Биолюминесцентные бактерии

Биолюминесцентные бактерии — это светообразующие бактерии , которые преимущественно присутствуют в морской воде, морских отложениях, на поверхности разлагающейся рыбы и в кишечнике морских животных. Бактериальная биолюминесценция встречается не так часто, но также встречается у наземных и пресноводных бактерий. [1] Эти бактерии [ нужны разъяснения ] могут жить свободно (например, Vibrio harveyi ) или находиться в симбиозе с животными, такими как гавайский бобтейл ( Aliivibrio fischeri ) или наземными нематодами ( Photorhabdus luminescens ). Организмы-хозяева обеспечивают эти бактерии. [ нужны разъяснения ] безопасный дом и достаточное питание. Взамен хозяева используют свет, производимый бактериями, для маскировки, привлечения добычи и/или партнера. Биолюминесцентные бактерии развили симбиотические отношения с другими организмами, в которых оба участника получают почти одинаковую выгоду. [2] Другая возможная причина, по которой бактерии используют реакцию люминесценции, — это чувство кворума , способность регулировать экспрессию генов в ответ на плотность бактериальных клеток. [3]
История
[ редактировать ]Записи о биолюминесценции бактерий существуют уже тысячи лет. [4] Они появляются в фольклоре многих регионов, включая Скандинавию и Индийский субконтинент. И Аристотель , и Чарльз Дарвин описали явление свечения океанов. [4] С момента своего открытия менее 30 лет назад фермент люцифераза и ее регуляторный ген lux привели к крупным достижениям в молекулярной биологии благодаря использованию в качестве репортерного гена. [5] Люцифераза была впервые очищена МакЭлроем и Грином в 1955 году. [6] Позже было обнаружено, что люцифераза состоит из двух субъединиц, называемых субъединицами α и β. Гены, кодирующие эти ферменты, luxA и luxB соответственно, были впервые выделены в lux опероне Aliivibrio fisheri. [4] .
Назначение биолюминесценции
[ редактировать ]Широкомасштабные биологические цели биолюминесценции включают, помимо прочего, привлечение партнеров, [7] защита от хищников и предупреждающие сигналы. В случае биолюминесцентных бактерий биолюминесценция в основном служит формой распространения. Была выдвинута гипотеза, что кишечные бактерии (бактерии, которые выживают в кишечнике других организмов) – особенно те, которые распространены в глубинах океана – используют биолюминесценцию как эффективную форму распространения. [8] Пробравшись в пищеварительный тракт рыб и других морских организмов и выделяясь с фекальными гранулами, биолюминесцентные бактерии способны использовать свои биолюминесцентные способности для заманивания других организмов и быстрого проглатывания этих содержащих бактерии фекальных гранул. [9] Таким образом, биолюминесценция бактерий обеспечивает их выживание, устойчивость и распространение, поскольку они могут проникать в другие организмы и заселять их.
Регуляция биолюминесценции
[ редактировать ]Регулирование биолюминесценции у бактерий достигается за счет регуляции окислительного фермента, называемого люциферазой . Важно, что биолюминесцентные бактерии снижают скорость производства люциферазы, когда популяция немногочисленна, в целях экономии энергии. Таким образом, бактериальная биолюминесценция регулируется посредством химической связи, называемой ощущением кворума . [10] По сути, определенные сигнальные молекулы, называемые аутоиндукторами. [11] со специфическими бактериальными рецепторами активируются, когда плотность популяции бактерий достаточно высока. Активация этих рецепторов приводит к скоординированной индукции выработки люциферазы, что в конечном итоге приводит к видимому свечению . [12]
Биохимия биолюминесценции
[ редактировать ]
Химическая реакция, отвечающая за биолюминесценцию, катализируется ферментом люциферазой . В присутствии кислорода люцифераза катализирует окисление органической молекулы, называемой люциферином . [13] Хотя биолюминесценция у самых разных организмов, таких как бактерии, насекомые и динофлагелляты, функционирует таким общим образом (с использованием люциферазы и люциферина), существуют разные типы люциферин-люциферазных систем. В частности, что касается бактериальной биолюминесценции, биохимическая реакция включает окисление алифатического альдегида восстановленным флавинмононуклеотидом . [14] Продукты этой реакции окисления включают окисленный флавинмононуклеотид, цепь жирной кислоты и энергию в виде сине-зеленого видимого света. [15]
Реакция: FMNH 2 + O 2 + RCHO → FMN + RCOOH + H 2 O + свет.
Эволюция биолюминесценции
[ редактировать ]Из всех излучателей света в океане биолюминесцентные бактерии являются наиболее многочисленными и разнообразными. Однако распределение биолюминесцентных бактерий неравномерно, что позволяет предположить эволюционную адаптацию. Виды бактерий наземных родов, таких как Photorhabdus, обладают биолюминесценцией. С другой стороны, морские роды с биолюминесцентными видами, такими как Vibrio и Shewanella oneidensis, имеют разные близкородственные виды, которые не являются излучателями света. [16] Тем не менее, все биолюминесцентные бактерии имеют общую последовательность генов: ферментативное окисление альдегида и восстановленного мононуклеотида флавина люциферазой , которые содержатся в опероне lux. [17] Бактерии из различных экологических ниш содержат эту последовательность генов; следовательно, идентичная последовательность генов, очевидно, предполагает, что биолюминесцентные бактерии являются результатом эволюционных адаптаций.
Использование в качестве лабораторного инструмента
[ редактировать ]Утверждается, что после открытия оперона люкс использование биолюминесцентных бактерий в качестве лабораторного инструмента произвело революцию в области экологической микробиологии. [4] Биолюминесцентные бактерии применяются в биосенсорах для обнаружения загрязняющих веществ, измерения токсичности загрязняющих веществ. [4] [18] [19] и мониторинг генетически модифицированных бактерий, попадающих в окружающую среду. [20] [21] [22] Биосенсоры, созданные путем помещения конструкции гена lux под контроль индуцируемого промотора, могут использоваться для определения концентрации конкретных загрязнителей. [4] Биосенсоры также способны отличать биодоступные загрязнители от инертных и недоступных. [4] Например, Pseudomonas fluorescens была генетически модифицирована так, чтобы она могла разлагать салицилат и нафталин, и используется в качестве биосенсора для оценки биодоступности салицилата и нафталина. [4] Биосенсоры также можно использовать в качестве индикатора клеточной метаболической активности и для обнаружения присутствия патогенов. [4]
Эволюция
[ редактировать ]Химия светопроизводства, лежащая в основе биолюминесценции, варьируется в зависимости от линии биолюминесцентных организмов. [16] На основании этого наблюдения считается, что биолюминесценция развивалась независимо по крайней мере 40 раз. [16] У биолюминесцентных бактерий реклассификация членов группы видов Vibrio fischeri в новый род Aliivibrio привела к повышенному интересу к эволюционному происхождению биолюминесценции. [16] . Среди бактерий распространение биолюминесцентных видов полифилетично. Например, в то время как все виды наземного рода Photorhabdus являются люминесцентными, роды Aliivibrio, Photobacterium, Shewanella и Vibrio содержат как светящиеся, так и несветящиеся виды. [16] Несмотря на то, что биолюминесценция бактерий не имеет общего происхождения, все они имеют общую последовательность генов. Появление высококонсервативного оперона lux у бактерий из самых разных экологических ниш предполагает сильное селективное преимущество, несмотря на высокие энергетические затраты на производство света. Считается, что восстановление ДНК является первоначальным селективным преимуществом производства света у бактерий. [16] Следовательно, оперон lux, возможно, был утерян у бактерий, которые развили более эффективные системы репарации ДНК, но сохранился у тех, у которых видимый свет стал селективным преимуществом. [16] [23] Считается, что эволюция чувства кворума дала дополнительные селективные преимущества в производстве света. Чувство кворума позволяет бактериям экономить энергию, гарантируя, что они не синтезируют светоизлучающие химические вещества, если их концентрация не достаточна для того, чтобы их можно было увидеть. [16]
Группы бактерий, проявляющие биолюминесценцию
[ редактировать ]Все виды бактерий, обладающие биолюминесценцией, принадлежат к семействам Vibrionaceae , Shewanellaceae или Enterobacteriaceae , все из которых относятся к классу Gammaproteobacteria . [24]
| Семья | Род | Разновидность |
|---|---|---|
| Энтеробактерии | Фоторабдус | Фоторабдус асимбиотический |
| Шеванелловые | Шеванелла | Шеванелла Вуди |
| Вибрионовые | Алиивибрио | Алиивибрио фишери |
| Фотобактерия | Фотобактерия аквимарис | |
| Вибрион | Вибрион лазурный | |
| « Кандидатус Фотодесмус ». | « Катоптрон Кандидат Фотодесмус » |
(Список Данлэпа и Хенрика (2013), «Светящиеся бактерии», «Прокариоты»). [24] )
Распределение
[ редактировать ]Биолюминесцентные бактерии наиболее распространены в морской среде во время весеннего цветения, когда наблюдается высокая концентрация питательных веществ. Эти светоизлучающие организмы встречаются в основном в прибрежных водах вблизи устья рек, таких как северная часть Адриатического моря, Триестский залив, северо-западная часть Каспийского моря, побережье Африки и многие другие. [25] Они известны как молочные моря . Биолюминесцентные бактерии также встречаются в пресноводной и наземной среде, но менее широко распространены, чем в морской воде. Они встречаются по всему миру в виде свободноживущих, симбиотических или паразитических форм. [1] и, возможно, как условно-патогенные микроорганизмы . [24] Факторы, влияющие на распространение биолюминесцентных бактерий, включают температуру, соленость, концентрацию питательных веществ, уровень pH и солнечную радиацию. [26] Например, Aliivibrio fischeri хорошо растет в среде с температурой от 5 до 30 °C и pH менее 6,8; тогда как Photobacterium фосфореум процветает в условиях с температурой от 5 до 25 ° C и pH менее 7,0. [27]
Генетическое разнообразие
[ редактировать ]Все биолюминесцентные бактерии имеют общую последовательность генов: оперон lux , характеризующийся организацией гена luxCDABE . [24] LuxAB кодирует люциферазу, а luxCDE кодирует комплекс редуктазы жирных кислот, который отвечает за синтез альдегидов для биолюминесцентной реакции. Несмотря на эту общую организацию генов, среди видов можно наблюдать вариации, такие как наличие других генов lux. На основании сходства в содержании и организации генов оперон lux можно разделить на следующие четыре различных типа: тип Aliivibrio / Shewanella , тип Photobacterium , тип Vibrio / Candidatus Photodesmus и тип Photorhabdus . Хотя эта организация соответствует уровню классификации родов представителей Vibrionaceae ( Aliivibrio , Photobacterium и Vibrio ), ее эволюционная история неизвестна. [24]
За исключением типа оперона Photorhabdus , все варианты оперона lux , кодирующий флавинредуктазу содержат ген luxG . [24] Большинство оперонов типа Aliivibrio / Shewanella содержат дополнительные регуляторные гены luxI/luxR , которые используются для аутоиндукции во время определения кворума . [28] Тип оперона Photobacterum характеризуется наличием ребристых генов, кодирующих рибофлавин, и образует lux-rib оперон . Тип оперона Vibrio / Candidatus Photodesmus отличается как от типов оперонов Aliivibrio / Shewanella, так и от типов оперонов Photobacterium тем, что оперон не имеет непосредственно связанных с ним регуляторных генов. [24]
Механизм
[ редактировать ]Все бактериальные люциферазы представляют собой гетеродимеры массой около 80 кДа, содержащие две субъединицы: α и β. Субъединица α отвечает за излучение света. [4] Гены luxA и luxB кодируют субъединицы α и β соответственно. У большинства биолюминесцентных бактерий гены luxA и luxB фланкированы выше luxC и luxD и ниже luxE . [4]
Биолюминесцентная реакция происходит следующим образом:
FMNH 2 + O 2 + R-CHO -> FMN + H 2 O + R-COOH + Свет (~495 нм)
Молекулярный кислород реагирует с FMNH 2 (восстановленным флавинмононуклеотидом) и длинноцепочечным альдегидом с образованием FMN (флавинмононуклеотида), воды и соответствующей жирной кислоты. В результате этой реакции возникает сине-зеленое световое излучение биолюминесценции, например, производимое Photobacterium фосфореум и Vibro harveyi . [4] Поскольку при излучении света на каждый фотон расходуется шесть молекул АТФ, это энергетически затратный процесс. По этой причине световое излучение у биолюминесцентных бактерий не выражено конститутивно; оно выражается только тогда, когда это физиологически необходимо.
Определение кворума
[ редактировать ]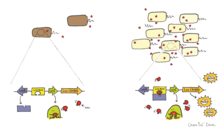
Биолюминесценцию бактерий можно регулировать посредством явления, известного как аутоиндукция или ощущение кворума . [4] Ощущение кворума — это форма межклеточной коммуникации, которая изменяет экспрессию генов в ответ на плотность клеток. Аутоиндуктор представляет собой диффундирующий феромон, конститутивно вырабатываемый биолюминесцентными бактериями и служащий внеклеточной сигнальной молекулой. [4] Когда концентрация аутоиндуктора, секретируемого биолюминесцентными клетками в окружающей среде, достигает порога (выше 10 7 клеток на мл), он индуцирует экспрессию люциферазы и других ферментов, участвующих в биолюминесценции. [4] Бактерии способны оценивать свою плотность, определяя уровень аутоиндукторов в окружающей среде, и регулировать свою биолюминесценцию так, что она проявляется только при достаточно высокой популяции клеток. Достаточно высокая популяция клеток гарантирует, что биолюминесценция, производимая клетками, будет видна в окружающей среде.
Хорошо известным примером ощущения кворума является то, что происходит между Aliivibrio fischeri и его хозяином. Этот процесс регулируется LuxI и LuxR, кодируемыми luxI и luxR соответственно. LuxI представляет собой аутоиндуктор-синтазу, которая производит аутоиндуктор (AI), тогда как LuxR действует как рецептор и фактор транскрипции для оперона lux. [4] Когда LuxR связывает AI, комплекс LuxR-AI активирует транскрипцию оперона lux и индуцирует экспрессию люциферазы . [28] Используя эту систему, A. fischeri показала, что биолюминесценция проявляется только тогда, когда бактерии ассоциированы с хозяином и достигли достаточной плотности клеток. [29]
Другим примером определения кворума биолюминесцентными бактериями являются Vibrio harveyi , которые, как известно, являются свободноживущими. В отличие от Aliivibrio fischeri , V. harveyi не обладает регуляторными генами luxI/luxR и, следовательно, имеет другой механизм регуляции кворума. Вместо этого они используют систему, известную как трехканальная система определения кворума. [30] Вибрионы используют небольшие некодирующие РНК, называемые Qrr РНК , для регуляции чувства кворума и контроля трансляции энергозатратных молекул.
Роль
[ редактировать ]возможности использования биолюминесценции Широко изучены , ее биологическое и экологическое значение для животных, в том числе для организмов-хозяев, для бактериального симбиоза. Биологическая роль и история эволюции именно биолюминесцентных бактерий до сих пор остаются весьма загадочными и неясными. [4] [31] Тем не менее, постоянно проводятся новые исследования, чтобы определить влияние, которое бактериальная биолюминесценция может оказать на нашу постоянно меняющуюся окружающую среду и общество. Помимо множества научных и медицинских применений, ученые также недавно начали объединяться с художниками и дизайнерами, чтобы исследовать новые способы включения биолюминесцентных бактерий, а также биолюминесцентных растений в городские источники света, чтобы снизить потребность в электроэнергии. [32] Они также начали использовать биолюминесцентные бактерии как форму искусства и городского дизайна, чтобы доставить удовольствие человеческому обществу. [33] [34] [35]
Одно из объяснений роли бактериальной биолюминесценции лежит в биохимическом аспекте. Несколько исследований показали биохимическую роль пути люминесценции. Он может функционировать как альтернативный путь потока электронов при низкой концентрации кислорода, что может быть выгодно, когда ферментируемый субстрат недоступен. [1] В этом процессе световое излучение является побочным продуктом обмена веществ.
Данные также свидетельствуют о том, что бактериальная люцифераза способствует устойчивости к окислительному стрессу. В лабораторной культуре luxA и luxB мутанты Vibrio harveyi , у которых отсутствовала люциферазная активность, показали нарушение роста в условиях высокого окислительного стресса по сравнению с диким типом. Мутанты luxD , которые имели незатронутую люциферазу, но не могли производить люминесценцию, показали незначительные различия или вообще не имели различий. Это предполагает, что люцифераза опосредует детоксикацию активного кислорода. [36]
Было также предложено, чтобы бактериальная биолюминесценция была источником внутреннего света при фотореактивации, процессе восстановления ДНК, осуществляемом фотолиазой. [37] Эксперименты показали, что нелюминесцентные мутанты V. harveyi более чувствительны к УФ-облучению, что позволяет предположить существование биолюминесцентно-опосредованной системы репарации ДНК. [23]
Другая гипотеза, названная «гипотезой приманки», заключается в том, что бактериальная биолюминесценция привлекает хищников, которые будут способствовать их распространению. [37] Они либо напрямую поедаются рыбой, либо косвенно поедаются зоопланктоном, который в конечном итоге будет потребляться более высокими трофическими уровнями. В конечном итоге это может позволить бактериям проникнуть в кишечник рыбы, богатую питательными веществами среду, где бактерии могут делиться, выделяться и продолжать свой цикл. Эксперименты с использованием люминесцентных Photobacterium leiognathi и нелюминесцентных мутантов показали, что свечение привлекает зоопланктон и рыб, что подтверждает эту гипотезу. [37]
Симбиоз с другими организмами
[ редактировать ]симбиотические отношения между сколопами гавайского кальмара Euprymna и морской грамотрицательной бактерией Aliivibrio fischeri Хорошо изучены . Эти два организма демонстрируют мутуалистические отношения, при которых биолюминесценция, производимая A. fischeri, помогает привлечь жертву к кальмару-хозяину, который обеспечивает богатые питательными веществами ткани и защищенную среду для A. fischeri . [38] Биолюминесценция, обеспечиваемая A. fischeri, также помогает защитить скопы E. кальмара , обеспечивая маскировку во время его ночной кормовой деятельности. [39] После бактериальной колонизации специализированные органы кальмара претерпевают изменения в развитии, и между ними устанавливаются взаимоотношения. Кальмар каждое утро уничтожает 90% популяции бактерий, поскольку ему больше не нужно производить биолюминесценцию при дневном свете. [4] Это изгнание приносит пользу бактериям, способствуя их распространению. Одно изгнание одного кальмара-куцехвоста производит достаточно бактериальных симбионтов, чтобы заполнить 10 000 м2. 3 морской воды в концентрации, сравнимой с той, что содержится в прибрежных водах. [39] Таким образом, по крайней мере в некоторых местообитаниях симбиотические отношения между A. fischeri и E. scolopes играют ключевую роль в определении численности и распространения E. scolopes . Численность A. fischeri выше в окрестностях популяции E. scolopes и заметно снижается по мере удаления от места обитания хозяина. [39]
Биолюминесцентные виды фотобактерий также вступают во взаимовыгодные связи с рыбами и кальмарами. [40] Плотные популяции P. kishitanii, P. leiogathi и . P. mandapamensis могут обитать в легких органах морских рыб и кальмаров и обеспечиваются питательными веществами и кислородом для размножения [40] в обмен на обеспечение биолюминесценции своим хозяевам, что может помочь в передаче сигналов в зависимости от пола, избегании хищников, обнаружении или привлечении добычи и обучении. [ нужна ссылка ] Интересно, что Мейер-Рохов сообщил в 1976 году, что если рыба не может добыть пищу и голодает, свет ее биолюминесцентного симбионта становится все более тусклым, пока излучение света не прекратится совсем. [41]
| В таблице ниже на изображениях справа синим цветом отмечены места расположения светового органа разных семейств симбиотически светящихся рыб и кальмаров. [42] E указывает на внешний выброс биолюминесцентных бактерий непосредственно в морскую воду. I указывает на внутреннее изгнание биолюминесцентных бактерий в пищеварительный тракт. (E) или (I) указывают на предполагаемую локализацию изгнания. [43] |
 |
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Нилсон, К.Х.; Гастингс, JW (1 декабря 1979 г.). «Бактериальная биолюминесценция: контроль и экологическое значение» . Микробиологические обзоры . 43 (4): 496–518. дои : 10.1128/mmbr.43.4.496-518.1979 . ISSN 0146-0749 . ПМК 281490 . ПМИД 396467 .
- ^ Макфолл-Нгай, Маргарет; Хит-Хекман, Элизабет AC; Джилетт, Амани А.; Пейер, Сюзанна М.; Харви, Элизабет А. (2012). «Тайные языки совместно развившихся симбиозов: выводы из симбиоза сколопов Euprymna и Vibrio fischeri» . Семинары по иммунологии . 24 (1): 3–8. дои : 10.1016/j.smim.2011.11.006 . ПМК 3288948 . ПМИД 22154556 .
- ^ Уотерс, Кристофер М.; Басслер, Бонни Л. (07 октября 2005 г.). «ЗОНДИРОВАНИЕ КВОРУМА: Межклеточная связь у бактерий». Ежегодный обзор клеточной биологии и биологии развития . 21 (1): 319–346. doi : 10.1146/annurev.cellbio.21.012704.131001 . ПМИД 16212498 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р Нуньес-Хальдорсон, Ваня да Силва; Дюран, Норма Летисия (1 июня 2003 г.). «Биолюминесцентные бактерии: гены lux как биосенсоры окружающей среды» . Бразильский журнал микробиологии . 34 (2): 91–96. дои : 10.1590/S1517-83822003000200001 . ISSN 1517-8382 .
- ^ «Репортеры люциферазы» . www.thermofisher.com . Проверено 24 марта 2017 г.
- ^ МакЭлрой, штат Вашингтон; Грин (1954). «Ферментативные свойства бактериальной люциферазы». Архив биохимии и биофизики . 56 (1): 240–255. дои : 10.1016/0003-9861(55)90353-2 . ПМИД 14377570 .
- ^ Херринг, Питер Дж. (1 августа 2007 г.). «ОБЗОР. Секс при включенном свете? Обзор биолюминесцентного полового диморфизма в море» . Журнал Морской биологической ассоциации Соединенного Королевства . 87 (4): 829–842. дои : 10.1017/S0025315407056433 . ISSN 1469-7769 . S2CID 86616254 .
- ^ Руби, Е.Г.; Морен, Дж. Г. (1 сентября 1979 г.). «Светящиеся кишечные бактерии морских рыб: исследование их распространения, плотности и дисперсии» . Прикладная и экологическая микробиология . 38 (3): 406–411. Бибкод : 1979ApEnM..38..406R . дои : 10.1128/АЕМ.38.3.406-411.1979 . ISSN 0099-2240 . ПМК 243508 . ПМИД 16345429 .
- ^ Руби, Е.Г.; Морен, Дж. Г. (1 сентября 1979 г.). «Светящиеся кишечные бактерии морских рыб: исследование их распределения, плотности и дисперсии» . Прикладная и экологическая микробиология . 38 (3): 406–411. Бибкод : 1979ApEnM..38..406R . дои : 10.1128/АЕМ.38.3.406-411.1979 . ISSN 0099-2240 . ПМК 243508 . ПМИД 16345429 .
- ^ Уилсон, Тереза; Гастингс, Дж. Вудленд (28 ноября 2003 г.). «БИОЛЮМИНЕСЦЕНЦИЯ». Ежегодный обзор клеточной биологии и биологии развития . 14 (1): 197–230. doi : 10.1146/annurev.cellbio.14.1.197 . ПМИД 9891783 .
- ^ Эберхард, А.; Берлингейм, Алабама; Эберхард, К.; Кеньон, GL; Нилсон, К.Х.; Оппенгеймер, Нью-Джерси (1 апреля 1981 г.). «Структурная идентификация аутоиндуктора люциферазы Photobacterium fischeri». Биохимия . 20 (9): 2444–2449. дои : 10.1021/bi00512a013 . ISSN 0006-2960 . ПМИД 7236614 .
- ^ Перес-Веласкес, Юдит; Гёлгели, Мельтем; Гарсиа-Контрерас, Родольфо (25 августа 2016 г.). «Математическое моделирование определения бактериального кворума: обзор» (PDF) . Бюллетень математической биологии . 78 (8): 1585–1639. дои : 10.1007/s11538-016-0160-6 . hdl : 11693/36891 . ISSN 0092-8240 . ПМИД 27561265 . S2CID 3940300 .
- ^ Каськова Зинаида М.; Царькова Александра С.; Ямпольский, Илья В. (24 октября 2016 г.). «1001 свет: люциферины, люциферазы, их механизмы действия и применение в химическом анализе, биологии и медицине». хим. Соц. Преподобный . 45 (21): 6048–6077. дои : 10.1039/c6cs00296j . ISSN 1460-4744 . ПМИД 27711774 .
- ^ Уайлс, Сьюзи ; Фергюсон, Кэтрин; Стефаниду, Марта; Янг, Дуглас Б.; Робертсон, Брайан Д. (1 июля 2005 г.). «Альтернативная люцифераза для мониторинга бактериальных клеток в неблагоприятных условиях» . Прикладная и экологическая микробиология . 71 (7): 3427–3432. Бибкод : 2005ApEnM..71.3427W . дои : 10.1128/АЕМ.71.7.3427-3432.2005 . ISSN 0099-2240 . ПМК 1169068 . ПМИД 16000745 .
- ^ Фишер, Эндрю Дж.; Томпсон, Томас Б.; Тоден, Джеймс Б.; Болдуин, Томас О.; Реймент, Иван (6 сентября 1996 г.). «Кристаллическая структура бактериальной люциферазы с разрешением 1,5 Å в условиях низкого содержания соли» . Журнал биологической химии . 271 (36): 21956–21968. дои : 10.1074/jbc.271.36.21956 . ISSN 0021-9258 . ПМИД 8703001 .
- ^ Jump up to: а б с д и ж г час Виддер, Э.А. (07 мая 2010 г.). «Биолюминесценция в океане: происхождение биологического, химического и экологического разнообразия». Наука . 328 (5979): 704–708. Бибкод : 2010Sci...328..704W . дои : 10.1126/science.1174269 . ISSN 0036-8075 . ПМИД 20448176 . S2CID 2375135 .
- ^ О'Кейн, Деннис Дж.; Прашер, Дуглас К. (1 февраля 1992 г.). «Эволюционное происхождение бактериальной биолюминесценции» . Молекулярная микробиология . 6 (4): 443–449. дои : 10.1111/j.1365-2958.1992.tb01488.x . ISSN 1365-2958 . ПМИД 1560772 . S2CID 42286135 .
- ^ Птицын Л.Р.; Хорнек, Г.; Комова О.; Козубек С.; Красавин Е.А.; Бонев, М.; Реттберг, П. (1 ноября 1997 г.). «Биосенсор для скрининга генотоксинов окружающей среды на основе анализа SOS lux в рекомбинантных клетках Escherichia coli» . Прикладная и экологическая микробиология . 63 (11): 4377–4384. Бибкод : 1997ApEnM..63.4377P . дои : 10.1128/АЕМ.63.11.4377-4384.1997 . ISSN 0099-2240 . ПМК 168758 . ПМИД 9361425 .
- ^ Виддер, Эдит А.; Фолс, Бет (март 2014 г.). «Обзор биолюминесценции для инженеров и ученых в области биофотоники». Журнал IEEE по избранным темам квантовой электроники . 20 (2): 232–241. Бибкод : 2014IJSTQ..20..232W . дои : 10.1109/JSTQE.2013.2284434 . ISSN 1077-260X . S2CID 33866877 .
- ^ Махаффи, WF; Бауске, Э.М.; ван Вуурде, JW; ван дер Вольф, Дж. М.; ван ден Бринк, М.; Клоппер, JW (1 апреля 1997 г.). «Сравнительный анализ устойчивости к антибиотикам, иммунофлуоресцентного окрашивания колоний и трансгенного маркера (биолюминесценции) для мониторинга судьбы ризобактерий в окружающей среде» . Прикладная и экологическая микробиология . 63 (4): 1617–1622. Бибкод : 1997ApEnM..63.1617M . дои : 10.1128/АЕМ.63.4.1617-1622.1997 . ISSN 0099-2240 . ПМК 168454 . ПМИД 9097457 .
- ^ Шоу, Джей-Джей; Дейн, Ф.; Гейгер, Д.; Клоппер, JW (1 января 1992 г.). «Использование биолюминесценции для обнаружения генно-инженерных микроорганизмов, попавших в окружающую среду» . Прикладная и экологическая микробиология . 58 (1): 267–273. Бибкод : 1992ApEnM..58..267S . дои : 10.1128/АЕМ.58.1.267-273.1992 . ISSN 0099-2240 . ПМК 195202 . ПМИД 1311542 .
- ^ Флемминг, Калифорния; Ли, Х.; Треворс, Джей Ти (1 сентября 1994 г.). «Биолюминесцентный метод наиболее вероятного числа для подсчета отмеченных люксом Pseudomonas aeruginosa UG2Lr в почве» . Прикладная и экологическая микробиология . 60 (9): 3458–3461. Бибкод : 1994ApEnM..60.3458F . дои : 10.1128/АЕМ.60.9.3458-3461.1994 . ISSN 0099-2240 . ЧВК 201832 . ПМИД 16349396 .
- ^ Jump up to: а б Чиз, А.; Врубель, Б.; Вегжин, Г. (1 февраля 2000 г.). «Биолюминесценция Vibrio harveyi играет роль в стимуляции восстановления ДНК». Микробиология . 146 (2): 283–288. дои : 10.1099/00221287-146-2-283 . ISSN 1350-0872 . ПМИД 10708366 .
- ^ Jump up to: а б с д и ж г Делонг, Эдвард Ф.; Штакебрандт, Эрко; Лори, Стивен; Томпсон, Фабиано (2013). Прокариоты — Спрингер . дои : 10.1007/978-3-642-31331-8 . ISBN 978-3-642-31330-1 . S2CID 5215416 .
- ^ Стейплс, Роберт Ф.; Штаты., США (1966). Подробности - Распространение и характеристики поверхностной биолюминесценции в океанах / Роберт Ф. Стейплс. - Библиотека наследия биоразнообразия . дои : 10.5962/bhl.title.46935 .
- ^ Сото, В.; Гутьеррес, Дж.; Ремменга, доктор медицины; Нисигути, МК (1 января 2009 г.). «Влияние солености и температуры на физиологические реакции Vibrio fischeri из разнообразных экологических ниш» . Микробная экология . 57 (1): 140–150. дои : 10.1007/s00248-008-9412-9 . ISSN 0095-3628 . ПМК 2703662 . ПМИД 18587609 .
- ^ Уотерс, Пол; Ллойд, Дэвид (1985). «Соль, pH и температурные зависимости роста и биолюминесценции трех видов светящихся бактерий, проанализированные на градиентных пластинах» . Журнал общей микробиологии . 131 (11): 65–9. дои : 10.1099/00221287-131-11-2865 .
- ^ Jump up to: а б Басслер, Б.Л. (1 декабря 1999 г.). «Как бактерии разговаривают друг с другом: регуляция экспрессии генов путем определения кворума». Современное мнение в микробиологии . 2 (6): 582–587. дои : 10.1016/s1369-5274(99)00025-9 . ISSN 1369-5274 . ПМИД 10607620 .
- ^ Фукуа, туалет; Винанс, Южная Каролина; Гринберг, EP (1 января 1994 г.). «Ощущение кворума у бактерий: семейство регуляторов транскрипции LuxR-LuxI, чувствительных к клеточной плотности» . Журнал бактериологии . 176 (2): 269–275. дои : 10.1128/jb.176.2.269-275.1994 . ISSN 0021-9193 . ПМК 205046 . ПМИД 8288518 .
- ^ Дефойрдт, Том; Бун, Нико; Беззаботный, Патрик; Стрете, Вилли; Боссье, Питер (1 января 2008 г.). «Ощущение кворума и подавление кворума у Vibrio harveyi: уроки, извлеченные из работы in vivo» . Журнал ISME . 2 (1): 19–26. дои : 10.1038/ismej.2007.92 . ISSN 1751-7362 . ПМИД 18180744 .
- ^ Ваннье, Томас; Хингамп, Паскаль; Террел, Флориана; Танет, Лиза; Лескот, Магали; Тимсит, Юрий (01.06.2020). «Разнообразие и эволюция бактериальных генов биолюминесценции в мировом океане» . НАР Геномика и биоинформатика . 2 (2): lqaa018. дои : 10.1093/nargab/lqaa018 . ПМЦ 7671414 . PMID 33575578 .
- ^ Ардавани, Олимпия; Зерефос, Стелиос; Дулос, Ламброс Т. (01 января 2020 г.). «Редизайн наружного освещения как части городского ландшафта: роль трансгенных биолюминесцентных растений в средиземноморских городских и пригородных условиях освещения» . Журнал чистого производства . 242 : 118477. doi : 10.1016/j.jclepro.2019.118477 . ISSN 0959-6526 .
- ^ Латц, Майкл И. (2019). «Создание куба бесконечности, выставки биолюминесцентного искусства» . Бюллетень лимнологии и океанографии . 28 (4): 130–134. дои : 10.1002/lob.10345 . ISSN 1539-6088 .
- ^ ДИБАС, ШЕРИЛ ЛИН (2019). «Новая волна НАУКИ И ИСКУССТВА ВСТРЕЧАЕТСЯ В МОРЕ». Океанография . 32 (1): 10–12. ISSN 1042-8275 . JSTOR 26604937 .
- ^ «Больше, чем человеческая медиа-архитектура» (PDF) . Материалы 4-й биеннале медиаархитектуры . дои : 10.1145/3284389.3284495 . S2CID 69160272 .
- ^ Шпилевска, Ханна; Чиз, Агата; Вегжин, Гжегож (1 ноября 2003 г.). «Экспериментальные доказательства физиологической роли бактериальной люциферазы в защите клеток от окислительного стресса». Современная микробиология . 47 (5): 379–382. дои : 10.1007/s00284-002-4024-y . ISSN 0343-8651 . ПМИД 14669913 . S2CID 20132449 .
- ^ Jump up to: а б с Зарубина, Маргарита; Белкин, Шимшон; Ионеску, Майкл; Генин, Аматция (17 января 2012 г.). «Бактериальная биолюминесценция как приманка для морского зоопланктона и рыб» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (3): 853–857. Бибкод : 2012PNAS..109..853Z . дои : 10.1073/pnas.1116683109 . ISSN 1091-6490 . ПМК 3271926 . ПМИД 22203999 .
- ^ Нотон, Линн М.; Мандель, Марк Дж. (01 марта 2012 г.). «Колонизация сколопов Euprymna кальмаров Vibrio fischeri » . Журнал визуализированных экспериментов (61): e3758. дои : 10.3791/3758 . ISSN 1940-087X . ПМЦ 3399469 . ПМИД 22414870 .
- ^ Jump up to: а б с Руби, Е.Г.; Ли, К.Х. (1 марта 1998 г.). «Ассоциация органов света Vibrio fischeri-Euprymna: современные экологические парадигмы» . Прикладная и экологическая микробиология . 64 (3): 805–812. Бибкод : 1998ApEnM..64..805R . дои : 10.1128/АЕМ.64.3.805-812.1998 . ISSN 0099-2240 . ПМЦ 106330 . ПМИД 16349524 .
- ^ Jump up to: а б Урбанчик, Хенрик; Аст, Дженнифер С.; Данлэп, Пол В. (01 марта 2011 г.). «Филогения, геномика и симбиоз фотобактерий» . Обзоры микробиологии FEMS . 35 (2): 324–342. дои : 10.1111/j.1574-6976.2010.00250.x . ISSN 0168-6445 . ПМИД 20883503 .
- ^ Мейер-Рохов, Виктор Бенно (1976). «Потеря биолюминесценции в катоптроне Anomalops из-за голодания». Эксперименты . 32 : 1175–1176.
- ^ Нилсон, К.Х. и Гастингс, Дж.В. (1979) «Бактериальная биолюминесценция: ее контроль и экологическое значение», Microbiol. Откр. , 43 : 496–518. дои : 10.1128/MMBR.43.4.496-518.1979 .
- ^ Танет, Лиза; Мартини, Северин; Казалот, Лори; Тамбурини, Кристиан (2020). «Обзоры и синтезы: Бактериальная биолюминесценция – экология и влияние на биологический углеродный насос» . Биогеонауки . 17 (14): 3757–3778. Бибкод : 2020BGeo...17.3757T . дои : 10.5194/bg-17-3757-2020 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .