Микробные сообщества гидротермальных источников

Микробное сообщество гидротермальных жерл включает все одноклеточные организмы, которые живут и размножаются в химически отличной области вокруг гидротермальных жерл . К ним относятся организмы в микробном мате , свободно плавающие клетки или бактерии, находящиеся в эндосимбиотических отношениях с животными. Хемолитоавтотрофные бактерии получают питательные вещества и энергию в результате геологической активности гидротермальных источников, чтобы фиксировать углерод в органических формах. Вирусы также являются частью микробного сообщества гидротермальных источников, и их влияние на микробную экологию в этих экосистемах является развивающейся областью исследований. [1]
Гидротермальные источники расположены там, где тектонические плиты расходятся и расширяются. Это позволяет воде из океана проникать в земную кору, где она нагревается магмой. Растущее давление и температура вынуждают воду выходить из этих отверстий, на выходе вода накапливает растворенные минералы и химические вещества из пород, с которыми она сталкивается. Обычно встречаются три типа жерл, и все они характеризуются температурой и химическим составом. Диффузные вентиляционные отверстия выпускают чистую воду обычно с температурой до 30 °C. Из вентиляционных отверстий белых курильщиков выделяется вода молочного цвета при температуре 200–330 °C, а из вентиляционных отверстий черных курильщиков обычно выделяется вода более горячая, чем из других вентиляционных отверстий, с температурой 300–400 °C. Воды у черных курильщиков темнеют из-за накапливающихся осадков сульфидов. [2] Из-за отсутствия солнечного света на этих глубинах океана энергия обеспечивается хемосинтезом , при котором симбиотические бактерии и археи образуют нижнюю часть пищевой цепи и способны поддерживать различные организмы, такие как Riftia pachyptila и Alvinella pompejana. Эти организмы используют эти симбиотические отношения, чтобы использовать и получать химическую энергию, которая высвобождается в этих гидротермальных жерлах. [3]
Экологические свойства
[ редактировать ]
Хотя существует большая разница температур на поверхности воды с сезонными изменениями глубины термоклина , температуры под термоклином и в водах вблизи глубокого моря относительно постоянны. Никакие изменения не вызваны сезонными эффектами или ежегодными изменениями. Эти температуры остаются в диапазоне 0–3 °C, за исключением вод, непосредственно окружающих гидротермальные источники, температура которых может достигать 407 °C. [4] [5] Эти воды не могут закипеть из-за высокого давления на этих глубинах.
С увеличением глубины начинает действовать высокое давление. Давление увеличивается примерно на 10 мегапаскалей (МПа) на каждый километр вертикального расстояния. Это значит, что на глубине траншей гидростатическое давление может достигать 110 МПа. [6]
Соленость остается относительно постоянной в глубоких морях по всему миру и составляет 35 частей на тысячу. [4]
Хотя в среде гидротермальных жерл очень мало света, были обнаружены фотосинтезирующие организмы. [7] Однако энергия, которую использует большинство организмов, поступает в результате хемосинтеза. Организмы используют минералы и химические вещества, которые выходят из жерл.
Адаптации
[ редактировать ]Экстремальные условия в среде гидротермальных жерл означают, что микробным сообществам, населяющим эти районы, необходимо адаптироваться к ним. Известно, что обитающие здесь микробы — гипертермофилы — микроорганизмы, развивающиеся при температуре выше 90 °C. Эти организмы встречаются там, где жидкости из вентиляционных отверстий выбрасываются и смешиваются с окружающей водой. Считается, что эти гипертермофильные микробы содержат белки, которые обладают повышенной стабильностью при более высоких температурах благодаря внутримолекулярным взаимодействиям, но точные механизмы пока не ясны. Механизмы стабилизации ДНК не так уж неизвестны, и считается, что денатурация ДНК сводится к минимуму за счет высоких концентраций солей, в частности Mg, K и PO4, которые высоко сконцентрированы у гипертермофилов. Наряду с этим, у многих микробов есть белки, похожие на гистоны, которые связаны с ДНК и могут обеспечивать защиту от высоких температур. Также обнаружено, что микробы находятся в симбиотических отношениях с другими организмами в среде гидротермальных жерл из-за их способности иметь механизм детоксикации, который позволяет им метаболизировать богатые сульфидами воды, которые в противном случае были бы токсичными для организмов и микробов. [8]
Микробная биогеохимия
[ редактировать ]Введение
[ редактировать ]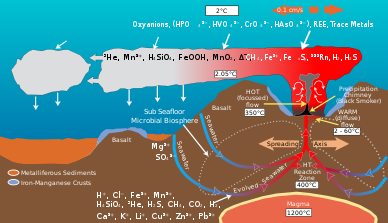
Микробные сообщества гидротермальных источников способствуют преобразованию энергии и минералов, образующихся в результате геологической деятельности, в органический материал . Органическое вещество, вырабатываемое автотрофными бактериями, затем используется для поддержания верхних трофических уровней . Гидротермальная жерла и окружающая океанская вода богаты такими элементами, как железо , марганец и различные виды серы, включая сульфиды , сульфиты , сульфаты и элементарную серу , из которых они могут получать энергию или питательные вещества. [9] Микробы получают энергию, окисляя или восстанавливая элементы. Различные виды микробов используют разные химические виды элемента в своих метаболических процессах. Например, некоторые виды микробов окисляют сульфид до сульфата, а другие виды восстанавливают сульфат до элементарной серы. В результате сеть химических путей, опосредованных различными видами микробов, преобразует такие элементы, как углерод, сера, азот и водород, из одного вида в другой. Их деятельность изменяет первоначальный химический состав, возникший в результате геологической активности среды гидротермальных жерл. [10]
Углеродный цикл
[ редактировать ]Геологическая деятельность в гидротермальных источниках приводит к обилию соединений углерода . [11] Шлейфы гидротермальных жерл содержат высокие концентрации метана и угарного газа , концентрация метана достигает 10 7 времена окружающей океанской воды. [11] [12] Глубокие океанские воды также являются большим резервуаром углерода и концентрации таких видов углекислого газа, как растворенный CO 2 и HCO 3. − около 2,2 мм. [13] Обильные акцепторы углерода и электронов, образующиеся в результате геологической деятельности, поддерживают оазис хемоавтотрофных микробных сообществ, которые фиксируют неорганический углерод, такой как CO 2 , используя энергию из таких источников, как окисление серы, железа, марганца, водорода и метана. [11] Эти бактерии поставляют большую часть органического углерода, который поддерживает гетеротрофную жизнь в гидротермальных источниках. [14]
Фиксация углерода
[ редактировать ]Фиксация углерода – это включение неорганического углерода в органическое вещество. В отличие от поверхности планеты, где свет является основным источником энергии для фиксации углерода, хемолитотрофные бактерии гидротермальных источников полагаются на химическое окисление для получения необходимой энергии. [15] Фиксация CO 2 наблюдается у представителей Gammaproteobacteria , Campylobacterota , Alphaproteobacteria и представителей домена Archaea в гидротермальных источниках. Четыре основных метаболических пути фиксации углерода, обнаруженные в микробных жерловых сообществах, включают цикл Кальвина-Бенсона-Бэсшама (CBB), цикл восстановительных трикарбоновых кислот (rTCA), цикл 3-гидроксипропионата (3-HP) и восстановительный ацетилкоэнзим A (ацетил-КоА). ) путь. [15]
Метаболические пути фиксации углерода
[ редактировать ]Цикл Кальвина-Бенсона-Бэсшема (CBB)
[ редактировать ]Цикл Кальвина-Бенсона-Башама (CBB) является наиболее распространенным путем фиксации CO2, обнаруженным среди автотрофов. [16] Ключевым ферментом является рибулозо-1,5-бисфосфаткарбоксилаза/оксигеназа ( RuBisCO ). [15] RuBisCO был идентифицирован у членов микробного сообщества, таких как Thiomicrospira, Beggiatoa , zetaproteobacterium и гаммапротеобактериальных эндосимбионтов трубчатых червей , двустворчатых моллюсков и брюхоногих моллюсков . [16]
Восстановительный цикл карбоновых кислот (rTCA)
[ редактировать ]Восстановительный цикл карбоновых кислот (rTCA) является вторым наиболее часто встречающимся путем фиксации углерода в гидротермальных источниках. [16] Цикл rTCA, по сути, представляет собой обращенный цикл TCA или цикл Кребса, который гетеротрофы используют для окисления органических веществ. Организмы, использующие цикл rTCA, предпочитают обитать в бескислородных зонах гидротермальной жерловой системы, поскольку некоторые ферменты цикла rTCA чувствительны к присутствию O 2 . [15] Он обнаружен в сульфатредуцирующих дельтапротеобактериях, таких как некоторые представители Desulfobacter , Aquificales , Aquifex и Thermoproteales . [15]
Циклы 3-HP и 3-HP/4-HB
[ редактировать ]Ключевыми ферментами циклов 3-HP и 3-HP/4-HB являются ацетил-КоА/пропионил-КоА-карбоксилаза, малонил-КоА-редуктаза и пропионил-КоА-синтаза. Большинство организмов, использующих этот путь, являются миксотрофами , способными использовать органический углерод в дополнение к фиксации углерода. [15]
Восстановительный путь ацетил-КоА
[ редактировать ]Путь восстановительного ацетил-КоА обнаружен только у хемоавтотрофов. Этот путь не требует АТФ , поскольку этот путь напрямую связан с восстановлением H 2 . Организмы, обнаруженные этим путем, предпочитают районы, богатые H 2 . Виды включают дельтапротеобактерии, такие как Dulfobacterium autotropicum , ацетогены и метаногенные археи . [15]
Метановый обмен
[ редактировать ]Гидротермальные источники производят большое количество метана , который может возникать как в результате геологических, так и биологических процессов. [11] [12] Концентрация метана в шлейфах гидротермальных жерл может превышать 300 мкм в зависимости от жерла. Для сравнения, вентиляционная жидкость содержит 10 6 – 10 7 раз больше метана, чем окружающая глубинная океанская вода, концентрация метана в которой колеблется от 0,2 до 0,3 нМ. [12] Микробные сообщества используют высокие концентрации метана в качестве источника энергии и источника углерода. [11] Метанотрофия , при которой вид использует метан как в качестве энергии, так и в качестве источника углерода, наблюдалась при наличии гаммапротеобактерий в линиях Mmethylococcaceae . [16] Метанотрофы преобразуют метан в углекислый газ и органический углерод. [12] Для них обычно характерно наличие межклеточных мембран, и было замечено, что микробы с межклеточными мембранами составляют 20% микробного мата в гидротермальных источниках. [11] [12]
Окисление метана
[ редактировать ]Производство энергии посредством окисления метана является следующим лучшим источником энергии после окисления серы. [11] Было высказано предположение, что микробное окисление способствует быстрому круговороту в гидротермальных источниках, поэтому большая часть метана окисляется на небольшом расстоянии от источника. [12] В сообществах гидротермальных жерл аэробное окисление метана обычно обнаруживается у эндосимбиотических микробов жерловых животных. [17] Анаэробное окисление метана (АОМ) обычно сочетается с восстановлением сульфата или Fe и Mn как терминальных акцепторов электронов , поскольку их больше всего в гидротермальных источниках. [12] [18] Обнаружено, что АОМ преобладает в морских отложениях гидротермальных источников. [19] [18] и может быть ответственным за потребление 75% метана, вырабатываемого источником. [18] Виды, которые выполняют АОМ, включают Archaea типа Thermoproteota (ранее Crenarchaeota ) и Thermococcus . [20]
Метаногенез
[ редактировать ]Производство метана посредством метаногенеза может происходить в результате разложения углеводородов , реакции углекислого газа или других соединений, таких как формиат . [17] Свидетельства метаногенеза можно найти рядом с АОМ в отложениях. [19] термофильные метаногены растут в шлейфах гидротермальных жерл при температуре от 55 ° C до 80 ° C. Обнаружено, что [21] Однако автотропный метаногенез, осуществляемый многими термофильными видами, требует H 2 в качестве донора электронов, поэтому рост микробов ограничивается доступностью H 2 . [21] [14] Роды термофильных метаногенов, обнаруженных в гидротермальных источниках, включают Methanocaldococcus , Methanothermococcus и Methanococcus . [21]

Цикл серы
[ редактировать ]Микробные сообщества гидротермальных источников преобразуют серу, такую как H 2 S, образующуюся в результате геологической деятельности, в другие формы, такие как сульфит , сульфат и элементарную серу , для получения энергии или ассимиляции в органические молекулы . [10] Сульфидов много в гидротермальных источниках, их концентрации от одного до десятков мМ, тогда как окружающая океанская вода обычно содержит лишь несколько наномолей. [22]
Окисление серы
[ редактировать ]Восстановленные соединения серы, такие как H 2 S, образующиеся в гидротермальных источниках, являются основным источником энергии для метаболизма серы у микробов. [11] Окисление восстановленных соединений серы в такие формы, как сульфит , тиосульфат и элементарная сера, используется для производства энергии для микробного метаболизма, например, для синтеза органических соединений из неорганического углерода . [10] Основные метаболические пути, используемые для окисления серы, включают SOX путь и диссимиляционное окисление. Путь Sox представляет собой мультиферментный путь, способный окислять сульфид, сульфит, элементарную серу и тиосульфат до сульфата. [10] Диссимиляционное окисление превращает сульфит в элементарную серу. [9] сероокисляющим видам относятся роды Thiomicrospira , Halothiobacillus , Beggiatoa , Persephonella и Sulfurimonas . К Симбиотические виды класса Gammaproteobacteria и филмы Campylobacterota также могут окислять серу. [10]
Снижение содержания серы
[ редактировать ]При восстановлении серы сульфат используется в качестве акцептора электронов для ассимиляции серы . Микробы, выполняющие сульфатредукцию, обычно используют водород , метан или органические вещества в качестве донора электронов . [18] [23] При анаэробном окислении метана (АОМ) в качестве акцептора электронов часто используют сульфат. [18] Этот метод предпочитают организмы, живущие в сильно бескислородных районах гидротермальных источников. [23] таким образом, это один из преобладающих процессов, происходящих в отложениях. [14] Виды, восстанавливающие сульфат, были идентифицированы у архей и представителей дельтапротеобактерий, таких как Desulfovibrio , Desulfobulbus , Desulfobacteria и Desulfuromonas , в гидротермальных источниках. [23]
Азотный цикл
[ редактировать ]Глубокие океанские воды содержат самый большой резервуар азота, доступный для гидротермальных источников, с содержанием растворенного газообразного азота около 0,59 ммоль. [24] [25] Аммоний является доминирующим видом растворенного неорганического азота и может производиться путем смешивания водных масс под гидротермальными жерлами и сброса их в жерловые жидкости. [25] Количество доступного аммония варьируется в зависимости от жерл в зависимости от геологической активности и микробного состава. [25] Концентрация нитратов и нитритов в гидротермальных источниках снижена по сравнению с окружающей морской водой. [24]
Изучение цикла азота в микробных сообществах гидротермальных источников все еще требует более комплексных исследований. [24] Однако изотопные данные показывают, что микроорганизмы влияют на количество и состав растворенного неорганического азота, и все пути азотного цикла, вероятно, могут быть обнаружены в гидротермальных источниках. [25] Биологическая фиксация азота важна для обеспечения части биологически доступного азота в азотном цикле, особенно в неосадочных гидротермальных источниках. [24] Азот фиксируется множеством различных микробов, включая метаноген из отрядов Methanomirobiales , Methanococcales и Methanobacteriales . [24] Было обнаружено, что термофильные микробы способны фиксировать азот при более высоких температурах, например 92 ° C. [24] Фиксация азота может быть особенно распространена в микробных матах и сыпучем материале , где биологически доступные уровни азота низки из-за высокой плотности микробов и анаэробной среды, которая обеспечивает функцию нитрогеназы , азотфиксирующего фермента. [24] Также были обнаружены доказательства ассимиляции , нитрификации , денитрификации , анаммокса , минерализации и диссимиляционного восстановления нитратов до аммония . [25] Например, сероокисляющие бактерии, такие как виды Begiatoa, выполняют денитрификацию и восстанавливают нитраты до окисления H 2 S. [24] Ассимиляцию нитратов осуществляют симбиотические виды трубчатого червя Riftia pachyptila . [24]
Бактериальное разнообразие
[ редактировать ]Наиболее распространенными бактериями гидротермальных источников являются хемолитотрофы. Эти бактерии используют восстановленные химические соединения, чаще всего серу, в качестве источников энергии для восстановления углекислого газа до органического углерода. [11] Численность хемолитотрофов в среде гидротермальных жерл определяется доступными источниками энергии; В жерлах с разной температурой концентрация питательных веществ разная, что предполагает большие различия между жерлами. В целом, большие микробные популяции обнаруживаются в шлейфах теплых жерловых вод (25 °C), на поверхностях, подвергающихся воздействию теплых жерловых шлейфов, а также в симбиотических тканях некоторых жерловых беспозвоночных вблизи жерловых жерл. [11]
Сероокисляющий
[ редактировать ]Эти бактерии используют различные формы доступной серы (S −2 , С 0 , OS2O3 −2 ) в присутствии кислорода. Они составляют преобладающую популяцию в большинстве гидротермальных источников, поскольку их источник энергии широко доступен, а скорость хемосинтеза увеличивается в аэробных условиях. Бактерии гидротермальных источников аналогичны типам серобактерий, обитающих в других что средах, богатых сероводородом, за исключением того, Thiomicrospira на смену Thiobacillus пришла . Другими распространенными видами являются Thiothrix и Beggiatoa, которые имеют особое значение из-за своей способности фиксировать азот. [11]
Метанокисляющий
[ редактировать ]Метан является существенным источником энергии в некоторых гидротермальных источниках, но не в других: в теплых источниках (25 ° C) метана больше, чем водорода. [11] Существует множество типов метанотрофных бактерий, которые нуждаются в кислороде и фиксируют CH 4 , CH 3 NH 2 и другие соединения C 1 , включая CO 2 и CO, если они присутствуют в жерловой воде. [11] Бактерии этого типа также обнаружены в трофосоме Riftia , что указывает на симбиотические отношения. [11] Здесь метанокисляющие бактерии относятся к метанотрофам , которые не тождественны метаногенам : Methanococcus и Methanocaldococcus jannaschii являются примерами метаногенов, [11] которые встречаются в гидротермальных источниках; тогда как Mmethylocystaceae являются метанотрофами, которые также были обнаружены в сообществах гидротермальных жерл. [26]
Водородокисляющий
[ редактировать ]О микробах, использующих водород в качестве источника энергии, известно немного, однако исследования показали, что они аэробны, а также симбиотичны с рифтией (см. ниже). [11] [27] полученный геотермальным путем . H 2 , Эти бактерии играют важную роль в первичном производстве органического углерода, поскольку в этом процессе используется [11] В жерлах, где NO 3 − -содержащая придонная морская вода смешивается с гидротермальным флюидом. [11] Desulfonauticus submarinus — гидрогенотроф , восстанавливающий соединения серы в теплых источниках; он был обнаружен у трубочных червей R. pachyptila и Alvinella pompejana. [28]
Окисляющие железо и марганец
[ редактировать ]Эти бактерии обычно встречаются в отложениях железа и марганца на поверхностях, периодически подвергающихся воздействию шлейфов гидротермальной и придонной морской воды. Однако из-за быстрого окисления Fe 2+ В нейтральных и щелочных водах (т.е. в пресной и морской воде) бактерии, ответственные за окислительное отложение железа, чаще встречаются в кислых водах. [11] Бактерии, окисляющие марганец, будут более распространены в пресной и морской воде по сравнению с бактериями, окисляющими железо, из-за более высокой концентрации доступного металла. [11]
Экология
[ редактировать ]Симбиотические отношения
[ редактировать ]Симбиотический хемосинтез - важный процесс для гидротермальных жерловых сообществ. [11] В теплых жерлах обычными симбионтами бактерий являются глубоководные моллюски Calpytogena Magnifica , мидии, такие как Bathyomodiolus thermophilus , и погонофоровые трубчатые черви, Riftia pachyptila и Alvinella pompejana . [11] [27] [ самостоятельно опубликованный источник? ] [28] Трофосомы . этих животных являются особыми органами для симбионтов, содержащими ценные молекулы для хемосинтеза Эти организмы стали настолько зависимы от своих симбионтов, что утратили все морфологические особенности, связанные с пищеварением и пищеварением, хотя бактерии обеспечиваются H 2 S и свободным O 2 . [11] были выделены метанокисляющие бактерии Кроме того, из C. Magnifica и R. pachyptila , что указывает на то, что ассимиляция метана может происходить внутри трофосомы этих организмов. [9]
Филы и роды
[ редактировать ]Чтобы проиллюстрировать невероятное разнообразие гидротермальных источников, приведенный ниже список представляет собой совокупное представление типов и родов бактерий в алфавитном порядке. Как показано, протеобактерии, по-видимому, являются наиболее доминирующим типом, присутствующим в глубоководных жерлах.
- Актиномицетота [16]
- Аквификота
- Хлорофлексота [16]
- Хлоробиота
- Деферрибактериота
- Одаренный драгоценными камнями [16]
- Нитроспирота
- Нитроспинота [16]
- Лептоспириллум феррифилум
- Бациллота
- Ацетоген: клостридии [16]
- Псевдомонадота
- Кампилобактерия [11] [15] [16]
- Термодесульфобактерии – сульфатредуцирующие, составляют более 25% бактериального сообщества. [15] [16] [23] [11]
восстановление ДНК
[ редактировать ]Микробные сообщества, населяющие глубоководные гидротермальные жерла, по-видимому, сильно обогащены генами, которые кодируют ферменты, используемые в репарации несоответствий ДНК и гомологичной рекомбинации . [29] Это открытие предполагает, что эти микробные сообщества развили обширные возможности восстановления ДНК , чтобы справляться с экстремально повреждающими ДНК условиями , в которых они существуют. [29]
Вирусы и глубоководные гидротермальные источники
[ редактировать ]
Вирусы — самая распространенная форма жизни в океане, таящая в себе величайший резервуар генетического разнообразия. [30] Поскольку их инфекции часто заканчиваются смертельным исходом, они представляют собой значительный источник смертности и, таким образом, оказывают широкое влияние на биологические океанографические процессы, эволюцию и биогеохимический цикл в океане. [31] Однако были обнаружены доказательства того, что вирусы, обнаруженные в жерловых средах обитания, приняли более мутуалистическую , чем паразитическую эволюционную стратегию, чтобы выжить в экстремальной и нестабильной среде, в которой они существуют. [32]
Было обнаружено, что глубоководные гидротермальные источники содержат большое количество вирусов, что указывает на высокую продукцию вирусов. [33] Образцы из гидротермальных источников Индевор у юго-западного побережья Британской Колумбии показали, что у активных чернокожих курильщиков содержание вируса составляло от 1,45×10. 5 до 9,90×10 7 на мл, с падением численности, обнаруженным в шлейфе гидротермальных источников (3,5 × 10 6 на мл) и вне системы вентиляции (2,94×10 6 за мл). Высокая плотность вирусов и, следовательно, их продукции (по сравнению с окружающими глубоководными водами) означает, что вирусы являются значительным источником микробной смертности в жерловых источниках. [33] Как и в других морских средах, глубоководные гидротермальные вирусы влияют на численность и разнообразие прокариот и, следовательно, влияют на микробный биогеохимический цикл, лизис своих хозяев для репликации. [34]
Однако, в отличие от своей роли источника смертности и контроля численности населения, предполагается, что вирусы также повышают выживаемость прокариот в экстремальных условиях, выступая в качестве резервуаров генетической информации. Поэтому считается, что взаимодействие виросферы с микроорганизмами в условиях стресса окружающей среды способствует выживанию микроорганизмов за счет распространения генов хозяина посредством горизонтального переноса генов . [35]
Каждую секунду « примерно такое же количество инфекций, как у Авогадро в океане происходит , и каждое из этих взаимодействий может привести к передаче генетической информации между вирусом и хозяином». — Кертис Саттл [36]
Умеренные фаги (не вызывающие немедленного лизиса) иногда могут придавать фенотипы , улучшающие приспособленность прокариот. [7] Лизогенный лизогенная жизненный цикл может стабильно сохраняться на протяжении тысяч поколений инфицированных бактерий, а вирусы могут изменять фенотип хозяина, активируя гены (процесс, известный как конверсия ), что, следовательно, может позволить хозяину справляться с различными средами. [37] Польза для популяции-хозяина также может быть получена за счет экспрессии фенотипов, кодируемых фагами и улучшающих приспособленность. [38]

Обзор вирусных исследований в гидротермальных жерлах, опубликованный в 2015 году, показал, что жерла содержат значительную часть лизогенных хозяев и что большая часть вирусов являются умеренными, что указывает на то, что жерла могут дать преимущество профагу. [39]
Одно исследование взаимодействий вирус-хозяин в среде гидротермальных жерл с диффузным потоком показало, что высокая заболеваемость лизогенными хозяевами и большие популяции вирусов умеренного пояса уникальны по своим масштабам и что эти вирусы, вероятно, имеют решающее значение для системной экологии прокариот. Генетический анализ того же исследования показал, что 51% последовательностей вирусного метагенома были неизвестны (отсутствовали гомологии с данными секвенирования), с высоким разнообразием в жерловых средах, но с меньшим разнообразием для конкретных жерловых участков, что указывает на высокую специфичность для вирусных мишеней. [38]
глубоководных гидротермальных источников Метагеномный анализ виромов показал, что вирусные гены манипулируют метаболизмом бактерий , участвуя в метаболических путях, а также образуя разветвленные пути микробного метаболизма, которые облегчают адаптацию к экстремальным условиям окружающей среды. [40]
Пример этого был связан с бактерией SUP05, потребляющей серу. Исследование показало, что 15 из 18 вирусных геномов, секвенированных из образцов жерловых шлейфов, содержали гены, тесно связанные с ферментом, который хемолитоавтотрофы SUP05 используют для извлечения энергии из соединений серы. Авторы пришли к выводу, что такие фаговые гены ( вспомогательные метаболические гены ), которые способны усиливать метаболизм окисления серы в своих хозяевах, могут обеспечивать вирусам селективные преимущества (продолжающееся заражение и репликация). [41] Сходство вирусных генов и генов SUP05, отвечающих за метаболизм серы, предполагает обмен генами в прошлом и может указывать на то, что вирусы являются агентами эволюции. [42]
Другое метагеномное исследование показало, что вирусные гены имеют относительно высокую долю генов метаболизма, витаминов и кофакторов , что указывает на то, что вирусные геномы кодируют вспомогательные метаболические гены. В сочетании с наблюдениями за высокой долей лизогенных вирусов это указывает на то, что вирусы отбираются в качестве интегрированных провирусов, а не свободно плавающих вирусов, и что вспомогательные гены могут экспрессироваться, принося пользу как хозяину, так и интегрированному вирусу. Вирусы улучшают физическую форму, ускоряя метаболизм или предлагая большую метаболическую гибкость своим хозяевам. Имеющиеся данные свидетельствуют о том, что вирусные эволюционные стратегии глубоководных гидротермальных жерл способствуют длительной интеграции хозяина, отдавая предпочтение форме мутуализма, а не классическому паразитизму. [32]
Как выходы гидротермальных источников [ нужны разъяснения ] Что касается материала под морским дном, то, вероятно, также существует связь между жерловыми вирусами и вирусами в земной коре. [39]
См. также
[ редактировать ]- Морской микроорганизм
- Пещерные курганы
- Бассейн Гуаймас
- Хемосинтез сероводорода - система получения энергии, используемая в гидротермальных источниках.
Ссылки
[ редактировать ]- ^ Андерсон Р.Э., Бразелтон В.Дж., Баросс Дж.А. (2011). «Затрагивается ли вирусами генетический ландшафт глубокой подповерхностной биосферы?» . Границы микробиологии . 2 : 219. дои : 10.3389/fmicb.2011.00219 . ПМК 3211056 . ПМИД 22084639 .
- ^ Лутц Р.А., Кенниш М.Дж. (август 1993 г.). «Экология глубоководных гидротермальных жерл: обзор». Обзоры геофизики . 31 (3): 211–242. Бибкод : 1993RvGeo..31..211L . дои : 10.1029/93rg01280 .
- ^ Кадар Э., Коста В., Сантос Р.С., Пауэлл Дж.Дж. (июль 2006 г.). «Тканевое распределение микроэссенциальных металлов в жерловых двустворчатых моллюсках Bathymodiolus azoricus и связанных с ними организмах (эндосимбионтных бактериях и паразитических полихетах) из геохимически различных жерл Срединно-Атлантического хребта». Журнал морских исследований . 56 (1): 45–52. Бибкод : 2006JSR....56...45K . дои : 10.1016/j.seares.2006.01.002 .
- ^ Jump up to: а б Бергман Дж. (16 февраля 2011 г.). «Температура океанской воды» . Окна во Вселенную .
- ^ Хаазе К.М., Петерсен С., Кощинский А., Зейферт Р., Деви К.В., Кейр Р. и др. (ноябрь 2007 г.). «Молодой вулканизм и связанная с ним гидротермальная активность на 5 ° ю.ш. на медленно расширяющемся южном Срединно-Атлантическом хребте» (PDF) . Геохимия, геофизика, геосистемы . 8 (11): н/д. Бибкод : 2007GGG.....811002H . дои : 10.1029/2006gc001509 .
- ^ Джеббар М., Францетти Б., Жирар Э., Огер П. (июль 2015 г.). «Микробное разнообразие и адаптация к высокому гидростатическому давлению у прокариот глубоководных гидротермальных источников». Экстремофилы . 19 (4): 721–740. дои : 10.1007/s00792-015-0760-3 . ПМИД 26101015 . S2CID 17213654 .
- ^ Битти Дж.Т., Оверманн Дж., Линс М.Т., Манске А.К., Ланг А.С., Бланкеншип Р.Э. и др. (июнь 2005 г.). «Облигатно фотосинтезирующий бактериальный анаэроб из глубоководных гидротермальных источников» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (26): 9306–9310. Бибкод : 2005PNAS..102.9306B . дои : 10.1073/pnas.0503674102 . ПМК 1166624 . ПМИД 15967984 .
- ^ Зиренберг Р.А., Адамс М.В., Арп А.Дж. (ноябрь 2000 г.). «Жизнь в экстремальных условиях: гидротермальные источники» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (24): 12961–12962. Бибкод : 2000PNAS...9712961Z . дои : 10.1073/pnas.210395997 . ПМК 34077 . ПМИД 11058150 .
- ^ Jump up to: а б с Клецин А., Урих Т., Мюллер Ф., Бандейрас Т.М., Гомес С.М. (февраль 2004 г.). «Диссимиляционное окисление и восстановление элементарной серы у термофильных архей». Журнал биоэнергетики и биомембран . 36 (1): 77–91. doi : 10.1023/b:jobb.0000019600.36757.8c . ПМИД 15168612 . S2CID 45653369 .
- ^ Jump up to: а б с д и Зиверт С.М., Хюглер М., Тейлор К.Д., Вирсен К.О. (2008). «Окисление серы в глубоководных гидротермальных источниках». Микробный метаболизм серы . стр. 238–258. дои : 10.1007/978-3-540-72682-1_19 . ISBN 978-3-540-72679-1 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С Яннаш Х.В., Моттл М.Дж. (август 1985 г.). «Геомикробиология глубоководных гидротермальных источников». Наука . 229 (4715): 717–725. Бибкод : 1985Sci...229..717J . дои : 10.1126/science.229.4715.717 . JSTOR 1696097 . ПМИД 17841485 . S2CID 24859537 .
- ^ Jump up to: а б с д и ж г де Анжелис, Массачусетс (1989). Исследования микробного окисления метана в глубоководных гидротермальных жерлах (Диссертация). ПроКвест 303750552 .
- ^ Дунк Р.М., Пельтцер Э.Т., Уолц П.М., Брюэр П.Г. (декабрь 2005 г.). «Взгляд на эксперимент по обогащению CO2 в глубоком океане в новом свете: лазерное рамановское обнаружение растворенного CO2 в морской воде». Экологические науки и технологии . 39 (24): 9630–9636. Бибкод : 2005EnST...39.9630D . дои : 10.1021/es0511725 . ПМИД 16475344 .
- ^ Jump up to: а б с Вер Ике Х.К., Баттерфилд Д.А., Хубер Дж.А., Лилли М.Д., Олсон Э.Дж., Роу К.К. и др. (август 2012 г.). «Ограниченный водородом рост гипертермофильных метаногенов в глубоководных гидротермальных источниках» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (34): 13674–13679. Бибкод : 2012PNAS..10913674V . дои : 10.1073/pnas.1206632109 . ПМК 3427048 . ПМИД 22869718 .
- ^ Jump up to: а б с д и ж г час я дж к л Накагава С., Такаи К. (июль 2008 г.). «Хемоавтотрофы глубоководных жерл: разнообразие, биохимия и экологическое значение» . ФЭМС Микробиология Экология . 65 (1): 1–14. дои : 10.1111/j.1574-6941.2008.00502.x . ПМИД 18503548 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Серкейра Т., Баррозу К., Фруф Х., Эгаш К., Бетанкур Р. (август 2018 г.). «Метагеномные признаки микробных сообществ в глубоководных гидротермальных отложениях жерловых полей Азорских островов». Микробная экология . 76 (2): 387–403. дои : 10.1007/s00248-018-1144-x . ПМИД 29354879 . S2CID 7879639 .
- ^ Jump up to: а б Мартин В., Баросс Дж., Келли Д., Рассел М.Дж. (ноябрь 2008 г.). «Гидротермальные источники и происхождение жизни». Обзоры природы. Микробиология . 6 (11): 805–814. дои : 10.1038/nrmicro1991 . ПМИД 18820700 . S2CID 1709272 .
- ^ Jump up to: а б с д и Ванкель С.Д., Адамс М.М., Джонстон Д.Т., Гензель С.М., Джой С.Б., Гиргис П.Р. (октябрь 2012 г.). «Анаэробное окисление метана в металлоносных гидротермальных отложениях: влияние на поток углерода и отделение от сульфатредукции». Экологическая микробиология . 14 (10): 2726–2740. дои : 10.1111/j.1462-2920.2012.02825.x . ПМИД 22827909 .
- ^ Jump up to: а б Биддл Дж. Ф., Кардман З., Мендловиц Х., Альберт Д.Б., Ллойд К.Г., Боэций А., Теске А. (май 2012 г.). «Анаэробное окисление метана при различных температурных режимах в гидротермальных отложениях бассейна Гуаймас» . Журнал ISME . 6 (5): 1018–1031. дои : 10.1038/ismej.2011.164 . ПМК 3329104 . ПМИД 22094346 .
- ^ Бразелтон В.Дж., Шренк М.О., Келли Д.С., Баросс Дж.А. (сентябрь 2006 г.). «Микробные сообщества, метаболизирующие метан и серу, доминируют в экосистеме гидротермального поля Затерянного города» . Прикладная и экологическая микробиология . 72 (9): 6257–6270. Бибкод : 2006ApEnM..72.6257B . дои : 10.1128/АЕМ.00574-06 . ПМЦ 1563643 . ПМИД 16957253 .
- ^ Jump up to: а б с Топчуоглу Б.Д., Стюарт Л.К., Моррисон Х.Г., Баттерфилд Д.А., Хубер Дж.А., Холден Дж.Ф. (5 августа 2016 г.). «Ограничение водорода и синтрофный рост среди природных комплексов термофильных метаногенов в глубоководных гидротермальных источниках» . Границы микробиологии . 7 : 1240. дои : 10.3389/fmicb.2016.01240 . ПМЦ 4974244 . ПМИД 27547206 .
- ^ Рэдфорд-Кноери Дж., Герман Ч.Р., Шарлу Дж.Л., Донваль Дж.П., Фуке Ю. (март 2001 г.). «Распределение и поведение растворенного сероводорода в гидротермальных шлейфах» . Лимнология и океанография . 46 (2): 461–464. Бибкод : 2001LimOc..46..461R . дои : 10.4319/lo.2001.46.2.0461 .
- ^ Jump up to: а б с д Фрэнк К.Л., Роджерс Д.Р., Олинс Х.К., Видудес К., Гиргис П.Р. (июль 2013 г.). «Характеристика распределения и скорости микробной сульфатредукции в гидротермальных источниках Средней долины» . Журнал ISME . 7 (7): 1391–1401. дои : 10.1038/ismej.2013.17 . ПМК 3695286 . ПМИД 23535916 .
- ^ Jump up to: а б с д и ж г час я Мехта, член парламента, Баттерфилд, Д.А., Баросс Дж.А. (февраль 2003 г.). «Филогенетическое разнообразие генов нитрогеназы (nifH) в глубоководных и гидротермальных жерлах хребта Хуан-де-Фука» . Прикладная и экологическая микробиология . 69 (2): 960–970. Бибкод : 2003ApEnM..69..960M . doi : 10.1128/aem.69.2.960-970.2003 . ПМК 143675 . ПМИД 12571018 .
- ^ Jump up to: а б с д и Бурбонне А., Леманн М.Ф., Баттерфилд Д.А., Джунипер С.К. (февраль 2012 г.). «Преобразования подводного азота в диффузных гидротермальных жерловых флюидах хребта Хуан-де-Фука, о чем свидетельствует изотопный состав нитратов и аммония» . Геохимия, геофизика, геосистемы . 13 (2): н/д. Бибкод : 2012GGG....13.2T01B . дои : 10.1029/2011gc003863 .
- ^ Скеннертон К.Т., Уорд Л.М., Мишель А., Меткалф К., Валиенте С., Маллин С. и др. (23 декабря 2015 г.). «Геномная реконструкция некультивируемого гидротермального источника гаммапротеобактериального метанотрофа (семейство Mmethylothermaceae) указывает на множественную адаптацию к ограничению кислорода» . Границы микробиологии . 6 : 1425. дои : 10.3389/fmicb.2015.01425 . ПМЦ 4688376 . ПМИД 26779119 .
- ^ Jump up to: а б «Гидротермальные источники — микробевики» . microbewiki.kenyon.edu . Проверено 22 октября 2018 г.
- ^ Jump up to: а б Одиффрин С., Кайол Дж.Л., Жулиан С., Касалот Л., Томас П., Гарсия Дж.Л., Оливье Б. (сентябрь 2003 г.). «Desulfonauticus submarinus gen. nov., sp. nov., новая сульфатредуцирующая бактерия, выделенная из глубоководных гидротермальных источников» . Международный журнал систематической и эволюционной микробиологии . 53 (Часть 5): 1585–1590. дои : 10.1099/ijs.0.02551-0 . ПМИД 13130052 .
- ^ Jump up to: а б Се В., Ван Ф., Го Л., Чен З., Зиверт С.М., Мэн Дж. и др. (март 2011 г.). «Сравнительная метагеномика микробных сообществ, населяющих глубоководные гидротермальные жерла с контрастным химическим составом» . Журнал ISME . 5 (3): 414–426. дои : 10.1038/ismej.2010.144 . ПМК 3105715 . ПМИД 20927138 .
- ^ Саттл, Калифорния (сентябрь 2005 г.). «Вирусы в море». Природа . 437 (7057): 356–361. Бибкод : 2005Natur.437..356S . дои : 10.1038/nature04160 . ПМИД 16163346 . S2CID 4370363 .
- ^ Саттл, Калифорния (октябрь 2007 г.). «Морские вирусы — основные игроки глобальной экосистемы». Обзоры природы. Микробиология . 5 (10): 801–812. дои : 10.1038/nrmicro1750 . ПМИД 17853907 . S2CID 4658457 .
- ^ Jump up to: а б Андерсон Р.Э., Согин М.Л., Баросс Дж.А. (3 октября 2014 г.). «Эволюционные стратегии вирусов, бактерий и архей в экосистемах гидротермальных жерл, выявленные с помощью метагеномики» . ПЛОС ОДИН . 9 (10): e109696. Бибкод : 2014PLoSO...9j9696A . дои : 10.1371/journal.pone.0109696 . ПМЦ 4184897 . ПМИД 25279954 .
- ^ Jump up to: а б Ортманн AC, Саттл, Калифорния (август 2005 г.). «Высокое содержание вирусов в глубоководной гидротермальной системе указывает на вирусно-опосредованную микробную смертность». Глубоководные исследования. Часть I: Статьи океанографических исследований . 52 (8): 1515–1527. Бибкод : 2005DSRI...52.1515O . дои : 10.1016/j.dsr.2005.04.002 .
- ^ Брейтбарт М (15 января 2012 г.). «Морские вирусы: правда или действие». Ежегодный обзор морской науки . 4 (1): 425–448. Бибкод : 2012ARMS....4..425B . doi : 10.1146/annurev-marine-120709-142805 . ПМИД 22457982 .
- ^ Гольденфельд Н., Везе С. (январь 2007 г.). «Следующая революция биологии». Природа . 445 (7126): 369. arXiv : q-bio/0702015 . Бибкод : 2007Natur.445..369G . дои : 10.1038/445369а . ПМИД 17251963 . S2CID 10737747 .
- ^ Каллауэй Э (21 августа 2008 г.). «Новый вирусный образ жизни обнаружен в глубоководных жерлах» . Новый учёный .
- ^ Клоки М.Р., Миллард А.Д., Летаров А.В., Хифи С. (январь 2011 г.). «Фаги в природе» . Бактериофаг . 1 (1): 31–45. дои : 10.4161/bact.1.1.14942 . ПМК 3109452 . ПМИД 21687533 .
- ^ Jump up to: а б Уильямсон С.Дж., Кэри СК, Уильямсон К.Э., Хелтон Р.Р., Бенч С.Р., Вингет Д., Уоммак К.Е. (ноябрь 2008 г.). «Лизогенные взаимодействия вируса и хозяина преобладают в глубоководных гидротермальных источниках с диффузным потоком». Журнал ISME . 2 (11): 1112–1121. дои : 10.1038/ismej.2008.73 . ПМИД 18719614 . S2CID 23516254 .
- ^ Jump up to: а б Калли А.И., Шакья М., Ланг А.С. (2015). «Вирусная эволюция на пределе» . В Бейкерманс С (ред.). Микробная эволюция в экстремальных условиях . Вальтер де Грюйтер ГмбХ & Ко КГ. стр. 209–222. ISBN 978-3-11-038964-7 .
- ^ Хэ Т, Ли Х, Чжан X (июль 2017 г.). «Вирусы глубоководных гидротермальных источников компенсируют микробный метаболизм во взаимодействиях вирус-хозяин» . мБио . 8 (4): mBio.00893–17, e00893–17. дои : 10.1128/mBio.00893-17 . ПМК 5513705 . ПМИД 28698277 .
- ^ Анантараман К., Дюхайм М.Б., Брейер Дж.А., Вендт К.А., Тонер Б.М., Дик Г.Дж. (май 2014 г.). «Гены окисления серы в различных глубоководных вирусах». Наука . 344 (6185): 757–760. Бибкод : 2014Sci...344..757A . дои : 10.1126/science.1252229 . HDL : 1912/6700 . ПМИД 24789974 . S2CID 692770 .
- ^ Уолл Т (2 мая 2014 г.). «Вирусы делают зомби глубоководных бактерий» . Новости АВС .
