Подвижность бактерий
| Часть серии о |
| Движение микробов и микроботов |
|---|
 |
| Микропловцы |
| Молекулярные моторы |
Подвижность бактерий – это способность бактерий передвигаться независимо, используя метаболическую энергию. Большинство механизмов подвижности, которые развились у бактерий, также развивались параллельно и у архей . Большинство палочковидных бактерий могут передвигаться за счет собственной силы, что позволяет колонизировать новые среды обитания и открывать новые ресурсы для выживания. Передвижение бактерий зависит не только от характеристик среды, но и от использования различных придатков для движения. Движения роения и плавания приводятся в движение вращающимися жгутиками . В то время как роение — это многоклеточное двумерное движение по поверхности, требующее присутствия поверхностно-активных веществ , плавание — это перемещение отдельных клеток в жидкой среде.
Другие типы движений, происходящих на твердых поверхностях, включают подергивания, скольжение и скольжение, которые не зависят от жгутиков. Подергивание зависит от вытягивания, прикрепления к поверхности и втягивания пилей IV типа , которые тянут клетку вперед подобно действию крюка, обеспечивая энергию для перемещения клетки вперед. При скольжении используются различные двигательные комплексы, например фокальной адгезии комплексы Myxococcus . В отличие от подергивающихся и скользящих движений, которые являются активными движениями, в которых движущая сила создается отдельной клеткой, скольжение является пассивным движением. Он основан на движущей силе, генерируемой клеточным сообществом за счет расширяющих сил, вызванных ростом клеток внутри колонии в присутствии поверхностно-активных веществ, которые уменьшают трение между клетками и поверхностью. Общее движение бактерии может быть результатом чередования фаз падения и плавания. В результате траектория движения бактерии, плавающей в однородной среде, будет образовывать случайное блуждание с относительно прямым плаванием, прерываемым случайными падениями, которые меняют ориентацию бактерии.
Бактерии также могут демонстрировать такси , то есть способность двигаться к раздражителям в их среде или от них. При хемотаксисе общее движение бактерий реагирует на присутствие химических градиентов. При фототаксисе бактерии могут двигаться к свету или от него. Это может быть особенно полезно для цианобактерий , которые используют свет для фотосинтеза . Точно так же магнитотактические бактерии выравнивают свое движение с магнитным полем Земли . У некоторых бактерий есть реакции бегства , позволяющие им уклоняться от раздражителей, которые могут причинить вред или убить. Это фундаментально отличается от навигации или исследования, поскольку время реагирования должно быть быстрым. Реакции ускользания достигаются за счет явлений, подобных потенциалу действия , и наблюдались в биопленках, а также в отдельных клетках, таких как кабельные бактерии .
В настоящее время существует интерес к разработке биогибридных микропловцов , микроскопических пловцов, частично биологических, частично созданных человеком, таких как плавающие бактерии, модифицированные для перевозки грузов.
Фон
[ редактировать ]В 1828 году британский биолог Роберт Браун обнаружил непрерывное покачивающееся движение пыльцы в воде и описал свое открытие в своей статье «Краткий отчет о микроскопических наблюдениях…». [1] что привело к расширенной научной дискуссии о происхождении этого движения. Эта загадка была решена только в 1905 году, когда Альберт Эйнштейн опубликовал свое знаменитое эссе « О движении частиц, взвешенных в покоящихся жидкостях», требуемое молекулярно-кинетической теорией тепла . [2] Эйнштейн не только обнаружил диффузию взвешенных частиц в покоящихся жидкостях, но и предположил, что эти результаты можно использовать для определения размера частиц — в каком-то смысле он был первым в мире микрореологом . [3]
С тех пор, как Ньютон разработал свои уравнения движения, загадка движения на микромасштабе часто всплывала в истории науки, что хорошо продемонстрировано в нескольких статьях, которые следует кратко обсудить. Во-первых, основная концепция, популяризированная Осборном Рейнольдсом , заключается в том, что относительная важность инерции и вязкости для движения жидкости зависит от определенных деталей рассматриваемой системы. [3] Число Рейнольдса Re , названное в его честь, количественно определяет это сравнение как безразмерное соотношение характерных инерционных и вязких сил:


«Быстро или медленно, он точно повторяет свою траекторию и возвращается туда, откуда начал». [4]

Здесь ρ представляет плотность жидкости; u — характерная скорость системы (например, скорость плывущей частицы); l — характерный масштаб длины (например, размер пловца); μ — вязкость жидкости. Принимая в качестве суспендирующей жидкости воду и используя экспериментально наблюдаемые значения u , можно определить, что инерция важна для макроскопических пловцов, таких как рыбы ( Re = 100), в то время как вязкость доминирует в движении микромасштабных пловцов, таких как бактерии ( Re = 10) . −4 ). [3]
Огромное значение вязкости для плавания в микрометровом масштабе имеет глубокие последствия для стратегии плавания. Об этом замечательно говорил Э. М. Перселл , приглашавший читателя в мир микроорганизмов и теоретически изучавший условия их движения. [4] Во-первых, стратегии движения крупных пловцов часто включают в себя передачу импульса окружающей жидкости в периодических дискретных событиях , таких как образование вихрей между этими событиями , и движение по инерции . Это не может быть эффективно для микронных пловцов, таких как бактерии: из-за большого вязкостного демпфирования время инерционного движения объекта микронного размера составляет порядка 1 мкс. Расстояние движения микроорганизма, движущегося с типичной скоростью, составляет около 0,1 ангстрема (Å). Перселл пришел к выводу, что только силы, действующие в настоящий момент на микромасштабное тело, способствуют его движению, поэтому необходим метод постоянного преобразования энергии. [4] [3]
Микроорганизмы оптимизировали свой метаболизм для непрерывного производства энергии, в то время как чисто искусственные микроплаватели (микророботы) должны получать энергию из окружающей среды, поскольку их бортовая емкость очень ограничена. Как дальнейшее следствие непрерывного рассеивания энергии, биологические и искусственные микропловцы не подчиняются законам равновесной статистической физики и должны описываться неравновесной динамикой. [3] С математической точки зрения Перселл исследовал последствия низкого числа Рейнольдса, взяв уравнение Навье-Стокса и исключив инерционные члены:

где - скорость жидкости и это градиент давления . Как заметил Перселл, полученное уравнение — уравнение Стокса — не содержит явной зависимости от времени. [4] Это имеет некоторые важные последствия для того, как подвешенное тело (например, бактерия) может плавать посредством периодических механических движений или деформаций (например, жгутика ) . Во-первых, скорость движения практически не имеет значения для движения микропловец и окружающей жидкости: изменение скорости движения изменит масштаб скоростей жидкости и микропловец, но не изменит структуру жидкости. поток. Во-вторых, изменение направления механического движения просто изменит все скорости в системе. Эти свойства уравнения Стокса сильно ограничивают диапазон возможных стратегий плавания. [4] [3]


В качестве конкретной иллюстрации рассмотрим математический гребешок , состоящий из двух жестких частей, соединенных шарниром. Может ли «гребешок» плавать, периодически открывая и закрывая шарнир? Нет: независимо от того, как цикл открытия и закрытия зависит от времени, гребешок всегда вернется в исходную точку в конце цикла. Отсюда возникла поразительная цитата: «Быстро или медленно, он точно повторяет свою траекторию и возвращается туда, откуда начал». [4] В свете этой теоремы о гребешках Перселл разработал подходы к тому, как можно создать искусственное движение на микромасштабе. [3] Эта статья продолжает вдохновлять постоянные научные дискуссии; например, недавняя работа группы Фишера из Института интеллектуальных систем Макса Планка экспериментально подтвердила, что принцип гребешка справедлив только для ньютоновских жидкостей . [5] [3]

Подвижные системы развивались в естественном мире с течением времени и в масштабах длины, охватывающих несколько порядков, и развивались анатомически и физиологически для достижения оптимальных стратегий самодвижения и преодоления последствий сил высокой вязкости и броуновского движения , как показано на диаграмме. справа. [6] [3]
Некоторые из наименьших известных подвижных систем представляют собой моторные белки , т. е. белки и белковые комплексы, присутствующие в клетках и выполняющие разнообразные физиологические функции путем преобразования химической энергии в механическую . Эти моторные белки классифицируются как миозины , кинезины или динеины . Миозиновые моторы отвечают за мышечные сокращения и транспортировку грузов, используя актиновые нити в качестве дорожек. динеиновые моторы С другой стороны, и кинезиновые моторы используют микротрубочки для транспортировки везикул через клетку. [7] [8] Механизм, который эти белковые моторы используют для преобразования химической энергии в движение, зависит от гидролиза АТФ , который приводит к модификации конформации глобулярного моторного домена, что приводит к направленному движению. [9] [10] [3]
Бактерии можно условно разделить на две принципиально разные группы: грамположительные и грамотрицательные бактерии , различающиеся архитектурой клеточной оболочки. В каждом случае клеточная оболочка представляет собой сложную многослойную структуру, защищающую клетку от окружающей среды. У грамположительных бактерий цитоплазматическая мембрана окружена только толстой клеточной стенкой из пептидогликана . Напротив, оболочка грамотрицательных бактерий более сложна и состоит (изнутри наружу) из цитоплазматической мембраны, тонкого слоя пептидогликана и дополнительной внешней мембраны, также называемой липополисахаридным слоем. Другие структуры поверхности бактериальных клеток варьируются от дезорганизованных слоев слизи до высокоструктурированных капсул . Они состоят из секретируемых слизистых или липких полисахаридов или белков, которые обеспечивают защиту клеток и находятся в прямом контакте с окружающей средой. У них есть и другие функции, в том числе крепление к твердым поверхностям. Дополнительно на поверхности могут присутствовать белковые придатки: бахромки и пили могут иметь разную длину и диаметр, а их функции включают адгезию и подергивающуюся подвижность . [11] [12] [3]
В частности, для микроорганизмов, живущих в водной среде, передвижение относится к плаванию, и, следовательно, мир полон различных классов плавающих микроорганизмов, таких как бактерии, сперматозоиды , простейшие и водоросли . Бактерии перемещаются благодаря вращению волосообразных нитей, называемых жгутиками , которые прикреплены к белковому моторному комплексу на клеточной стенке бактерий. [3]
Механизмы движения
[ редактировать ]
У бактерий есть два разных основных механизма, которые они используют для движения. Жгутик используется для плавания и роения, а пилус (или бахромка) — для подергивания.
Жгутик
[ редактировать ]Жгутик . (множественное число, жгутик; группа жгутиков называется пучком) представляет собой спиральный, тонкий и длинный придаток, прикрепленный к поверхности клетки одним из своих концов, совершающий вращательное движение для толкания или вытягивания клетки [14] [3] Во время вращения бактериального жгутикового двигателя, расположенного в мембране, жгутики вращаются со скоростью от 200 до 2000 об/мин, в зависимости от вида бактерий. Крючковая субструктура бактериального жгутика действует как универсальный сустав, соединяющий двигатель с жгутиковой нитью. [13]


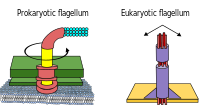
Прокариоты, как бактерии, так и археи, в основном используют жгутики для передвижения.
- Бактериальные жгутики представляют собой спиральные нити, каждая из которых имеет вращающийся двигатель , который может вращаться по или против часовой стрелки. в основании [16] [17] [18] Они обеспечивают два из нескольких видов подвижности бактерий. [19] [20]
- Архейные жгутики называются архаеллами и функционируют почти так же, как бактериальные жгутики. По строению архелла внешне похожа на жгутик бактерий, но отличается во многих деталях и считается негомологичным . [21] [15]
Некоторые эукариотические клетки также используют жгутики — их можно найти у некоторых простейших и растений, а также в клетках животных. Эукариотические жгутики представляют собой сложные клеточные выступы, которые совершают движения взад и вперед, а не круговые движения. Прокариотические жгутики используют вращающийся двигатель, а эукариотические жгутики используют сложную систему скользящих нитей. Эукариотические жгутики управляются АТФ , тогда как прокариотические жгутики могут управляться АТФ (археи) или протонами (бактерии). [22]
В зависимости от количества и расположения жгутиков на поверхности клетки обнаруживаются различные типы жгутиков, например, только у полюсов клетки или рассредоточенные по поверхности клетки. [23] При полярном жгутике жгутики присутствуют на одном или обоих концах клетки: если к одному полюсу прикреплен один жгутик, клетка называется монотрихической; если пучок жгутиков расположен на одном полюсе, клетки лофотриховые; когда жгутики присутствуют на обоих концах, клетка амфитрихна. При перитрихозном жгутиковании жгутики распределяются в разных местах по поверхности клетки. Тем не менее, в рамках этой классификации можно обнаружить вариации, такие как латеральное и субполярное (вместо полярного) монотриховое и лофотриховое жгутикование. [24] [3]
Модель роторного двигателя, используемая бактериями, использует протоны электрохимического градиента для перемещения своих жгутиков. Крутящий момент в жгутиках бактерий создается частицами, которые проводят протоны вокруг основания жгутика. Направление вращения жгутиков у бактерий обусловлено заполненностью протонных каналов по периметру жгутикового мотора. [25]
Жгутик бактерий представляет собой белок-наномашину, преобразующую электрохимическую энергию в виде градиента ионов H+ или Na+ в механическую работу. [26] [27] [28] Жгутик состоит из трёх частей: базального тела, крючка и нити. Базальное тельце представляет собой обратимый двигатель, охватывающий оболочку бактериальной клетки. Он состоит из центрального стержня и нескольких колец: у грамотрицательных бактерий это наружное L-кольцо ( липополисахарид ) и Р-кольцо ( пептидогликан ), а также внутреннее MS-кольцо (мембранное/надмембранное) и С-кольцо. ( цитоплазматический ). У грамположительных бактерий имеются только внутренние кольца. [29] Белки Mot ( MotA и MotB ) окружают внутренние кольца цитоплазматической мембраны ; Транслокация ионов через белки Mot обеспечивает энергию для вращения жгутиков. [26] Белки Fli позволяют менять направление вращения жгутиков в ответ на определенные стимулы. [30] [31] Крючок соединяет нить с моторным белком в основании. Спиральная нить состоит из множества копий белка флагеллина и может вращаться по часовой стрелке (CW) и против часовой стрелки (CCW). [32] [33] [34] [35] [3]
волосы (челка)
[ редактировать ]

Пилюс « ( лат. волосы») — это волосообразный придаток, обнаруженный на поверхности многих бактерий и архей . [37] Термины «пилус» и «бахромка » (лат. «бахрома») могут использоваться как взаимозаменяемые, хотя некоторые исследователи оставляют термин «пилус» для обозначения придатка, необходимого для бактериальной конъюгации . На поверхности бактерий и архей могут существовать десятки таких структур.
Подергивающаяся подвижность — это форма ползающей подвижности бактерий, используемая для перемещения по поверхностям. Подергивание опосредовано активностью пилуса определенного типа, называемого пилусом IV типа , который выходит из внешней части клетки, связывается с окружающими твердыми субстратами и втягивается, вытягивая клетку вперед, подобно действию крюка-кошки . [38] [39] [40] Пили используются не только для твичинга. Они также являются антигенными и необходимы для образования биопленки, поскольку прикрепляют бактерии к поверхности хозяина для колонизации во время инфекции. Они хрупкие и постоянно заменяются, иногда пилями разного состава. [41]
Другой
[ редактировать ]Скользящая подвижность — это тип транслокации, который не зависит от двигательных структур, таких как жгутики или пили . [42] Скольжение позволяет микроорганизмам перемещаться по поверхности пленок с низким содержанием воды. Механизмы этой подвижности известны лишь частично. Скользящая моторика использует весьма разнообразный набор различных двигательных комплексов, включая, например, фокальной адгезии комплексы Myxococcus . [43] [44] Скорость скольжения варьируется у разных организмов, а изменение направления, по-видимому, регулируется какими-то внутренними часами. [45]
Способы передвижения
[ редактировать ]
Большинство палочковидных бактерий могут передвигаться за счет собственной силы, что позволяет колонизировать новые среды обитания и открывать новые ресурсы для выживания. Передвижение бактерий зависит не только от характеристик среды, но и от использования различных придатков для движения. Движения роения и плавания приводятся в движение вращающимися жгутиками. [14] [46] Если роение представляет собой двумерное многоклеточное движение по поверхности и требует присутствия поверхностно-активных веществ, то плавание — это перемещение отдельных клеток в жидкой среде. [47] [3]
Другие типы движений, происходящих на твердых поверхностях, включают подергивания, скольжение и скольжение, которые не зависят от жгутиков. Подергивающаяся подвижность зависит от растяжения, прикрепления к поверхности и втягивания пилей IV типа , которые обеспечивают энергию, необходимую для продвижения клетки вперед. [48] Скользящая моторика использует весьма разнообразный набор различных двигательных комплексов, включая, например, фокальной адгезии комплексы Myxococcus . [43] [49] В отличие от подергивающихся и скользящих движений, которые являются активными движениями, в которых движущая сила создается отдельной клеткой, скольжение является пассивным движением. Он основан на движущей силе, генерируемой клеточным сообществом за счет расширяющих сил, вызванных ростом клеток внутри колонии в присутствии поверхностно-активных веществ, которые уменьшают трение между клетками и поверхностью. [50] [3]
Плавание
[ редактировать ]
| Внешние видео | |
|---|---|
Многие бактерии плавают за счет вращения жгутиков вне тела клетки. В отличие от жгутиков протистов , бактериальные жгутики представляют собой роторы и — независимо от вида и типа жгутиков — имеют только два режима работы: вращение по часовой стрелке (CW) или против часовой стрелки (CCW). Плавание бактерий используется в бактериальном такси (опосредованном специфическими рецепторами и путями передачи сигнала ), чтобы бактерия направленно двигалась по градиентам и достигала более благоприятных условий для жизни. [51] [52] Направление вращения жгутиков контролируется типом молекул, обнаруживаемых рецепторами на поверхности клетки: при наличии аттрактантного градиента скорость плавного плавания увеличивается, а при наличии репеллентного градиента увеличивается скорость кувыркания. . [53] [3]
Архетип бактериального плавания представлен хорошо изученным модельным организмом Escherichia coli . [3] Благодаря перитрихическому жгутикованию E. coli выполняет модель плавания «беги и кувыркайся» , как показано на схеме справа. Вращение жгутиковых моторов против часовой стрелки приводит к образованию жгутиковых пучков, которые толкают клетку вперед, параллельно длинной оси клетки. Вращение по часовой стрелке разбирает пучок, и ячейка вращается случайным образом (переворачиваясь). После кувырка прямое плавание восстанавливается в новом направлении. [53] То есть вращение против часовой стрелки приводит к устойчивому движению, а вращение по часовой – к кувырку; Вращение против часовой стрелки в заданном направлении сохраняется дольше в присутствии интересующих молекул (например, сахаров или аминокислот). [53] [3]

Пример: Vibrio alginolyticus.
Адаптировано из Son et al., 2013 г. [54]
Однако тип плавательного движения (приводимый в движение вращением жгутиков вне тела клетки) значительно варьируется в зависимости от вида и количества/распределения жгутиков на теле клетки. Например, морская бактерия Vibrio alginolyticus с единственным полярным жгутиком плавает по циклическому трехэтапному принципу (вперед, назад и движение). Плавание вперед происходит, когда жгутик толкает головку клетки, тогда как плавание назад основано на том, что жгутик тянет головку при реверсе движения. [3]

Пример: Rhodobacter sphaeroides.
Адаптировано из книги «Армитаж и Макнаб», 1987 г.; [55] Армитидж и др., 1999. [56]

Пример: Pseudomonas putida.
Адаптировано из Hintsche et al., 2017. [57]
Помимо этих разворотов на 180 °, ячейки могут переориентироваться («перемахивать») на угол около 90 °, что называется поворотом за счет выпучивания. [58] [54] Rhodobacter sphaeroides с его субполярным монотриховым жгутиком представляет собой еще одну стратегию подвижности: [55] [24] жгутик вращается только в одном направлении, время от времени он останавливается и прижимается к телу клетки, что приводит к переориентации тела клетки, [56] [59] [60] У почвенной бактерии Pseudomonas putida к ее заднему полюсу прикрепляется пучок спиральных жгутиков. P. putida чередует три режима плавания: толкание, вытягивание и обхватывание. [57] [3]
В режиме толкания вращающиеся жгутики (собранные в пучок или в виде открытого пучка отдельных нитей) приводят в движение задний конец тела клетки. Траектории либо прямые, либо, вблизи твердой поверхности, изогнуты вправо из-за гидродинамического взаимодействия ячейки с поверхностью. Направление кривизны указывает на то, что толкатели приводятся в движение левой спиралью, вращающейся против часовой стрелки. В тянущем режиме вращающийся пучок жгутиков направлен вперед. В этом случае траектории либо прямые, либо с тенденцией к изгибу влево, что указывает на то, что пуллеры плывут, поворачивая левовинтовой пучок по часовой стрелке. Наконец, P. putida может плавать, обертывая пучок нитей вокруг тела своей клетки, причем задний полюс указывает в направлении движения. В этом случае жгутиковый пучок принимает форму левой спирали, поворачивающейся по часовой стрелке, а траектории движения преимущественно прямые. [57] [3]
Роение
[ редактировать ]Роевая подвижность — это быстрое (2–10 мкм/с) и скоординированное перемещение бактериальной популяции по твердым или полутвердым поверхностям. [61] и является примером бактериальной многоклеточности и роевого поведения . Впервые о роевой подвижности сообщил в 1972 году Йорген Хенрихсен. [62]

Переход от плавательной к роевой подвижности обычно связан с увеличением числа жгутиков на клетку, сопровождающимся удлинением клеток. [63] Эксперименты с Proteus mirabilis показали, что для роения необходим контакт между клетками: роящиеся клетки движутся бок о бок группами, называемыми плотами, которые динамически добавляют или теряют клетки: когда клетка остается позади плота, ее движение через короткое время прекращается; когда группа клеток, движущихся в плоту, вступает в контакт с неподвижной клеткой, она реактивируется и включается в плот. [64] Совсем недавно Свеницки и его коллеги разработали полимерную микрофлюидную систему для ограничения клеток E. coli в квазидвумерном слое подвижного буфера, чтобы изучить различное поведение клеток, переходящих от плавания к роенству. [65] Для этого они заставили клетки E. coli планктонные приобрести фенотип роящихся клеток, ингибируя деление клеток (что приводит к удлинению клеток) и удаляя хемосенсорную систему (что приводит к плавному плаванию клеток, которые не переворачиваются). Увеличение плотности бактерий внутри канала привело к образованию все более крупных рафтов. Клетки, сталкивающиеся с плотом, способствовали увеличению его размеров, а клетки, движущиеся со скоростью, отличной от средней скорости внутри плота, отделялись от него. [65] [3]
Траектории клеток и движение жгутиков во время роения были тщательно изучены для E. coli в сочетании с флуоресцентно меченными жгутиками. [66] [46] Авторы описали четыре различных типа следов во время роения бактерий: движение вперед, разворот, боковое движение и стойло. [46] При движении вперед длинная ось клетки, жгутиковый пучок и направление движения совпадают, и движение аналогично движению свободно плавающей клетки. При реверсировании жгутиковый пучок ослабевает, при этом нити в пучке изменяются из своей «нормальной формы» (левосторонние спирали) в «кудрявую» форму правосторонних спиралей с меньшим шагом и амплитудой. Не меняя своей ориентации, тело клетки движется назад через разрыхленный пучок. Пучок заново формируется из вьющихся нитей на противоположном полюсе тела клетки, и нити в конечном итоге возвращаются к своей нормальной форме. Боковое движение может быть вызвано столкновениями с другими клетками или реверсом двигателя. Наконец, остановившиеся клетки приостанавливаются, но жгутики продолжают вращаться и перекачивать жидкость перед роем, обычно по краю роя. [46] [3]
Подергивание
[ редактировать ]
Подергивающаяся подвижность — это форма ползающей подвижности бактерий, используемая для перемещения по поверхностям. Подергивание опосредовано активностью волосообразных нитей, называемых пилями IV типа , которые выходят из внешней части клетки, связываются с окружающими твердыми субстратами и втягиваются, вытягивая клетку вперед, подобно действию крюка -кошки . [38] [68] [69] Название «подергивающаяся подвижность» происходит от характерных прерывистых и нерегулярных движений отдельных клеток, если смотреть под микроскопом. [70]
Бактериальная биопленка представляет собой бактериальное сообщество, прикрепленное к поверхности посредством внеклеточных полимерных материалов . [71] Прежде чем сформировать биопленку, бактериям, возможно, придется отложиться на поверхности из планктонного состояния. После того, как бактерии осаждаются на поверхности, они могут «дергаться» или ползать по поверхности, используя придатки, называемые пилями IV типа, чтобы «исследовать» субстрат в поисках подходящих мест для роста и, таким образом, образования биопленки. [72] [73] [74] [75] Пили исходят от поверхности бактерий и могут достигать нескольких микрометров в длину (хотя их нанометры ). диаметр составляет [76] Бактериальное подергивание происходит в результате циклов полимеризации и деполимеризации пилей IV типа. [77] [78] Полимеризация приводит к удлинению ворса и, в конечном итоге, к его прикреплению к поверхностям. Деполимеризация заставляет пилус втягиваться и отсоединяться от поверхности. Втягивание пилей создает на бактерию тянущие силы, которые будут тянуться в направлении векторной суммы сил пилей, что приводит к резким движениям. Типичный пилус типа IV может создавать силу, превышающую 100 пиконьютонов. [79] и тогда пучок пилей может создавать силу тяги до нескольких наноньютонов . [80] Бактерии могут использовать пили не только для подергивания, но и для межклеточных взаимодействий. [81] [82] поверхностное зондирование, [83] [84] и поглощение ДНК. [85] [67]
Скольжение
[ редактировать ]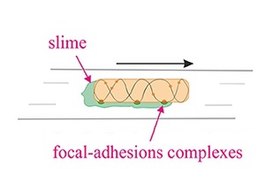
Скользящая подвижность — это тип транслокации, который не зависит от двигательных структур, таких как жгутики или пили . [42] Скольжение позволяет микроорганизмам перемещаться по поверхности пленок с низким содержанием воды. Механизмы этой подвижности известны лишь частично. Скорость скольжения варьируется у разных организмов, а изменение направления, по-видимому, регулируется какими-то внутренними часами. [45] Например, апикомплексаны способны перемещаться с высокой скоростью от 1 до 10 мкм/с. Напротив, Myxococcus xanthus , слизистая бактерия, может скользить со скоростью 5 мкм/мин. [86] [87] В миксобактериях отдельные бактерии перемещаются вместе, образуя волны клеток, которые затем дифференцируются, образуя плодовые тела, содержащие споры. [88] Миксобактерии перемещаются только на твердых поверхностях, в отличие, скажем, от кишечной палочки , которая подвижна в жидких или твердых средах. [89]
Неподвижный
[ редактировать ]Неподвижным видам не хватает способностей и структур, которые позволили бы им самостоятельно передвигаться по окружающей среде. Когда неподвижные бактерии культивируют в колотой пробирке, они растут только вдоль линии укола. Если бактерии подвижны, линия будет выглядеть размытой и распространяться в среду. [90]
Бактериальные такси: направленное движение
[ редактировать ]Говорят, что бактерии проявляют такси , если они движутся к какому-либо стимулу в окружающей среде или от него. Такое поведение позволяет бактериям менять свое положение по отношению к раздражителю. В зависимости от природы стимула, контролирующего направленное движение, можно выделить различные типы такси, такие как хемотаксис (химические градиенты, такие как глюкоза), аэротаксис (кислород), фототаксис (свет), термотаксис (тепло) и магнитотаксис (магнитные поля). . [3]
Хемотаксис
[ редактировать ]Общее движение бактерии может быть результатом чередования фаз падения и плавания. [91] В результате траектория движения бактерии, плавающей в однородной среде, будет представлять собой случайное блуждание с относительно прямым плаванием, прерываемым случайными падениями, которые переориентируют бактерию. [92] Бактерии, такие как E. coli, не могут выбирать направление своего плавания и не могут плавать по прямой более нескольких секунд из-за вращательной диффузии ; другими словами, бактерии «забывают» направление, в котором они движутся. Постоянно оценивая свой курс и корректируя свое движение в неправильном направлении, бактерии могут направить свое случайное движение в сторону благоприятных мест. [93]
При наличии химического градиента бактерии будут хемотаксировать или направлять свое общее движение в зависимости от градиента. Если бактерия почувствует, что движется в правильном направлении (к аттрактанту/от репеллента), она будет продолжать плыть по прямой в течение более длительного времени, прежде чем упасть; однако, если он движется в неправильном направлении, он упадет раньше. Бактерии, подобные E. coli, используют временное зондирование, чтобы решить, улучшается ли их ситуация или нет, и таким образом находят место с самой высокой концентрацией аттрактанта, обнаруживая даже небольшие различия в концентрации. [94]
Это предвзятое случайное блуждание является результатом простого выбора между двумя методами случайного движения; а именно акробатика и прямое плавание. [95] Спиральная природа отдельной жгутиковой нити имеет решающее значение для возникновения этого движения. Белковая структура, составляющая жгутиковую нить, флагеллин , консервативна среди всех жгутиковых бактерий. Позвоночные животные, похоже, воспользовались этим фактом, обладая иммунным рецептором ( TLR5 ), предназначенным для распознавания этого консервативного белка.
Как и во многих случаях в биологии, существуют бактерии, которые не следуют этому правилу. Многие бактерии, такие как вибрионы , моножгутиковые и имеют один жгутик на одном полюсе клетки. Их метод хемотаксиса различен. У других есть единственный жгутик, который находится внутри клеточной стенки. Эти бактерии передвигаются, вращая всю клетку, имеющую форму штопора. [96]
Способность морских микробов перемещаться к химическим горячим точкам может определять поглощение ими питательных веществ и потенциально влиять на круговорот элементов в океане. Связь между бактериальной навигацией и круговоротом питательных веществ подчеркивает необходимость понимания того, как функционирует хемотаксис в контексте морской микросреды. Хемотаксис зависит от стохастического связывания/развязывания молекул с поверхностными рецепторами, передачи этой информации через внутриклеточный сигнальный каскад , а также активации и контроля жгутиковых моторов. Присущая этим процессам случайность является главной проблемой, с которой клетки должны справляться, чтобы ориентироваться, особенно в условиях разбавления, когда шум и сигнал одинаковы по величине. Такие условия повсеместно распространены в океане, где концентрации питательных веществ часто чрезвычайно низки и подвержены быстрым изменениям в пространстве (например, твердые частицы, шлейфы питательных веществ) и времени (например, диффузия источников, смешивание жидкостей). [97]
Мелкомасштабные взаимодействия между морскими бактериями и растворенными и твердыми органическими веществами лежат в основе морской биогеохимии, тем самым поддерживая продуктивность и влияя на хранение и секвестрацию углерода в океанах планеты. [98] Исторически было очень сложно охарактеризовать морскую среду на микромасштабе, который наиболее подходит для отдельных бактерий. Скорее, исследовательские усилия обычно отбирали гораздо большие объемы воды и сравнивали разные места отбора проб с другим. [99] [100] Однако в масштабах, соответствующих отдельным микробам, океан представляет собой сложный и динамичный ландшафт с участками питательных веществ, порой слишком маленькими, чтобы их можно было перемешать турбулентностью. [101] [102] Способность микробов активно перемещаться в этой структурированной среде с помощью хемотаксиса может сильно влиять на поглощение ими питательных веществ. Хотя в некоторых работах изучались химические профили, зависящие от времени, [103] Прошлые исследования хемотаксиса с использованием E. coli и других модельных организмов регулярно исследовали устойчивые химические градиенты, достаточно сильные, чтобы вызвать заметный хемотаксический ответ. [104] [105] Однако типичные химические градиенты, с которыми сталкиваются дикие морские бактерии, часто очень слабы, эфемерны по своей природе и имеют низкие фоновые концентрации. [102] Мелкие градиенты актуальны для морских бактерий, потому что, как правило, градиенты становятся слабее по мере удаления от источника. Тем не менее, обнаружение таких градиентов на расстоянии имеет огромную ценность, поскольку они указывают на источники питательных веществ. Мелкие градиенты важны именно потому, что их можно использовать для навигации к регионам вблизи источников, где градиенты становятся крутыми, концентрации высоки, а бактерии могут приобретать ресурсы с высокой скоростью. [97]
Фототакси
[ редактировать ]Фототаксис — это разновидность такси или локомоторного движения, которое происходит, когда весь организм или от стимулу движется к световому него . [106] Это выгодно для фототрофных организмов, поскольку они могут наиболее эффективно ориентироваться на получение света для фотосинтеза . Фототаксис называется положительным, если движение происходит в направлении увеличения силы света, и отрицательным, если направление противоположное. [107]
наблюдаются два типа положительного фототаксиса У прокариот . Первый называется «скотофоботаксис» (от слова « скотофобия »), который наблюдается только под микроскопом. Это происходит, когда бактерия случайно выплывает из области, освещаемой микроскопом. Наступление темноты сигнализирует клетке о необходимости изменить направление вращения жгутиков и снова войти в свет. Второй тип фототаксиса — это истинный фототаксис, который представляет собой направленное движение вверх по градиенту к увеличению количества света. Это аналогично положительному хемотаксису, за исключением того, что аттрактант представляет собой свет, а не химическое вещество.
Фототаксические реакции наблюдаются у ряда бактерий и архей, таких как Serratia marcescens . Белки фоторецепторов представляют собой светочувствительные белки, участвующие в восприятии света и реакции на него у различных организмов. Некоторыми примерами являются бактериородопсин и бактериофитохромы у некоторых бактерий. См. также: Фитохром и фототропизм .
Большинство прокариот (бактерий и архей) неспособны определять направление света, потому что в таком маленьком масштабе очень сложно создать детектор, который мог бы различать одно направление света. Тем не менее, прокариоты могут измерять интенсивность света и двигаться по градиенту интенсивности света. Некоторые планирующие нитевидные прокариоты могут даже чувствовать направление света и совершать направленные повороты, но их фототаксическое движение очень медленное. Некоторые бактерии и археи фототаксичны. [108] [109] [110]
В большинстве случаев механизм фототаксиса представляет собой смещенное случайное блуждание, аналогичное бактериальному хемотаксису. Галофильные археи, такие как Halobacterium salinarum , используют сенсорные родопсины (SR) для фототаксиса. [111] [112] Родопсины — это семь трансмембранных белков, которые связывают ретиналь в качестве хромофора . Свет запускает изомеризацию сетчатки, [113] что приводит к фототрансдуктивной передаче сигналов через двухкомпонентную релейную систему фосфопереноса. Halobacterium salinarum имеет два SR, SRI и SRII, которые передают сигнал через белки-преобразователи HtrI и HtrII (галобактериальные преобразователи для SR I и II) соответственно. [114] [115] Нижестоящая передача сигналов у фототаксических архебактерий включает CheA, гистидинкиназу , которая фосфорилирует регулятор ответа CheY. [116] Фосфорилированный CheY вызывает разворот плавания. Два SR в Halobacterium имеют разные функции. SRI действует как аттрактантный рецептор для оранжевого света и, посредством двухфотонной реакции, как репеллентный рецептор для ближнего УФ-излучения, тогда как SRII является репеллентным рецептором для синего света. В зависимости от того, какой рецептор экспрессируется, если клетка плывет вверх или вниз по крутому градиенту света, вероятность переключения жгутика будет низкой. Если интенсивность света постоянна или меняется в неправильном направлении, переключение направления вращения жгутиков переориентирует клетку в новом, случайном направлении. [117] Поскольку длина треков увеличивается, когда клетка следует световому градиенту, клетки в конечном итоге будут приближаться к источнику света или удаляться от него. Эта стратегия не позволяет ориентироваться по вектору света и работает только при наличии крутого градиента света (т. е. не в открытой воде). [110]
Некоторые цианобактерии (например, Anabaena , Synechocystis ) способны медленно ориентироваться вдоль вектора света. Эта ориентация происходит в нитях или колониях, но только на поверхности, а не в суспензии. [118] [119] Нитчатая цианобактерия Synechocystis способна как к положительной, так и к отрицательной двумерной фототаксической ориентации. Положительный ответ, вероятно, опосредован бактериофитохромным фоторецептором TaxD1. Этот белок имеет два хромофорсвязывающих домена GAF, которые связывают биливердина . хромофор [120] и С-концевой домен, типичный для бактериальных рецепторов таксиса ( сигнальный домен MCP ). TaxD1 также имеет два N-концевых трансмембранных сегмента, которые закрепляют белок на мембране. [121] [122] [123] Фоторецепторный и сигнальный домены являются цитоплазматическими и передают сигнал через систему передачи сигналов типа CheA/CheY, регулируя подвижность пилей IV типа. [124] TaxD1 локализуется на полюсах палочковидных клеток Synechococcus elongatus , аналогично МСР, содержащим хемосенсорные рецепторы у бактерий и архей. [125] Как достигается управление нитями, неизвестно. Медленное управление этими цианобактериальными нитями — единственное поведение прокариот по распознаванию направления света, которое могло развиться из-за сложности определения направления света в таком небольшом масштабе. [110]


Магнитотаксис
[ редактировать ]Магнитотактические бактерии ориентируются вдоль силовых линий Земли магнитного поля . [127] Считается, что такое выравнивание помогает этим организмам достичь областей с оптимальной концентрацией кислорода. [128] Для выполнения этой задачи у этих бактерий есть биоминерализованные органеллы, называемые магнитосомами , которые содержат магнитные кристаллы . Биологическое явление микроорганизмов, стремящихся двигаться в ответ на магнитные характеристики окружающей среды, известно как магнитотаксис . Однако этот термин вводит в заблуждение, поскольку любое другое его применение предполагает наличие механизма стимул-реакция. В отличие от магниторецепции животных, бактерии содержат фиксированные магниты, которые заставляют бактерии выравниваться — даже мертвые клетки притягиваются к ним, как стрелка компаса. [128]
Ответ на побег
[ редактировать ]
Размерная полоса = 270 нм
Реакция бегства — это форма негативного такси. Стимулы, которые могут нанести вред или убить, требуют быстрого обнаружения. Это принципиально отличается от навигации или разведки с точки зрения временных рамок, доступных для реагирования. Большинство подвижных видов обладают формой фобической или экстренной реакции, отличной от их устойчивого передвижения. [129] Реакции бегства не являются строго ориентированными, но обычно включают движение назад, иногда с негативным геотаксическим компонентом. [130] [129] У бактерий и архей потенциалу действия. наблюдались явления, подобные в биопленках [131] а также отдельные клетки, такие как кабельные бактерии . [129] Архей Halobacterium salinarium демонстрирует фотофобную реакцию, характеризующуюся изменением направления плавания на 180 °, вызванным изменением направления вращения жгутиков. По крайней мере, некоторые аспекты этого ответа, вероятно, опосредованы изменениями мембранного потенциала бактериородопсина управляемого светом , протонного насоса, . [132] Явления, подобные потенциалу действия, у прокариот отличаются от классических потенциалов действия эукариот. Первые менее воспроизводимы, медленнее и имеют более широкое распределение амплитуды и длительности импульса. [133] [129]
Другие налоги
[ редактировать ]- Аэротаксис — это реакция организма на изменение концентрации кислорода, которая в основном встречается у аэробных бактерий. [106]
- Энергетическое такси — это ориентация бактерий на условия оптимальной метаболической активности путем определения внутреннего энергетического состояния клетки. Следовательно, в отличие от хемотаксиса (таксис к определенному внеклеточному соединению или от него), энергетическое такси реагирует на внутриклеточный стимул (например, движущую силу протонов , активность NDH-1 ) и требует метаболической активности. [134]
Математическое моделирование
[ редактировать ]
плавательного поведения Serratia marcescens
(а) без и (б) с присутствием хемоаттрактанта [137]
Математические модели, используемые для описания динамики плавания бактерий, можно разделить на две категории. Первая категория основана на микроскопическом (то есть на клеточном уровне) взгляде на плавание бактерий с помощью набора уравнений, где каждое уравнение описывает состояние отдельного агента. [138] [139] [140] [141] [142] Вторая категория обеспечивает макроскопический взгляд (т.е. на уровне популяции) посредством уравнений в частных производных, основанных на континууме, которые фиксируют динамику плотности населения в пространстве и времени, без непосредственного рассмотрения внутриклеточных характеристик. [143] [144] [145] [146] [147] [148] [149] [150] [151] [137]
Среди существующих моделей Шнитцер использует уравнение Смолуховского для описания смещенного случайного блуждания бактерий во время хемотаксиса в поисках пищи. [152] Чтобы сосредоточиться на подробном описании движения бактерий, происходящего в течение одного интервала пробега, де Жен вычисляет среднюю длину пробега, пройденную бактериями за один интервал против часовой стрелки. [153] В том же направлении, чтобы рассмотреть условия окружающей среды, влияющие на смещенное случайное блуждание бактерий, Кроз и его коллеги экспериментально и теоретически изучают влияние концентрации мягкого агара на хемотаксис бактерий. [154] [137]
Чтобы изучить влияние препятствий (еще одного условия окружающей среды) на движение бактерий, Чепижко и его сотрудники изучают движение самодвижущихся частиц в гетерогенной двумерной среде и показывают, что среднеквадратичное смещение частиц зависит от плотность препятствий и скорость вращения частицы. [154] [155] Опираясь на эти модели, Кейтс подчеркивает, что динамика бактерий не всегда подчиняется детальному балансу, а это означает, что это процесс смещенной диффузии, зависящий от условий окружающей среды. [156] Более того, Ариэль и его коллеги сосредоточились на диффузии бактерий и показали, что бактерии осуществляют супердиффузию во время роения на поверхности. [157] [137]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Браун, Джеймс Ф. (1852). «XXIV. О некоторых солях и продуктах разложения пиромеконовой кислоты». Лондонский, Эдинбургский и Дублинский философский журнал и научный журнал . 4 (24): 161–168. дои : 10.1080/14786445208647098 .
- ^ Эйнштейн, А. (1905). «О движении частиц, взвешенных в покоящихся жидкостях, требуемых молекулярно-кинетической теорией теплоты» . Анналы физики . 322 (8): 549–560. Бибкод : 1905АнП...322..549Е . дои : 10.1002/andp.19053220806 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также Бастос-Аррьета, Хулио; Ревилла-Гуаринос, Эноа; Успал, Уильям Э.; Зиммхен, Джулиана (2018). «Бактериальные биогибридные микропловцы» . Границы робототехники и искусственного интеллекта . 5 : 97. дои : 10.3389/frobt.2018.00097 . ПМЦ 7805739 . ПМИД 33500976 .
 Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Перейти обратно: а б с д и ж Перселл, Э.М. (1977). «Жизнь при низком числе Рейнольдса». Американский журнал физики . 45 (1): 3–11. Бибкод : 1977AmJPh..45....3P . дои : 10.1119/1.10903 .
- ^ Цю, Тянь; Ли, Тунг-Чун; Марк, Эндрю Г.; Морозов Константин Игоревич; Мюнстер, Рафаэль; Мирка, Отто; Турек, Стефан; Лешанский Александр М.; Фишер, Пер (2014). «Плавание возвратно-поступательным движением при малых числах Рейнольдса» . Природные коммуникации . 5 : 5119. Бибкод : 2014NatCo...5.5119Q . дои : 10.1038/ncomms6119 . ПМК 4241991 . ПМИД 25369018 .
- ^ Лауга, Эрик; Пауэрс, Томас Р. (2009). «Гидродинамика плавающих микроорганизмов». Отчеты о прогрессе в физике . 72 (9): 096601. arXiv : 0812.2887 . Бибкод : 2009RPPH...72i6601L . дои : 10.1088/0034-4885/72/9/096601 . S2CID 3932471 .
- ^ Фогель, Пиа Д. (2005). «Природный дизайн наномоторов». Европейский журнал фармацевтики и биофармацевтики . 60 (2): 267–277. дои : 10.1016/j.ejpb.2004.10.007 . ПМИД 15939237 .
- ^ Патра, Дебабрата; Сенгупта, Самудра; Дуань, Вэньтао; Чжан, Хуа; Павлик, Райан; Сен, Аюсман (2013). «Интеллектуальные автономные системы доставки лекарств». Наномасштаб . 5 (4): 1273–1283. Бибкод : 2013Nanos...5.1273P . дои : 10.1039/C2NR32600K . ПМИД 23166050 .
- ^ Феринга, Бен Л. (2001). «Управление движением: от молекулярных переключателей к молекулярным двигателям» . Отчеты о химических исследованиях . 34 (6): 504–513. дои : 10.1021/ar0001721 . hdl : 11370/a0b20090-34b9-4e2d-8450-bc2afbea2fcf . ПМИД 11412087 .
- ^ Соколов А.; Аподака, ММ; Гжибовский, бакалавр; Арансон, И.С. (2010). «Плавающие бактерии приводят в действие микроскопические механизмы» . Труды Национальной академии наук . 107 (3): 969–974. Бибкод : 2010PNAS..107..969S . дои : 10.1073/pnas.0913015107 . ПМЦ 2824308 . ПМИД 20080560 .
- ^ Мэдиган, Майкл Т.; Бендер, Келли С.; Бакли, Дэниел Х.; Брок, Томас Д.; Мэтью Сэттли, В.; Шталь, Дэвид Аллан (29 января 2018 г.). Брок Биология микроорганизмов . Пирсон. ISBN 9781292235103 .
- ^ Дюфрен, Ив Ф. (2015). «Липкие микробы: силы адгезии микробных клеток». Тенденции в микробиологии . 23 (6): 376–382. дои : 10.1016/j.tim.2015.01.011 . ПМИД 25684261 .
- ^ Перейти обратно: а б Баркер, Клайв С.; Мещерякова Ирина Владимировна; Костюкова Алла С.; Фреддолино, Питер Л.; Саматей, Фадель А. (27 октября 2017 г.). «Внутренне неупорядоченный линкер, контролирующий образование и стабильность бактериального жгутикового крючка» . БМК Биология . 15 (1). Springer Science and Business Media LLC: 97. doi : 10.1186/s12915-017-0438-7 . ISSN 1741-7007 . ПМК 5660449 . ПМИД 29078764 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б Берг, Ховард К.; Андерсон, Роберт А. (1973). «Бактерии плавают, вращая жгутиковые нити». Природа . 245 (5425): 380–382. Бибкод : 1973Natur.245..380B . дои : 10.1038/245380a0 . ПМИД 4593496 . S2CID 4173914 .
- ^ Перейти обратно: а б Альберс С.В., Джаррелл К.Ф. (27 января 2015 г.). «Архееллум: как плавают археи» . Границы микробиологии . 6:23 . дои : 10.3389/fmicb.2015.00023 . ПМЦ 4307647 . ПМИД 25699024 .
- ^ Сильверман М., Саймон М. (май 1974 г.). «Вращение жгутиков и механизм подвижности бактерий». Природа . 249 (452): 73–4. Бибкод : 1974Natur.249...73S . дои : 10.1038/249073a0 . ПМИД 4598030 . S2CID 10370084 .
- ^ Мейстер Г.Л., Берг Х.К. (1987). «Быстрое вращение жгутиковых пучков у плавающих бактерий». Природа . 325 (6105): 637–640. Бибкод : 1987Natur.325..637L . дои : 10.1038/325637a0 . S2CID 4242129 .
- ^ Берг Х.К., Андерсон Р.А. (октябрь 1973 г.). «Бактерии плавают, вращая свои жгутиковые нити». Природа . 245 (5425): 380–2. Бибкод : 1973Natur.245..380B . дои : 10.1038/245380a0 . ПМИД 4593496 . S2CID 4173914 .
- ^ Ян Т.Л., Бови ЕС (1965). «Передвижение и локомоция микроорганизмов». Ежегодный обзор микробиологии . 19 :21–58. дои : 10.1146/annurev.mi.19.100165.000321 . ПМИД 5318439 .
- ^ Харши РМ (2003). «Подвижность бактерий на поверхности: много путей к общей цели». Ежегодный обзор микробиологии . 57 : 249–73. дои : 10.1146/annurev.micro.57.030502.091014 . ПМИД 14527279 .
- ^ Джаррелл К. (2009). «Археальные жгутики и пили». Пили и жгутики: текущие исследования и будущие тенденции . Кайстер Академик Пресс. ISBN 978-1-904455-48-6 .
- ^ Штрайф С., Штаудингер В.Ф., Марван В., Остерхельт Д. (2008). «Вращение жгутиков у архей Halobacterium salinarum зависит от АТФ». Журнал молекулярной биологии . 384 (1): 1–8. дои : 10.1016/j.jmb.2008.08.057 . ПМИД 18786541 .
- ^ Ло, Ю-Ран (9 марта 2007 г.). Комплексный справочник по энергиям химической связи . ЦРК Пресс. ISBN 9781420007282 .
- ^ Перейти обратно: а б Хая, Шохей; Токумару, Юя; Абэ, Наоки; Канеко, Джун; Айзава, Син-Ичи (2011). «Характеристика боковых жгутиков Selenomonas ruminantium» . Прикладная и экологическая микробиология . 77 (8): 2799–2802. Бибкод : 2011ApEnM..77.2799H . дои : 10.1128/АЕМ.00286-11 . ПМК 3126368 . ПМИД 21335384 .
- ^ Брэди, Ричард М. (1993). «Крутящий момент и переключение бактериального жгутикового двигателя. Электростатическая модель» . Биофизический журнал . 64 (4): 961–973. Бибкод : 1993BpJ....64..961B . дои : 10.1016/S0006-3495(93)81462-0 . ПМЦ 1262414 . ПМИД 7684268 .
- ^ Перейти обратно: а б Мэнсон, доктор медицины; Тедеско, П.; Берг, ХК; Гарольд, FM; Ван Дер Дрифт, К. (1977). «Протонодвижущая сила приводит в движение жгутики бактерий» . Труды Национальной академии наук . 74 (7): 3060–3064. Бибкод : 1977PNAS...74.3060M . дои : 10.1073/pnas.74.7.3060 . ПМК 431412 . ПМИД 19741 .
- ^ Хирота, Норифуми; Китада, Макио; Имаэ, Иисус (1981). «Жгутиковые моторы алкалофильных бактерий, работающие за счет градиента электрохимического потенциала Na+» . Письма ФЭБС . 132 (2): 278–280. дои : 10.1016/0014-5793(81) 81178-7 S2CID 85138168 .
- ^ Элстон, Техас; Остер, Г. (1997). «Белковые турбины. I: Бактериальный жгутиковый мотор» . Биофизический журнал . 73 (2): 703–721. Бибкод : 1997BpJ....73..703E . дои : 10.1016/S0006-3495(97)78104-9 . ПМК 1180968 . ПМИД 9251788 .
- ^ Чен, Сонге; Биби, Морган; Мерфи, Гэвин Э.; Ледбеттер, Джаред Р.; Хендриксон, Дэвид Р.; Бригель, Ариана; Ли, Чжо; Ши, Цзянь; Точева, Элица И.; Мюллер, Аксель; Добро, Меган Дж.; Дженсен, Грант Дж. (2011). «Структурное разнообразие бактериальных жгутиковых моторов» . Журнал ЭМБО . 30 (14): 2972–2981. дои : 10.1038/emboj.2011.186 . ПМК 3160247 . ПМИД 21673657 .
- ^ Сокетт, Х.; Ямагучи, С.; Кихара, М.; Ирикура, В.М.; Макнаб, Р.М. (1992). «Молекулярный анализ белка-переключателя жгутиков FliM Salmonella typhimurium» . Журнал бактериологии . 174 (3): 793–806. дои : 10.1128/jb.174.3.793-806.1992 . ПМК 206156 . ПМИД 1732214 .
- ^ Уэлч, М.; Осава, К.; Айзава, С.; Айзенбах, М. (1993). «Зависимое от фосфорилирования связывание сигнальной молекулы с жгутиковым переключателем бактерий» . Труды Национальной академии наук . 90 (19): 8787–8791. Бибкод : 1993PNAS...90.8787W . дои : 10.1073/pnas.90.19.8787 . ПМК 47445 . ПМИД 8415608 .
- ^ Берг, Ховард К. (2003). «Вращающийся двигатель бактериальных жгутиков». Ежегодный обзор биохимии . 72 : 19–54. doi : 10.1146/annurev.biochem.72.121801.161737 . ПМИД 12500982 .
- ^ Эрхардт, М.; Намба, К.; Хьюз, КТ (2010). «Бактериальные наномашины: жгутик и инъекция III типа» . Перспективы Колд-Спринг-Харбор в биологии . 2 (11): а000299. doi : 10.1101/cshperspect.a000299 . ПМК 2964186 . ПМИД 20926516 .
- ^ Эванс, Льюис Д.Б.; Хьюз, Колин; Фрейзер, Джиллиан М. (2014). «Построение жгутика вне бактериальной клетки» . Тенденции в микробиологии . 22 (10): 566–572. дои : 10.1016/j.tim.2014.05.009 . ПМК 4183434 . ПМИД 24973293 .
- ^ Минамино, Тору; Имада, Кацуми (2015). «Бактериальный жгутиковый двигатель и его структурное разнообразие». Тенденции в микробиологии . 23 (5): 267–274. дои : 10.1016/j.tim.2014.12.011 . ПМИД 25613993 .
- ^ Гросс, Лиза (29 августа 2006 г.). «Бактериальные фимбрии, созданные для того, чтобы оставаться в потоке» . ПЛОС Биология . 4 (9). Публичная научная библиотека (PLoS): e314. дои : 10.1371/journal.pbio.0040314 . ISSN 1545-7885 . ПМЦ 1557401 . ПМИД 20076642 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ «пилус» в Медицинском словаре Дорланда.
- ^ Перейти обратно: а б Скеркер, Дж. М.; Берг, ХК (5 июня 2001 г.). «Прямое наблюдение за расширением и втягиванием пилей IV типа» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (12): 6901–6904. Бибкод : 2001PNAS...98.6901S . дои : 10.1073/pnas.121171698 . ISSN 0027-8424 . ПМК 34450 . ПМИД 11381130 .
- ^ Мэттик, Джон С. (2002). «Пили IV типа и подергивания моторики». Ежегодный обзор микробиологии . 56 : 289–314. дои : 10.1146/annurev.micro.56.012302.160938 . ISSN 0066-4227 . ПМИД 12142488 .
- ^ Мерц, AJ; Итак, М.; Шитц, член парламента (7 сентября 2000 г.). «Втягивание пилуса приводит к подергивающейся подвижности бактерий». Природа . 407 (6800): 98–102. Бибкод : 2000Natur.407...98M . дои : 10.1038/35024105 . ISSN 0028-0836 . ПМИД 10993081 . S2CID 4425775 .
- ^ Бринтон, Чарльз (1954). «Исследование электрофореза и чувствительности к фагам варианта бактерии E. coli, продуцирующего нити». Биохимика и биофизика Acta . 15 (4): 533–542. дои : 10.1016/0006-3002(54)90011-6 . ПМИД 13230101 .
- ^ Перейти обратно: а б Нан, Бэйян (февраль 2017 г.). «Скользящая подвижность бактерий: разработка консенсусной модели» . Современная биология . 27 (4): Р154–Р156. дои : 10.1016/j.cub.2016.12.035 . ПМИД 28222296 .
- ^ Перейти обратно: а б с Ислам, Салим Т.; Миньо, Там (2015). «Загадочная природа подвижности поверхности (скольжения) бактерий: механизм, основанный на фокальной адгезии, у Myxococcus xanthus». Семинары по клеточной биологии и биологии развития . 46 : 143–154. дои : 10.1016/j.semcdb.2015.10.033 . ПМИД 26520023 .
- ^ Нан, Бэйян; Зусман, Дэвид Р. (2016). «Новые механизмы обеспечения скользящей подвижности бактерий» . Молекулярная микробиология . 101 (2): 186–193. дои : 10.1111/mmi.13389 . ПМК 5008027 . ПМИД 27028358 .
- ^ Перейти обратно: а б Нан, Бэйян; Макбрайд, Марк Дж.; Чен, Цзин; Зусман, Дэвид Р.; Остер, Джордж (февраль 2014 г.). «Бактерии, скользящие по спиральным дорожкам» . Современная биология . 24 (4): 169–174. дои : 10.1016/j.cub.2013.12.034 . ПМЦ 3964879 . ПМИД 24556443 .
- ^ Перейти обратно: а б с д Вишничай, Гаштон; Франжипане, Джакомо; Магги, Клаудио; Сальимбени, Филиппо; Бьянки, Сильвио; Ди Леонардо, Роберто (2017). «Световые управляемые 3D-микромоторы, работающие на бактериях» . Природные коммуникации . 8 : 15974. Бибкод : 2017NatCo...815974V . дои : 10.1038/ncomms15974 . ПМЦ 5493761 . ПМИД 28656975 .
- ^ Хенрихсен, Дж. (1972) «Бактериальная поверхностная транслокация: обзор и классификация». Бактериол. Откр. , 36 : 478–503.
- ^ Мэттик, Джон С. (2002). «Пили IV типа и подергивания моторики». Ежегодный обзор микробиологии . 56 : 289–314. дои : 10.1146/annurev.micro.56.012302.160938 . ПМИД 12142488 .
- ^ Нан, Бэйян; Зусман, Дэвид Р. (2016). «Новые механизмы обеспечения скользящей подвижности бактерий» . Молекулярная микробиология . 101 (2): 186–193. дои : 10.1111/mmi.13389 . ПМК 5008027 . ПМИД 27028358 .
- ^ Хёльшер, Тереза; Ковач, Акош Т. (2017). «Скольжение по поверхности: распространение бактерий без активного двигателя». Экологическая микробиология . 19 (7): 2537–2545. дои : 10.1111/1462-2920.13741 . hdl : 11858/00-001M-0000-002D-2464-7 . ПМИД 28370801 .
- ^ Сова, Ёсиюки; Берри, Ричард М. (2008). «Бактериальный жгутиковый мотор» . Ежеквартальные обзоры биофизики . 41 (2): 103–132. дои : 10.1017/S0033583508004691 . ПМИД 18812014 . S2CID 3297704 .
- ^ Крелл, Тино; Лакаль, Хесус; Муньос-Мартинес, Франциско; Рейес-Дариас, Хосе Антонио; Кадирчи, Бильге Хилал; Гарсиа-Фонтана, Кристина; Рамос, Хуан Луис (2011). «Разнообразие в лучшем виде: бактериальные такси» . Экологическая микробиология . 13 (5): 1115–1124. дои : 10.1111/j.1462-2920.2010.02383.x . ПМИД 21087385 .
- ^ Перейти обратно: а б с Берг, Ховард К. (11 января 2008 г.). Кишечная палочка в движении . Спрингер. ISBN 9780387216386 .
- ^ Перейти обратно: а б Сын, Кванмин; Гуасто, Джеффри С.; Стокер, Роман (2013). «Бактерии могут использовать нестабильность изгиба жгутиков, чтобы изменить направление» . Физика природы . 9 (8): 494–498. Бибкод : 2013NatPh...9..494S . дои : 10.1038/nphys2676 .
- ^ Перейти обратно: а б Армитидж, JP; Макнаб, Р.М. (1987). «Однонаправленное прерывистое вращение жгутика Rhodobacter sphaeroides» . Журнал бактериологии . 169 (2): 514–518. дои : 10.1128/jb.169.2.514-518.1987 . ПМК 211807 . ПМИД 3492489 .
- ^ Перейти обратно: а б Армитидж, Джудит П.; Питта, Томас П.; Вигеант, Марго А.-С.; Пакер, Хелен Л.; Форд, Розанна М. (1999). «Преобразования в структуре жгутиков Rhodobacter sphaeroides и возможная связь с изменением скорости плавания» . Журнал бактериологии . 181 (16): 4825–4833. дои : 10.1128/JB.181.16.4825-4833.1999 . ПМК 93968 . ПМИД 10438751 .
- ^ Перейти обратно: а б с Хинче, Мариус; Валджор, Вероника; Гроссманн, Роберт; Кюн, Марко Дж.; Торманн, Кай М.; Перуани, Фернандо; Бета, Карстен (2017). «Полярный пучок жгутиков может стимулировать плавание бактерий, толкая, вытягивая или обвивая тело клетки» . Научные отчеты . 7 (1): 16771. Бибкод : 2017NatSR...716771H . дои : 10.1038/s41598-017-16428-9 . ПМК 5711944 . ПМИД 29196650 .
- ^ Се, Л.; Альтиндал, Т.; Чаттопадхьяй, С.; Ву, Х.-Л. (2011). «Бактериальный жгутик как пропеллер и руль направления для эффективного хемотаксиса» . Труды Национальной академии наук . 108 (6): 2246–2251. дои : 10.1073/pnas.1011953108 . ПМК 3038696 . ПМИД 21205908 .
- ^ Пилизота, Теута; Браун, Мостин Т.; Лик, Марк С.; Бранч, Ричард В.; Берри, Ричард М.; Армитидж, Джудит П. (2009). «Молекулярный тормоз, а не сцепление, останавливает сферический двигатель Rhodobacter sphaeroidesflagellar» . Труды Национальной академии наук . 106 (28): 11582–11587. Бибкод : 2009PNAS..10611582P . дои : 10.1073/pnas.0813164106 . ПМК 2710667 . ПМИД 19571004 .
- ^ Россер, Габриэль; Бейкер, Рут Э.; Армитидж, Джудит П.; Флетчер, Александр Г. (2014). «Моделирование и анализ бактериальных следов позволяют предположить активный механизм переориентации у Rhodobacter sphaeroides» . Журнал интерфейса Королевского общества . 11 (97). дои : 10.1098/rsif.2014.0320 . ПМК 4208361 . ПМИД 24872500 .
- ^ Харши, Расика М. (1 января 2003 г.). «Подвижность бактерий на поверхности: множество путей к общей цели». Ежегодный обзор микробиологии . 57 (1): 249–73. дои : 10.1146/annurev.micro.57.030502.091014 . ПМИД 14527279 .
- ^ Хенрихсен, Дж (1972). «Бактериальная поверхностная транслокация: обзор и классификация» . Бактериологические обзоры . 36 (4): 478–503. дои : 10.1128/BR.36.4.478-503.1972 . ПМК 408329 . ПМИД 4631369 .
- ^ Перейти обратно: а б Кернс, Дэниел Б. (2010). «Полевой справочник по подвижности бактерий» . Обзоры природы Микробиология . 8 (9): 634–644. дои : 10.1038/nrmicro2405 . ПМК 3135019 . ПМИД 20694026 .
- ^ Пак, Бён Вук; Чжуан, Цзян; Яса, Онкей; Ситти, Метин (2017). «Многофункциональные бактериальные микропловцы для адресной активной доставки лекарств». АСУ Нано . 11 (9): 8910–8923. дои : 10.1021/acsnano.7b03207 . ПМИД 28873304 .
- ^ Перейти обратно: а б Свечицкий, Жан-Мари; Слюсаренко Олеский; Вайбель, Дуглас Б. (2013). «От плавания к роению: подвижность клеток Escherichia coli в двух измерениях» . Интегративная биология . 5 (12): 1490–1494. дои : 10.1039/c3ib40130h . ПМЦ 4222179 . ПМИД 24145500 .
- ^ Тернер, Линда; Рю, Уильям С.; Берг, Ховард К. (2000). «Визуализация флуоресцентных жгутиковых нитей в реальном времени» . Журнал бактериологии . 182 (10): 2793–2801. дои : 10.1128/JB.182.10.2793-2801.2000 . ПМК 101988 . ПМИД 10781548 .
- ^ Перейти обратно: а б Джаятилаке, Пахала Гедара; Ли, Боуэн; Сулиани, Паоло; Кертис, Том; Чен, Чинджу (2019). «Моделирование бактериальных подергиваний в потоках жидкости: подход CFD-DEM» . Научные отчеты . 9 (1): 14540. Бибкод : 2019NatSR...914540J . дои : 10.1038/s41598-019-51101-3 . ПМЦ 6787227 . ПМИД 31601892 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Мэттик, Джон С. (2002). «Пили IV типа и подергивания моторики». Ежегодный обзор микробиологии . 56 : 289–314. дои : 10.1146/annurev.micro.56.012302.160938 . ISSN 0066-4227 . ПМИД 12142488 .
- ^ Мерц, AJ; Итак, М.; Шитц, член парламента (7 сентября 2000 г.). «Втягивание пилуса приводит к подергивающейся подвижности бактерий». Природа . 407 (6800): 98–102. Бибкод : 2000Natur.407...98M . дои : 10.1038/35024105 . ISSN 0028-0836 . ПМИД 10993081 . S2CID 4425775 .
- ^ Хенриксен, Дж. (декабрь 1972 г.). «Бактериальная поверхностная транслокация: обзор и классификация» . Бактериологические обзоры . 36 (4): 478–503. дои : 10.1128/BR.36.4.478-503.1972 . ISSN 0005-3678 . ПМК 408329 . ПМИД 4631369 .
- ^ о'Тул, Джордж А.; Вонг, Джерард CL (2016). «Сенсационные биопленки: поверхностное зондирование бактерий» . Современное мнение в микробиологии . 30 : 139–146. дои : 10.1016/j.mib.2016.02.004 . ПМЦ 4843124 . ПМИД 26968016 .
- ^ Майер, Беренике; Вонг, Джерард CL (2015). «Как бактерии используют оборудование для пилей типа IV на поверхностях». Тенденции в микробиологии . 23 (12): 775–788. дои : 10.1016/j.tim.2015.09.002 . ПМИД 26497940 .
- ^ Джин, Ф.; Конрад, JC; Гибиански, М.Л.; Вонг, GCL (2011). «Бактерии используют пили типа IV, чтобы выстреливать по поверхностям» . Труды Национальной академии наук . 108 (31): 12617–12622. дои : 10.1073/pnas.1105073108 . ПМК 3150923 . ПМИД 21768344 .
- ^ Брилл-Карниели, Ифат; Джин, Фан; Вонг, Джерард CL; Френкель, Даан; Добникар, Юре (2017). «Появление сложного поведения в подвижности пилей на ранних стадиях поверхностной адаптации P. Aeruginosa» . Научные отчеты . 7 : 45467. Бибкод : 2017NatSR...745467B . дои : 10.1038/srep45467 . ПМЦ 5385500 . ПМИД 28393835 .
- ^ Семмлер, Анналезе Б.Т.; Уитчерч, Синтия Б.; Мэттик, Джон С. (1999). «Повторное исследование подергивания моторики Pseudomonas aeruginosa» . Микробиология . 145 (10): 2863–2873. дои : 10.1099/00221287-145-10-2863 . ПМИД 10537208 .
- ^ Де Хаан, Хендрик В. (2016). «Моделирование и моделирование динамики распространения пилей IV типа Pseudomonas aeruginosa» . Биофизический журнал . 111 (10): 2263–2273. Бибкод : 2016BpJ...111.2263D . дои : 10.1016/j.bpj.2016.09.050 . ПМК 5112937 . ПМИД 27851948 .
- ^ Майер, Беренике (2013). «Бактериальная пилусная система IV типа – перестраиваемый молекулярный мотор» . Мягкая материя . 9 (24): 5667. Бибкод : 2013SMat....9.5667M . дои : 10.1039/c3sm50546d .
- ^ Скеркер, Дж. М.; Берг, ХК (2001). «Прямое наблюдение за расширением и втягиванием пилей IV типа» . Труды Национальной академии наук . 98 (12): 6901–6904. Бибкод : 2001PNAS...98.6901S . дои : 10.1073/pnas.121171698 . ПМК 34450 . ПМИД 11381130 .
- ^ Майер, Б.; Поттер, Л.; Итак, М.; Зейферт, HS; Шитц, член парламента; Шитц, член парламента (2002). «Сила одного пильного двигателя превышает 100 пН» . Труды Национальной академии наук . 99 (25): 16012–16017. Бибкод : 2002PNAS...9916012M . дои : 10.1073/pnas.242523299 . ПМК 138556 . ПМИД 12446837 .
- ^ Биэ, Николя; Ладу, Бенуа; Хигаши, Дастин; Итак, Магдалина; Шитц, Майкл (2008). «Совместное втягивание связанных пилей типа IV позволяет генерировать наноньютоновую силу» . ПЛОС Биология . 6 (4): е87. doi : 10.1371/journal.pbio.0060087 . ПМК 2292754 . ПМИД 18416602 .
- ^ Пениш, Вольфрам; Вебер, Кристоф А.; Юкеланд, Гвидо; Биэ, Николя; Забурдаев, Василий (2017). «Многомасштабное моделирование бактериальных колоний: как пили опосредуют динамику отдельных клеток и клеточных агрегатов» . Новый журнал физики . 19 (1): 015003. Бибкод : 2017NJPh...19a5003P . дои : 10.1088/1367-2630/aa5483 . ПМЦ 8132470 . ПМИД 34017216 .
- ^ Девентер, Лена; Олдевуртель, Энно Р.; Кузель, Надежда; Фолькманн, Торстен; Хенселер, Катя; Майер, Беренике (2016). «Силы дифференциального взаимодействия управляют сортировкой бактерий и стабильностью ранних биопленок» . Биофизический журнал . 110 (3): 469а. Бибкод : 2016BpJ...110..469D . дои : 10.1016/j.bpj.2015.11.2513 .
- ^ Эллисон, Кортни К.; Кан, Цзинбо; Диллард, Ребекка С.; Кисела, Дэвид Т.; Дюкре, Адриан; Берн, Сесиль; Хэмптон, Шери М.; Кэ, Цзуньлун; Райт, Элизабет Р.; Биэ, Николя; Далия, Анкур Б.; Брун, Ив В. (2017). «Препятствие ретракции пилуса стимулирует поверхностное зондирование бактерий» . Наука . 358 (6362): 535–538. Бибкод : 2017Sci...358..535E . дои : 10.1126/science.aan5706 . ПМК 5805138 . ПМИД 29074778 .
- ^ Персат, Александр; Инклан, Юки Ф.; Энгель, Джоан Н.; Стоун, Ховард А.; Гитай, Земер (2015). «Пили IV типа механохимически регулируют факторы вирулентности Pseudomonas aeruginosa» . Труды Национальной академии наук . 112 (24): 7563–7568. Бибкод : 2015PNAS..112.7563P . дои : 10.1073/pnas.1502025112 . ПМЦ 4475988 . ПМИД 26041805 .
- ^ Эллисон, Кортни К.; Далия, Триана Н.; Видаль Себальос, Альфредо; Ван, Джозеф Че-Йен; Биэ, Николя; Брун, Ив В.; Далия, Анкур Б. (2018). «Ретракция ДНК-связанных пилей IV типа инициирует поглощение ДНК во время естественной трансформации холерного вибриона» . Природная микробиология . 3 (7): 773–780. дои : 10.1038/s41564-018-0174-y . ПМК 6582970 . ПМИД 29891864 .
- ^ Сибли, Л.Дэвид; Хоканссон, Себастьян; Каррутерс, Верн Б. (1 января 1998 г.). «Скользящая подвижность: эффективный механизм проникновения в клетки» . Современная биология . 8 (1): С12–Р14. дои : 10.1016/S0960-9822(98)70008-9 . ПМИД 9427622 . S2CID 17555804 .
- ^ Сибли, LDI (октябрь 2010 г.). «Как апикомплексные паразиты проникают в клетки и выходят из них» . Curr Opin Biotechnol . 21 (5): 592–8. дои : 10.1016/j.copbio.2010.05.009 . ПМЦ 2947570 . ПМИД 20580218 .
- ^ Кайзер Д. (2004). «Сигнализация в миксобактериях». Ежегодный обзор микробиологии . 58 : 75–98. дои : 10.1146/annurev.micro.58.030603.123620 . ПМИД 15487930 .
- ^ Нан Б., Зусман Д.Р. (2011). «Раскрытие тайны скользящей подвижности миксобактерий» . Ежегодный обзор генетики . 45 : 21–39. doi : 10.1146/annurev-genet-110410-132547 . ПМЦ 3397683 . ПМИД 21910630 .
- ^ «Лабораторное руководство BIOL 230: Неподвижные бактерии в подвижной среде» . факультет.ccbcmd.edu . Архивировано из оригинала 15 апреля 2017 года . Проверено 8 июня 2021 г. Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Берг Х.К., Браун Д.А. (октябрь 1972 г.). «Хемотаксис Escherichia coli, анализируемый с помощью трехмерного отслеживания». Природа . 239 (5374): 500–504. Бибкод : 1972Natur.239..500B . дои : 10.1038/239500a0 . ПМИД 4563019 . S2CID 1909173 .
- ^ Сурджик В., Вингрин Н.С. (апрель 2012 г.). «Реакция на химические градиенты: бактериальный хемотаксис» . Современное мнение в области клеточной биологии . 24 (2): 262–268. дои : 10.1016/j.ceb.2011.11.008 . ПМК 3320702 . ПМИД 22169400 .
- ^ Берг, Ховард К. (1993). Случайные блуждания в биологии (Расширенная, переработанная ред.). Принстон, Нью-Джерси: Princeton Univ. Нажимать. стр. 83–94. ISBN 978-0-691-00064-0 .
- ^ Сурджик В., Вингрин Н. (апрель 2012 г.). «Реакция на химические градиенты: бактериальный хемотаксис» . Современное мнение в области клеточной биологии . 24 (2): 262–8. дои : 10.1016/j.ceb.2011.11.008 . ПМК 3320702 . ПМИД 22169400 .
- ^ Макнаб Р.М., Кошланд, Д.Э. (сентябрь 1972 г.). «Механизм восприятия градиента в бактериальном хемотаксисе» . Труды Национальной академии наук Соединенных Штатов Америки . 69 (9): 2509–12. Бибкод : 1972PNAS...69.2509M . дои : 10.1073/pnas.69.9.2509 . ПМК 426976 . ПМИД 4560688 .
- ^ Берг ХК (2003). кишечной палочки Перемещение . Нью-Йорк, штат Нью-Йорк: Спрингер. ISBN 978-0-387-00888-2 . [ нужна страница ]
- ^ Перейти обратно: а б Дуглас Р. Брамли; Франческо Каррара; Эндрю М. Хейн; Джордж И. Хагстром; Саймон А. Левин; Роман Штокер (24 июля 2020 г.). «Проходя сквозь шум: бактериальный хемотаксис в морской микросреде» . Границы морской науки . 7 . дои : 10.3389/fmars.2020.00527 . hdl : 11343/274205 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Азам, Ф. (1998). «ОКЕАНОГРАФИЯ: Микробиологический контроль океанического потока углерода: сюжет утолщается». Наука . 280 (5364): 694–696. Бибкод : 1998Sci...280..694. . дои : 10.1126/science.280.5364.694 . S2CID 83762501 .
- ^ Карсенти, Эрик; Ацинас, Сильвия Г.; Борк, Пер; Боулер, Крис; Де Варгас, Колумбан; Раес, Йерун; Салливан, Мэтью; Арендт, Детлев; Бензони, Франческа; Клавери, Жан-Мишель; Следующее, Мик; Горский, Габи; Хингамп, Паскаль; Юдикон, Дэниел; Жайон, Оливье; Кандельс-Льюис, Стефани; Кржич, Урос; Нет, Фабрис; Огата, Хироюки; Пезан, Стефан; Рейно, Эммануэль Жорж; Сардет, Кристиан; Сьераки, Майкл Э.; Речь, Сабрина; Велаюдон, Дидье; Вайсенбах, Жан; Винкер, Патрик (2011). «Целостный подход к биологии морских экосистем» . ПЛОС Биология . 9 (10): e1001177. дои : 10.1371/journal.pbio.1001177 . ПМК 3196472 . ПМИД 22028628 .
- ^ Борк, П.; Боулер, К.; Де Варгас, К.; Горский, Г.; Карсенти, Э.; Винкер, П. (2015). «Тара Оушенс изучает планктон в планетарном масштабе» . Наука . 348 (6237): 873. Бибкод : 2015Sci...348..873B . doi : 10.1126/science.aac5605 . ПМИД 25999501 . S2CID 36627341 .
- ^ Кьёрбо, Томас (2009). Механистический подход к экологии планктона . дои : 10.1515/9780691190310 . ISBN 9780691190310 .
- ^ Перейти обратно: а б Стокер, Р. (2012). «Морские микробы видят море градиентов». Наука . 338 (6107): 628–633. Бибкод : 2012Sci...338..628S . дои : 10.1126/science.1208929 . ПМИД 23118182 . S2CID 7919921 .
- ^ Хэ, Цзян, Лили; Ту, Юхай Ци ; Гуанвэй; Оуян , Чжу , ( 2012 . ) Обзорные письма . 108 (12): 128101. Бибкод : 2012PhRvL.108l8101Z . doi : 10.1103/ . PMC 3412125. . PMID 22540625 PhysRevLett.108.128101
- ^ Снеддон, штат Вашингтон; Понтий, В.; Эмонет, Т. (2012). «Стохастическая координация нескольких актуаторов уменьшает задержку и улучшает хемотаксический ответ у бактерий» . Труды Национальной академии наук . 109 (3): 805–810. Бибкод : 2012PNAS..109..805S . дои : 10.1073/pnas.1113706109 . ПМК 3271881 . ПМИД 22203971 .
- ^ Салек, М. Мехди; Каррара, Франческо; Фернандес, Висенте; Гуасто, Джеффри С.; Стокер, Роман (2019). «Бактериальный хемотаксис в микрофлюидном Т-лабиринте обнаруживает сильную фенотипическую гетерогенность в хемотаксической чувствительности» . Природные коммуникации . 10 (1): 1877. Бибкод : 2019NatCo..10.1877S . дои : 10.1038/s41467-019-09521-2 . ПМК 6478840 . ПМИД 31015402 .
- ^ Перейти обратно: а б Мартин, Э.А., изд. (1983), Словарь наук о жизни Macmillan (2-е изд.), Лондон: Macmillan Press, стр. 362, ISBN 978-0-333-34867-3
- ^ Мензель, Рэндольф (1979), «Спектральная чувствительность и цветовое зрение у беспозвоночных», в Х. Аутруме (ред.), Сравнительная физиология и эволюция зрения у беспозвоночных - A: Фоторецепторы беспозвоночных , Справочник по сенсорной физиологии, том. VII/6A, Нью-Йорк: Springer-Verlag, стр. 503–580. См. раздел D: Поведение в зависимости от длины волны и цветовое зрение, ISBN. 978-3-540-08837-0
- ^ Шарф, Биргит; Вольф, Эльмар К. (1994). «Фототаксическое поведение архебактерий Natronobacterium pharaonis». Письма ФЭБС . 340 (1–2): 114–116. дои : 10.1016/0014-5793(94)80183-5 . ПМИД 8119392 . S2CID 20435383 .
- ^ Армитидж, Джудит П.; Хеллингверф, Клаас Дж. (2003). «Свет-индуцированные поведенческие реакции (фототаксис) у прокариот». Исследования фотосинтеза . 76 (1–3): 145–155. дои : 10.1023/A:1024974111818 . ПМИД 16228574 . S2CID 9325066 .
- ^ Перейти обратно: а б с Жекели, Гаспар (2009). «Эволюция фототаксиса» . Философские труды Королевского общества B: Биологические науки . 364 (1531): 2795–2808. дои : 10.1098/rstb.2009.0072 . ПМЦ 2781859 . ПМИД 19720645 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Люке, Х.; Шоберт, Б.; Лани, Дж. К.; Спудич, Э.Н.; Спудич, Дж.Л. (2001). «Кристаллическая структура сенсорного родопсина II при 2,4 ангстрема: понимание настройки цвета и взаимодействия преобразователей» . Наука . 293 (5534): 1499–1503. Бибкод : 2001Sci...293.1499L . дои : 10.1126/science.1062977 . ПМК 4996266 . ПМИД 11452084 .
- ^ Спудич, Джон Л. (2006). «Многоталантливые микробные сенсорные родопсины». Тенденции в микробиологии . 14 (11): 480–487. дои : 10.1016/j.tim.2006.09.005 . ПМИД 17005405 .
- ^ Ян, Б.; Такахаши, Т.; Джонсон, Р.; Дергини, Ф.; Наканиши, К.; Спудич, Дж. Л. (1990). «Все-транс/13-цис-изомеризация сетчатки необходима для передачи сигналов фототаксиса сенсорными родопсинами в Halobacterium halobium» . Биофизический журнал . 57 (4): 807–814. Бибкод : 1990BpJ....57..807Y . дои : 10.1016/S0006-3495(90)82600-X . ПМЦ 1280781 . ПМИД 2344465 .
- ^ Горделий Валентин Иванович; Лабан, Йорг; Мухаметзянов Руслан; Ефремов, Руслан; Гранзин, Иоахим; Шлезингер, Рамона; Бюльдт, Георг; Савополь, Тюдор; Шайдиг, Аксель Дж.; Клер, Иоганн П.; Энгельхард, Мартин (2002). «Молекулярные основы трансмембранной передачи сигналов сенсорным комплексом родопсин II-преобразователь». Природа . 419 (6906): 484–487. Бибкод : 2002Natur.419..484G . дои : 10.1038/nature01109 . ПМИД 12368857 . S2CID 4425659 .
- ^ Сасаки, Джун; Спудич, Джон Л. (2008). «Передача сигнала в галоархейных сенсорных комплексах преобразователей родопсина» . Фотохимия и фотобиология . 84 (4): 863–868. дои : 10.1111/j.1751-1097.2008.00314.x . ПМИД 18346091 . S2CID 2811584 .
- ^ Рудольф Дж.; Остерхельт, Д. (1995). «Хемотаксис и фототаксис требуют наличия гистидинкиназы CheA у археи Halobacterium salinarium» . Журнал ЭМБО . 14 (4): 667–673. дои : 10.1002/j.1460-2075.1995.tb07045.x . ПМЦ 398130 . ПМИД 7882970 .
- ^ Маккейн, округ Колумбия; Амичи, Луизиана; Спудич, Дж. Л. (1987). «Кинетически разрешенные состояния жгутикового моторного переключателя Halobacterium halobium и модуляция переключателя сенсорным родопсином I» . Журнал бактериологии . 169 (10): 4750–4758. дои : 10.1128/jb.169.10.4750-4758.1987 . ПМК 213850 . ПМИД 3654583 .
- ^ Нульч, Вильгельм; Шухарт, Хартвиг; Хёль, Марга (1979). «Исследования фототаксической ориентации Anabaena variabilis». Архив микробиологии . 122 : 85–91. дои : 10.1007/BF00408050 . S2CID 12242837 .
- ^ Чхве, Чон-Сун; Чанг, Ён-Хо; Мун, Юн Чжон; Ким, Чанхун; Ватанабэ, Масакацу; Песня, Таблетка-Скоро; Джо, Чол-О; Богорад, Лоуренс; Пак, Ён Мок (1999). «Фотодвижение скользящей цианобактерии Synechocystis sp. PCC 6803». Фотохимия и фотобиология . 70 (1): 95–102. дои : 10.1111/j.1751-1097.1999.tb01954.x . ПМИД 10420848 . S2CID 25364218 .
- ^ Бху, Сон Хи; Дэвис, Сет Дж.; Уокер, Джозеф; Карниол, Барух; Виерстра, Ричард Д. (2001). «Бактериофитохромы представляют собой фотохромные гистидинкиназы, использующие хромофор биливердина». Природа . 414 (6865): 776–779. Бибкод : 2001Natur.414..776B . дои : 10.1038/414776a . ПМИД 11742406 . S2CID 4424642 .
- ^ Жулин, И.Б. (2000) «Новый рецептор фототаксиса, скрытый в геноме цианобактерий». Журнал молекулярной микробиологии и биотехнологии , 2 (4): 491–494.
- ^ Бхая, Деваки (2004). «Свет имеет значение: фототаксис и передача сигналов у одноклеточных цианобактерий» . Молекулярная микробиология . 53 (3): 745–754. дои : 10.1111/j.1365-2958.2004.04160.x . PMID 15255889 . S2CID 9549058 .
- ^ Ёшихара, Шизуэ; Икеучи, Масахико (2004). «Фототаксическая подвижность одноклеточной цианобактерии Synechocystis sp. PCC 6803» . Фотохимические и фотобиологические науки . 3 (6): 512–518. дои : 10.1039/b402320j . ПМИД 15170479 . S2CID 27566851 .
- ^ Ёшихара, Шизуэ; Сузуки, Фумико; Фудзита, Хиронори; Гэн, Сяо Син; Икеучи, Масахико (2000). «Новые предполагаемые фоторецепторы и регуляторные гены, необходимые для положительного фототаксического движения одноклеточной подвижной цианобактерии Synechocystis sp. PCC 6803» . Физиология растений и клеток . 41 (12): 1299–1304. дои : 10.1093/pcp/pce010 . ПМИД 11134414 .
- ^ Гествицки, Джейсон Э.; Ламанна, Эллисон К.; Харши, Расика М.; Маккартер, Линда Л.; Кисслинг, Лаура Л.; Адлер, Юлиус (2000). «Эволюционная консервация расположения метил-акцепторного белка хемотаксиса у бактерий и архей» . Журнал бактериологии . 182 (22): 6499–6502. дои : 10.1128/JB.182.22.6499-6502.2000 . ПМК 94798 . ПМИД 11053396 .
- ^ Посфаи М., Лефевр К., Трубицын Д., Базылински Д.А. и Франкель Р. (2013) «Филогенетическое значение состава и кристаллической морфологии минералов магнитосом». Границы микробиологии , 4 : 344. два : 10.3389/fmicb.2013.00344 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 3.0 .
- ^ Линь, Вэй; Чжан, Вэньси; Чжао, Сян; Робертс, Эндрю; Патерсон, Грейг; Базылински, Деннис; Пан, Юнсинь (март 2018 г.). «Геномная экспансия магнитотаксисных бактерий показывает раннее общее происхождение магнитотаксиса с эволюцией, специфичной для каждой линии» . Журнал ISME . 12 (6): 1508–1519. дои : 10.1038/s41396-018-0098-9 . ПМЦ 5955933 . ПМИД 29581530 .
- ^ Перейти обратно: а б Дюсенбери, Дэвид Б. (2009). Жизнь на микроуровне: неожиданная физика маленького размера . Кембридж, Массачусетс: Издательство Гарвардского университета. стр. 100–101. ISBN 978-0-674-03116-6 .
- ^ Перейти обратно: а б с д Ван, Кирсти Ю.; Жекели, Гаспар (2021). «Происхождение возбудимости эукариот» . Философские труды Королевского общества B: Биологические науки . 376 (1820). arXiv : 2007.13388 . дои : 10.1098/rstb.2019.0758 . ПМЦ 7935092 . ПМИД 33487111 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Якобсен, Х.Х. (2001). «Реакция бегства планктонных простейших на жидкие механические сигналы» . Серия «Прогресс в области морской экологии» . 214 : 67–78. Бибкод : 2001MEPS..214...67J . дои : 10.3354/meps214067 .
- ^ Ян, Чи-Ю; Бялецка-Форнал, Майя; Уэзервакс, Коллин; Ларкин, Джозеф В.; Приндл, Артур; Лю, Цзиньтао; Гарсия-Охальво, Хорди; Суэль, Гюрол М. (2020). «Кодирование памяти, основанной на мембранном потенциале, в микробном сообществе» . Клеточные системы . 10 (5): 417–423.e3. doi : 10.1016/j.cels.2020.04.002 . ПМЦ 7286314 . ПМИД 32343961 .
- ^ Гришанин Р.Н.; Бибиков С.И.; Альтшулер, И.М.; Каулен, AD; Казимирчук, С.Б.; Армитидж, JP; Скулачев, ВП (1996). «Дельта-пси-опосредованная передача сигналов в бактериородопсин-зависимом фотоответе» . Журнал бактериологии . 178 (11): 3008–3014. дои : 10.1128/JB.178.11.3008-3014.1996 . ПМК 178045 . ПМИД 8655473 .
- ^ Бруни, Джанкарло Н.; Уикли, Р. Эндрю; Додд, Бенджамин Дж.Т.; Краль, Джоэл М. (2017). «Потенциал-управляемый поток кальция опосредует механочувствительность Escherichia coli» . Труды Национальной академии наук . 114 (35): 9445–9450. Бибкод : 2017PNAS..114.9445B . дои : 10.1073/pnas.1703084114 . ПМЦ 5584419 . ПМИД 28808010 .
- ^ Швайницер Т., Йозенханс К. Бактериальные энергетические такси: глобальная стратегия? Арка Микробиол. Июль 2010 г.;192(7):507-20.
- ^ BNSim Техасского университета. По состоянию на 10 июня 2021 г.
- ^ Вэй, Гопенг; Богдан, Павел; Маркулеску, Раду (2013). «Эффективное моделирование и моделирование наносетей на основе бактерий с помощью BNSim». Журнал IEEE по избранным областям коммуникаций . 31 (12): 868–878. doi : 10.1109/JSAC.2013.SUP2.12130019 . S2CID 1888353 .
- ^ Перейти обратно: а б с д Курехдавуди, Хана; Богдан, Павел; Вэй, Гопенг; Маркулеску, Раду; Чжуан, Цзян; Карлсен, Рика Райт; Ситти, Метин (2017). «Мультифрактальная характеристика динамики плавания бактерий: тематическое исследование реальных и смоделированных Serratia marcescens» . Труды Королевского общества A: Математические, физические и технические науки . 473 (2203). Бибкод : 2017RSPSA.47370154K . дои : 10.1098/rspa.2017.0154 . ПМЦ 5549567 . ПМИД 28804259 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Маттеус, Франциска; Моммер, Марио С.; Курк, Тайн; Добникар, Юре (2011). «О происхождении и характеристиках индуцированных шумом прогулок Леви e. Coli» . ПЛОС ОДИН . 6 (4): e18623. Бибкод : 2011PLoSO...618623M . дои : 10.1371/journal.pone.0018623 . ПМК 3072995 . ПМИД 21494629 .
- ^ Струк, Дэниел В. (1974). «Некоторые случайные процессы, возникающие из модели движения бактерии» . Журнал теории вероятностей и смежных областей . 28 (4): 305–315. дои : 10.1007/BF00532948 . S2CID 123428558 .
- ^ Сельмечи, Дэвид; Мослер, Стефан; Хагедорн, Питер Х.; Ларсен, Нильс Б.; Фливбьерг, Хенрик (2005). «Подвижность клеток как постоянное случайное движение: теории из экспериментов» . Биофизический журнал . 89 (2): 912–931. Бибкод : 2005BpJ....89..912S . дои : 10.1529/biophysj.105.061150 . ПМЦ 1366641 . ПМИД 15951372 .
- ^ Кодлинг, Эдвард А.; Планк, Майкл Дж.; Бенаму, Саймон (2008). «Модели случайного блуждания в биологии» . Журнал интерфейса Королевского общества . 5 (25): 813–834. дои : 10.1098/rsif.2008.0014 . ПМК 2504494 . ПМИД 18426776 .
- ^ Курк, Тайн; Марендуццо, Давиде; Добникар, Юре (2013). «Хемотаксическое зондирование окружающего и секретируемого аттрактанта управляет коллективным поведением e. Coli» . ПЛОС ОДИН . 8 (10): е74878. Бибкод : 2013PLoSO...874878C . дои : 10.1371/journal.pone.0074878 . ПМЦ 3789734 . ПМИД 24098352 .
- ^ Курехдавуди, Хана; Богдан, Павел (2016). «Статистическая физическая характеристика динамики сложных систем: количественная оценка сложности на основе пространственно-временных взаимодействий» . Научные отчеты . 6 : 27602. Бибкод : 2016NatSR...627602K . дои : 10.1038/srep27602 . ПМК 4906350 . ПМИД 27297496 .
- ^ Брикар, Антуан; Коссен, Жан-Батист; Дерёмо, Николя; Дошо, Оливье; Бартоло, Денис (2013). «Появление макроскопического направленного движения в популяциях подвижных коллоидов». Природа . 503 (7474): 95–98. arXiv : 1311.2017 . Бибкод : 2013Natur.503...95B . дои : 10.1038/nature12673 . ПМИД 24201282 . S2CID 1174081 .
- ^ Отмер, Ганс Г.; Сюэ, Чуан (2013). «Математический анализ биологической агрегации и расселения: прогресс, проблемы и перспективы». В Льюисе, Марк А.; Майни, Филип К.; Петровский, Сергей В. (ред.). Расселение, индивидуальное движение и пространственная экология: математическая перспектива . Конспект лекций по математике. Том. 2071. Спрингер. стр. 79–127. дои : 10.1007/978-3-642-35497-7_4 . ISBN 978-3-642-35497-7 .
- ^ Эрбан, Радек; Хашковец, Январь (2012). «От индивидуального к коллективному поведению связанных процессов скачка скорости: пример саранчи». Кинетические и родственные модели . 5 (4): 817–842. arXiv : 1104.2584 . дои : 10.3934/krm.2012.5.817 . S2CID 18153478 .
- ^ Сюэ, Чуан; Отмер, Ханс Г. (2009). «Многомасштабные модели формирования паттернов, управляемых таксисом, в бактериальных популяциях» . SIAM Journal по прикладной математике . 70 (1): 133–167. дои : 10.1137/070711505 . ПМК 2752049 . ПМИД 19784399 .
- ^ Хиллен, Т.; Художник, К.Дж. (2009). «Руководство пользователя по моделям ФДЭ для хемотаксиса». Журнал математической биологии . 58 (1–2): 183–217. дои : 10.1007/s00285-008-0201-3 . ПМИД 18626644 . S2CID 249201 .
- ^ Эскудеро, Карлос (2006). «Дробная модель Келлера – Сигела». Нелинейность . 19 (12): 2909–2918. arXiv : math/0611496 . Бибкод : 2006Nonli..19.2909E . дои : 10.1088/0951-7715/19/12/010 . S2CID 17774620 .
- ^ Полежаев Андрей А.; Пашков Руслан А.; Лобанов Алексей Иванович; Петров, Игорь Б. (2006). «Пространственные узоры, образуемые хемотаксическими бактериями Escherichia coli» . Международный журнал биологии развития . 50 (2–3): 309–314. дои : 10.1387/ijdb.052048ap . ПМИД 16479498 .
- ^ Бейкер, Рут Э.; Йейтс, Кристиан А.; Эрбан, Радек (2010). «От микроскопического к макроскопическому описанию миграции клеток в растущих доменах» . Бюллетень математической биологии . 72 (3): 719–762. дои : 10.1007/s11538-009-9467-x . ПМИД 19862577 . S2CID 17797787 .
- ^ Шнитцер, Марк Дж. (1993). «Теория случайных блужданий в континууме и ее применение к хемотаксису». Физический обзор E . 48 (4): 2553–2568. Бибкод : 1993PhRvE..48.2553S . дои : 10.1103/PhysRevE.48.2553 . ПМИД 9960890 .
- ^ Де Женн, П.-Ж. (2004). «Хемотаксис: роль внутренних задержек». Европейский биофизический журнал . 33 (8): 691–693. дои : 10.1007/s00249-004-0426-z . ПМИД 15257424 . S2CID 412945 .
- ^ Перейти обратно: а б Кроз, Оттавио А.; Фергюсон, Гейл П.; Кейтс, Майкл Э.; Пун, Уилсон С.К. (2011). «Миграция хемотаксических бактерий в мягком агаре: роль концентрации геля» . Биофизический журнал . 101 (3): 525–534. arXiv : 1101.5063 . Бибкод : 2011BpJ...101..525C . дои : 10.1016/j.bpj.2011.06.023 . ПМК 3145277 . ПМИД 21806920 .
- ^ Чепижко, Александр; Перуани, Фернандо (2013). «Диффузия, субдиффузия и захват активных частиц в гетерогенных средах». Письма о физических отзывах . 111 (16): 160604. arXiv : 1310.0830 . Бибкод : 2013PhRvL.111p0604C . doi : 10.1103/PhysRevLett.111.160604 . ПМИД 24182247 . S2CID 11143298 .
- ^ Кейтс, Мэн (2012). «Диффузионный транспорт без детального баланса в подвижных бактериях: нужна ли микробиологии статистическая физика?». Отчеты о прогрессе в физике . 75 (4): 042601. arXiv : 1208.3957 . Бибкод : 2012РПФ...75д2601С . дои : 10.1088/0034-4885/75/4/042601 . ПМИД 22790505 . S2CID 3122097 .
- ^ Ариэль, Гил; Рабани, Амит; Бенисти, Сиван; Партридж, Джонатан Д.; Харши, Расика М.; Беер, Авраам (2015). «Роящиеся бактерии мигрируют по Леви-Уоку» . Природные коммуникации . 6 : 8396. Бибкод : 2015NatCo...6.8396A . дои : 10.1038/ncomms9396 . ПМЦ 4598630 . ПМИД 26403719 .
Внешние ссылки
[ редактировать ]- Обзор гидродинамики бактериального плавания: Лауга, Эрик (2016). «Бактериальная гидродинамика». Ежегодный обзор механики жидкости . 48 (1): 105–130. arXiv : 1509.02184 . Бибкод : 2016АнРФМ..48..105Л . doi : 10.1146/annurev-fluid-122414-034606 . S2CID 13849152 .
- Электронный учебник по бактериологии (2015 г.)
