CO 2 . Для измерения дыхания почвы используется множество методов; однако закрытая динамическая камера и использование соотношений стабильных изотопов являются двумя наиболее распространенными методами. Люди изменили уровни CO 2 в атмосфере , характер осадков и нормы внесения удобрений, и все это оказало значительную роль на скорость дыхания почвы. Изменения этих показателей могут изменить глобальные циклы углерода и питательных веществ, а также сыграть значительную роль в изменении климата.
Ссылки
[ редактировать ]- ^ Берг Дж., Тимочко Дж., Страйер Л. (2002). Биохимия. WH Фриман и компания.
- ^ Кляйн Д., Прескотт Л., Харли Дж. (2005). Микробиология. МакГроу-Хилл.
- ^ Шибистова, Ольга; Ллойд, Джон; Евграфова Светлана; Савушкина, Надя; Зражевская Галина; Арнет, Альмут; Кноль, Александр; Колле, Олаф; Шульце, Э.-Детлеф (ноябрь 2002 г.). «Сезонная и пространственная изменчивость темпов выбросов CO 2 в почву для центрально-сибирского леса Pinus sylvestris». Теллус Б. 54 (5): 552–567. дои : 10.1034/j.1600-0889.2002.01348.x .
- ^ Хатч Б., Августин Дж., Мербах В. (2002) Ризоотложение растений – важный источник оборота углерода в почвах. Журнал питания растений и почвоведения. 165, 4, 397–407.
- ^ Перейти обратно: а б Вэнс Э., Чапин III Ф. (2001) Субстратные ограничения микробной активности в таежных лесах. Биология и биохимия почвы. 33, 2, 173–188.
- ^ Харрисон М. (2005) Мирные переговоры и торговые сделки. Ключи к долгосрочной гармонии в бобово-микробных симбиозах. Физиология растений. 137, 4, 1205–1210.
- ^ Чапин III Ф, Мэтсон П., Муни Х. (2002) Принципы экологии наземных экосистем. Спрингер-Верлаг, Нью-Йорк.
- ^ Аткин О., Эдвардс Э., Лавис Б. (2000) Реакция дыхания корней на изменения температуры и ее связь с глобальным потеплением. Новый фитолог. 147, 141–154.
- ^ Микан С., Шимел Дж., Дойл А. (2002) Контроль температуры микробного дыхания в почвах арктической тундры выше и ниже точки замерзания. Биология и биохимия почвы. 34, 1785–1795.
- ^ Перейти обратно: а б Сюй Л., Бальдокки Д., Тан Дж. (2004) Как влага почвы, пульсация дождя и рост меняют реакцию дыхания и температуры экосистемы. Глобальные биогеохимические циклы. 18.
- ^ Ламберс Х., Чапин III Ф., Понс Т. (1998) Физиологическая экология растений. Спрингер-Верлаг, Нью-Йорк.
- ^ Пезешки, СР; ДеЛон, РД (26 июля 2012 г.). «Окисление-восстановление почвы на водно-болотных угодьях и его влияние на функционирование растений» . Биология . 1 (2): 196–221. дои : 10.3390/biology1020196 . ПМЦ 4009779 . ПМИД 24832223 .
- ^ Пейт Дж., Лэйзелл Д. (1990)Энергетика и биологические затраты на ассимиляцию азота. Биохимия растений. 1–42.
- ^ Синсабо Р., Каррейро М., Реперт Д. (2002)Распределение внеклеточной ферментативной активности в зависимости от состава подстилки, отложения N и потери массы. Биогеохимия. 60, 1–24.
- ^ Пумпанен, Юкка; Лонгдоз, Бернард; Л. Куч, Вернер (2010), «Полевые измерения дыхания почвы: принципы и ограничения, возможности и ограничения различных методов» , Динамика углерода в почве , Cambridge University Press, стр. 16–33, doi : 10.1017/cbo9780511711794.003 , hdl : 2268/211761 , ISBN 978-0-511-71179-4 , получено 7 марта 2020 г.
- ^ Уэйсон С., Рэндольф Дж., Хэнсон П., Гриммонд П., Шмид Х. (2006) Сравнение методов дыхания почвы в лиственном лесу средних широт. Биогеохимия. 80, 173–189.
- ^ Филд C, Болл Дж., Берри Дж. (1989) Фотосинтез, принципы и полевые методы. Физиологическая экология растений, полевые методы и приборы. Чепмен и Холл, Нью-Йорк.
- ^ Конен Ф. и Смит К. (2000) Объяснение линейного увеличения концентрации газа в закрытых камерах, используемых для измерения газообмена между почвой и атмосферой. Европейский журнал почвоведения. 51, 111–117.
- ^ Рошетт П., Фланаган Л., Грегорич Э. (1999) Разделение дыхания почвы на растительные и почвенные компоненты с использованием анализа естественного содержания углерода-13. Журнал Американского общества почвоведения. 63, 1207–1213.
- ^ Экблад, Альф; Хёгберг, Питер (1 марта 2000 г.). «Анализ δ13C CO2 позволяет различать микробное дыхание добавленной C4-сахарозы и другое дыхание почвы в C3-экосистеме» . Растение и почва . 219 (1): 197–209. дои : 10.1023/А:1004732430929 . ISSN 1573-5036 . S2CID 11240355 .
- ^ Хёгберг, П.; Экблад, А. (1 сентября 1996 г.). «Субстратно-индуцированное дыхание, измеренное in situ в экосистеме C3-растений с использованием добавок C4-сахарозы» . Биология и биохимия почвы . 28 (9): 1131–1138. дои : 10.1016/0038-0717(96)00124-1 . ISSN 0038-0717 .
- ^ Меникетти, Л.; Экблад, А.; Кеттерер, Т. (2013). «Органические поправки влияют на δ13C-характеристику дыхания почвы и накопления органического углерода в почве в ходе долгосрочного полевого эксперимента в Швеции» . Европейский журнал почвоведения . 64 (5): 621–628. дои : 10.1111/ejss.12077 . ISSN 1365-2389 . S2CID 93540574 .
- ^ Перейти обратно: а б Райк, Ингрид Дж. К.; Экблад, Альф (1 апреля 2020 г.). «Круговорот углерода и азота на лугах, загрязненных свинцом, оценен с использованием стабильных изотопов (δ13C и δ15N), а также микробных, растительных и почвенных параметров» . Растение и почва . 449 (1): 249–266. дои : 10.1007/s11104-020-04467-7 . ISSN 1573-5036 . S2CID 212689936 .
- ^ Плантон, Серж; Дек, Мишель; Шовен, Фабрис; Террей, Лоран (2008). «Ожидаемое воздействие изменения климата на экстремальные климатические явления» . Геофизические отчеты . 340 (9–10): 564–574. дои : 10.1016/j.crte.2008.07.009 .
- ^ Берар, А; Бен Сасси, М; Кайзерманн, А; Рено, П (3 декабря 2015 г.). «Реакция почвенного микробного сообщества на компоненты тепловой волны: засуха и высокая температура» . Климатические исследования . 66 (3): 243–264. дои : 10.3354/cr01343 . ISSN 0936-577X .
- ^ Липсон Д., Уилсон Р., Очел В. (2005) Влияние повышенного содержания CO 2 в атмосфере на биомассу, активность и разнообразие почвенных микробов в экосистеме чапараля. Прикладная и экологическая микробиология. 71, 12, 8573–8580
- ^ Кинг Дж., Хэнсон П., Бернхардт Э., Деангелис П., Норби Р., Прегитцер К. (2004) Многолетний синтез реакций дыхания почвы на повышенный уровень CO 2 в атмосфере на основе четырех лесных экспериментов FACE. Биология глобальных изменений. 10, 1027–1042.
- ^ Оечел В. , Вурлитис Г., Гастингс С. (1995) Изменение потока CO 2 в Арктике за два десятилетия, Последствия изменения климата в Барроу, Аляска. Экологические приложения. 5, 3, 846–855.
- ^ Лютце Дж., Гиффорд Р., Адамс Х. (2000) Качество подстилки и разложение в дернах Danthonia richardsonii в ответ на поставку CO 2 и азота в течение четырех лет роста. Биология глобальных изменений. 6, 13–24.
- ^ Сабина С, Хеманн М, Артаксо П, Баккер Д, Чен С, Филд С, Грубер Н, Ле Кер С, Принн Р, Ричи Дж, Ромеро-Ланкао П, Сатхай Дж, Валентини Р. (2003) Текущий статус и прошлое тенденции углеродного цикла. На пути к стабилизации CO 2 : проблемы, стратегии и последствия. Остров Пресс. Вашингтон, округ Колумбия.
- ^ Райх Дж. и Поттер С. (1995) Глобальные закономерности выбросов углекислого газа из почв. Глобальные биогеохимические циклы. 9, 23–36.
- ^ Фридлингштейн П., Дюфрен Дж., Кокс П. (2003) Насколько положительна обратная связь между изменением климата и глобальным углеродным циклом? Теллус. 55Б, 692–700.
- ^ Кокс П., Беттс Р., Джонс С., Сполл С., Тоттерделл И. (2000) Ускорение глобального потепления из-за обратной связи углеродного цикла в связанной климатической модели. Природа. 408, 184–187.
- Ван Ю, АмундсоР, Трамбор С (1999). «Влияние изменения землепользования на оборот углерода в почвах» (PDF) . Глобальные биогеохимические циклы . 13 (1): 47–57. Бибкод : 1999GBioC..13...47W . дои : 10.1029/1998GB900005 .
- Су Б. (2005) Взаимодействие между циклами углерода, азота и воды в экосистемах в условиях глобальных изменений: результаты полевых экспериментов и экспериментов в мезокосме. Университет Оклахомы, Норман, ОК.
- Фланаган Л. и Веум А. (1974) Взаимосвязь между дыханием, потерей веса, температурой и влажностью в органических остатках в тундре. Почвенные организмы и разложение в тундре. 249–277.