Исправление несоответствия ДНК
| Часть серии о |
| Генетика |
|---|
 |
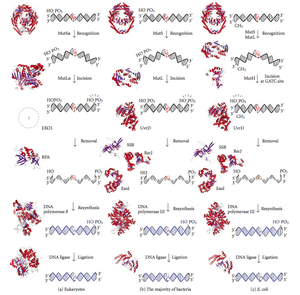
Восстановление несоответствия ДНК ( MMR ) — это система распознавания и исправления ошибочной вставки, удаления и неправильного включения оснований , которые могут возникнуть во время репликации и рекомбинации ДНК , а также восстановления некоторых форм повреждений ДНК . [1] [2]
Восстановление несоответствия зависит от цепи. Во время синтеза ДНК вновь синтезированная (дочерняя) цепь обычно содержит ошибки. Чтобы начать восстановление, механизм восстановления несоответствия отличает вновь синтезированную цепь от матрицы (родительской). У грамотрицательных бактерий транзиторное гемиметилирование различает цепи (родительские метилированы , а дочерние нет). Однако у других прокариот и эукариот точный механизм не ясен. Предполагается, что у эукариот вновь синтезированная ДНК с отстающей цепью временно содержит разрывы (до того, как она будет запечатана ДНК-лигазой) и обеспечивает сигнал, который направляет системы проверки несоответствий на соответствующую цепь. Это означает, что эти зазубрины должны присутствовать в ведущей цепи, и недавно были найдены доказательства этого. [3] Недавняя работа [4] показали, что разрывы являются местами для RFC-зависимой загрузки репликационного скользящего зажима, ядерного антигена пролиферирующих клеток (PCNA), в зависимости от ориентации, так что одна грань белка в форме пончика прилегает к 3'-ОН. закончиться на нике. Нагруженный PCNA затем направляет действие эндонуклеазы MutLalpha. [5] к дочерней цепи при наличии несовпадения и MutSalpha или MutSbeta.
Любое мутационное событие, разрушающее суперспиральную структуру ДНК , потенциально может поставить под угрозу генетическую стабильность клетки. Тот факт, что системы обнаружения и восстановления повреждений так же сложны, как и сам механизм репликации, подчеркивает важность, которую эволюция придает точности ДНК.
Примеры несовпадающих оснований включают спаривание G/T или A/C (см. Репарация ДНК ). Несовпадения обычно возникают из-за таутомеризации оснований во время репликации ДНК. Повреждение устраняется путем распознавания деформации, вызванной несоответствием, определения матрицы и цепи, не являющейся матрицей, а также удаления неправильно включенного основания и замены его правильным нуклеотидом . Процесс удаления включает в себя нечто большее, чем просто сам несовпадающий нуклеотид. От нескольких до тысяч пар оснований вновь синтезированной цепи ДНК можно удалить.
Белки репарации несоответствия
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Май 2024 г. ) |
| Белок восстановления несоответствия ДНК, С-концевой домен | |||
|---|---|---|---|
 hpms2-atpgs | |||
| Идентификаторы | |||
| Символ | DNA_mis_repair | ||
| Пфам | PF01119 | ||
| Пфам Клан | CL0329 | ||
| ИнтерПро | ИПР013507 | ||
| PROSITE | PDOC00057 | ||
| СКОП2 | 1бкн / СКОПе / СУПФАМ | ||
| |||
Репарация ошибочных спариваний — высококонсервативный процесс от прокариот к эукариотам . Первые доказательства репарации ошибочных спариваний были получены от S. pneumoniae hexA и hexB ( гены ). Последующие исследования E. coli выявили ряд генов, мутационная инактивация которых приводит к появлению гипермутабельных штаммов. Поэтому генные продукты называются белками «Mut» и являются основными активными компонентами системы репарации ошибочных спариваний. Три из этих белков необходимы для обнаружения несоответствия и направления на него механизма восстановления: MutS , MutH и MutL (MutS является гомологом HexA и MutL HexB).
MutS образует димер (MutS2 ) , который распознает несовпадающее основание на дочерней цепи и связывает мутированную ДНК. MutH связывается с гемиметилированными сайтами дочерней ДНК, но его действие латентно и активируется только при контакте с димером MutL (MutL 2 ), который связывает комплекс MutS-ДНК и действует как посредник между MutS 2 и MutH, активируя последний. ДНК закольцована для поиска ближайшего к месту несовпадения сайта метилирования d(GATC), который может находиться на расстоянии до 1 т.п.н. При активации комплексом MutS-ДНК MutH разрывает дочернюю цепь рядом с гемиметилированным сайтом. MutL привлекает UvrD хеликазу (ДНК-хеликаза II) для разделения двух цепей с определенной полярностью от 3' до 5'. Затем весь комплекс MutSHL скользит вдоль ДНК в направлении несоответствия, освобождая цепь для вырезания по ходу ее движения. Экзонуклеаза следует за комплексом и переваривает хвост оц-ДНК. Рекрутируемая экзонуклеаза зависит от того, с какой стороны несоответствия MutH разрезает цепь – 5' или 3'. Если разрыв, сделанный MutH, находится на 5'-конце несоответствия, используется либо RecJ, либо ExoVII (обе экзонуклеазы с 5' по 3'). Если, однако, ник находится на 3'-конце несоответствия, ExoI Используется (фермент от 3' до 5').
Весь процесс заканчивается за пределами сайта несоответствия, т.е. как сам сайт, так и окружающие его нуклеотиды полностью вырезаются. Одноцепочечный разрыв, созданный экзонуклеазой, затем может быть восстановлен ДНК-полимеразой III (с помощью одноцепочечного белка), которая использует другую цепь в качестве матрицы и, наконец, запечатана ДНК-лигазой. Затем ДНК-метилаза быстро метилирует дочернюю цепь.
Гомологи MutS
[ редактировать ]При связывании димер MutS 2 изгибает спираль ДНК и защищает примерно 20 пар оснований. Он обладает слабой АТФазной активностью, и связывание АТФ приводит к образованию третичных структур на поверхности молекулы. Кристаллическая структура MutS показывает, что она исключительно асимметрична, и, хотя ее активная конформация представляет собой димер, только одна из двух половин взаимодействует с сайтом несоответствия.
У эукариот M ut Sho омологи образуют два основных гетеродимера: Msh2 /Msh6 (MutSα) и Msh2 /Msh3 (MutSβ). Путь MutSα участвует в первую очередь в замене оснований и репарации несоответствий малых петель. Путь MutSβ также участвует в репарации малой петли в дополнение к репарации большой петли (~ 10 нуклеотидных петель). Однако MutSβ не восстанавливает замены оснований.
Гомологи MutL
[ редактировать ]MutL также обладает слабой активностью АТФазы (он использует АТФ для движения). Он образует комплекс с MutS и MutH, увеличивая присутствие MutS в ДНК.
Однако процессивность (расстояние, которое фермент может пройти по ДНК перед диссоциацией) UvrD составляет всего ~ 40–50 п.н. Поскольку расстояние между разрывом, созданным MutH, и несоответствием может составлять в среднем ~ 600 п.н., если нет загруженного другого UvrD, размотанная секция затем может повторно отжигаться со своей комплементарной цепью, заставляя процесс начинаться заново. Однако при помощи MutL скорость загрузки UvrD значительно увеличивается. Хотя процессивность (и использование АТФ) отдельных молекул UvrD остается прежней, общее воздействие на ДНК значительно усиливается; у ДНК нет шансов на повторный отжиг, так как каждый UvrD раскручивает 40-50 п.н. ДНК, диссоциирует, а затем немедленно заменяется другим UvrD, повторяя процесс. Это подвергает большие участки ДНК расщеплению экзонуклеазой , что позволяет быстро удалить (а затем заменить) неправильную ДНК.
Эукариоты имеют пять M ut L, омологов обозначенных как MLH1, MLH2, MLH3, PMS1 и PMS2. Они образуют гетеродимеры, имитирующие MutL в E. coli . Человеческие гомологи прокариотического MutL образуют три комплекса, называемые MutLα, MutLβ и MutLγ. Комплекс MutLα состоит из субъединиц MLH1 и PMS2, гетеродимер MutLβ состоит из MLH1 и PMS1, тогда как MutLγ состоит из MLH1 и MLH3. MutLα действует как эндонуклеаза, которая вводит разрывы цепи в дочернюю цепь при активации несоответствием и другими необходимыми белками, MutSα и PCNA. Эти прерывания цепи служат точками входа для активности экзонуклеазы, которая удаляет несовпадающую ДНК. Роли, которые играют MutLβ и MutLγ в репарации ошибочного спаривания, менее понятны.
MutH: эндонуклеаза, присутствующая в E. coli и сальмонелле.
[ редактировать ]MutH — очень слабая эндонуклеаза , которая активируется после связывания с MutL (который сам связан с MutS). Он разрывает неметилированную ДНК и неметилированную цепь полуметилированной ДНК, но не разрушает полностью метилированную ДНК. Эксперименты показали, что репарация ошибочного спаривания является случайной, если ни одна цепь не метилирована. [ нужна ссылка ] Такое поведение привело к предположению, что MutH определяет, какая цепь содержит несовпадение. MutH не имеет эукариотического гомолога. Его эндонуклеазную функцию берут на себя гомологи MutL, которые обладают некоторой специализированной 5'-3' экзонуклеазной активностью. Смещение цепи для удаления несоответствий из вновь синтезированной дочерней цепи у эукариот может обеспечиваться свободными 3'-концами фрагментов Оказаки в новой цепи, созданной во время репликации.
PCNA β-скользящий зажим
[ редактировать ]PCNA и β-скользящий зажим ассоциированы с MutSα/β и MutS соответственно. Хотя первоначальные сообщения предполагали, что комплекс PCNA-MutSα может усиливать распознавание несовпадений, [6] это было недавно продемонстрировано [7] что нет видимых изменений в сродстве MutSα к несовпадению в присутствии или в отсутствие PCNA. Более того, мутанты MutSα, которые не способны взаимодействовать с PCNA in vitro, демонстрируют способность осуществлять распознавание несовпадений и удаление несовпадений до уровня, близкого к дикому типу. Такие мутанты дефектны в реакции репарации, направленной за счет разрыва 5'-цепи, что впервые позволяет предположить функцию MutSα на постэксцизионной стадии реакции.
Клиническое значение
[ редактировать ]Наследственные дефекты при исправлении несоответствия
[ редактировать ]Мутации в человеческих гомологах белков Mut влияют на стабильность генома, что может привести к микросателлитной нестабильности (MSI), причастной к некоторым видам рака человека. В частности, наследственный неполипозный колоректальный рак ( HNPCC или синдром Линча) обусловлен повреждением зародышевых вариантов генов, кодирующих гомологи MutS и MutL MSH2 и MLH1 соответственно, которые, таким образом, классифицируются как гены-супрессоры опухолей. Один подтип HNPCC, синдром Мьюира-Торре (MTS), связан с опухолями кожи. Если обе унаследованные копии (аллели) гена MMR несут повреждающие генетические варианты, это приводит к очень редкому и тяжелому состоянию: раковому синдрому репарации несоответствия (или конституциональному дефициту репарации несоответствия, CMMR-D), проявляющемуся в виде множественных опухолей на ранний возраст, часто опухоли толстой кишки и головного мозга . [8]
Эпигенетическое подавление генов репарации ошибочного спаривания
[ редактировать ]Спорадические виды рака с дефицитом репарации ДНК лишь изредка имеют мутацию в гене репарации ДНК, но вместо этого они имеют тенденцию иметь эпигенетические изменения, такие как метилирование промотора, которые ингибируют экспрессию гена репарации ДНК. [9] Около 13% случаев колоректального рака имеют дефицит репарации несоответствия ДНК, обычно из-за потери MLH1 (9,8%) или иногда MSH2, MSH6 или PMS2 (все ≤1,5%). [10] Для большинства спорадических колоректальных раков с дефицитом MLH1 дефицит был обусловлен метилированием промотора MLH1. [10] Другие типы рака имеют более высокую частоту потери MLH1 (см. таблицу ниже), что опять же во многом является результатом метилирования промотора гена MLH1 . Другой эпигенетический механизм, лежащий в основе дефицита MMR, может включать сверхэкспрессию микроРНК, например, уровни миР-155 обратно коррелируют с экспрессией MLH1 или MSH2 при колоректальном раке. [11]
| Тип рака | Частота дефицита при раке | Частота дефекта соседнего поля |
|---|---|---|
| Желудок | 32% [12] [13] | 24%-28% |
| Желудок (опухоли фовеолярного типа) | 74% [14] | 71% |
| Желудок в Кашмирской долине с высокой заболеваемостью | 73% [15] | 20% |
| Пищеводный | 73% [16] | 27% |
| Плоскоклеточный рак головы и шеи (HNSCC) | 31%-33% [17] [18] | 20%-25% |
| Немелкоклеточный рак легкого (НМРЛ) | 69% [19] | 72% |
| Колоректальный | 10% [10] |
Сбои MMR при полевых дефектах
[ редактировать ]Дефект поля (канцеризация поля) — это участок эпителия, обусловленный эпигенетическими или генетическими изменениями, предрасполагающими его к развитию рака. Как отметил Рубин, «…есть доказательства того, что более 80% соматических мутаций, обнаруженных в мутаторных фенотипах колоректальных опухолей человека, возникают до начала терминальной клональной экспансии». [20] [21] Аналогично, Фогельштейн и др. [22] отмечают, что более половины соматических мутаций, выявленных в опухолях, возникали в предопухолевой фазе (в полевом дефекте), во время роста внешне нормальных клеток.
Дефицит MLH1 часто встречался в полевых дефектах (гистологически нормальных тканях), окружающих опухоли; см. таблицу выше. Эпигенетически подавленный или мутированный MLH1, скорее всего, не даст селективного преимущества стволовой клетке, однако это приведет к увеличению частоты мутаций, и один или несколько мутированных генов могут обеспечить клетке селективное преимущество. Дефицитный ген MLH1 может затем использоваться как селективно почти нейтральный ген пассажира (автостопщика), когда мутировавшая стволовая клетка генерирует расширенный клон. Продолжающееся присутствие клона с эпигенетически репрессированным MLH1 будет продолжать генерировать дальнейшие мутации, некоторые из которых могут привести к опухоли.
MSI и реакция блокады иммунных контрольных точек
[ редактировать ]Первоначально в исследовании, изучавшем участников, реагирующих на анти-PD1, было обнаружено, что мутации репарации MMR и ошибочного спаривания связаны с эффективностью блокады иммунных контрольных точек. [23] Связь между положительностью MSI и положительным ответом на анти-PD1 была впоследствии подтверждена в проспективном клиническом исследовании и одобрена FDA. [24]
Компоненты MMR у человека
[ редактировать ]У людей семь белков восстановления несоответствий ДНК (MMR) ( MLH1 , MLH3 , MSH2 , MSH3 , MSH6 , PMS1 и PMS2 ) работают скоординировано, последовательно, инициируя восстановление несоответствий ДНК. [25] Кроме того, существуют Exo1 -зависимые и Exo1-независимые подпути MMR. [26]
Другие генные продукты, участвующие в восстановлении ошибочных спариваний (после инициации генами MMR) у людей, включают ДНК-полимеразу дельта , PCNA , RPA , HMGB1 , RFC и ДНК-лигазу I , а также гистон и хроматин . факторы, модифицирующие [27] [28]
В определенных обстоятельствах путь MMR может рекрутировать склонную к ошибкам ДНК-полимеразу эта ( POLH ). Это происходит в B-лимфоцитах во время соматической гипермутации , когда POLH используется для внесения генетических изменений в гены антител. [29] Однако этот подверженный ошибкам путь MMR может запускаться в других типах клеток человека при воздействии генотоксинов. [30] и действительно, он широко активен при различных видах рака у человека, вызывая мутации, которые несут признаки активности ПОЛГ. [31]
MMR и частота мутаций
[ редактировать ]Распознавание и исправление несовпадений и инделей важно для клеток, поскольку неспособность сделать это приводит к микросателлитной нестабильности (MSI) и повышенной частоте спонтанных мутаций (мутаторный фенотип). По сравнению с другими типами рака, рак с дефицитом MMR (MSI) имеет очень высокую частоту мутаций, близкую к меланоме и раку легких. [32] типы рака, вызванные сильным воздействием УФ-излучения и мутагенных химикатов.
Помимо очень высокого мутационного бремени, дефицит MMR приводит к необычному распределению соматических мутаций по человеческому геному: это предполагает, что MMR преимущественно защищает богатые генами, рано реплицирующиеся эухроматиновые области. [33] Напротив, бедные генами и поздно реплицирующиеся гетерохроматические области генома демонстрируют высокую частоту мутаций во многих опухолях человека. [34]
Модификация гистона H3K36me3 , эпигенетическая метка активного хроматина, обладает способностью рекрутировать комплекс MSH2-MSH6 (hMutSα). [35] Соответственно, области генома человека с высоким уровнем H3K36me3 накапливают меньше мутаций из-за активности MMR. [31]
Потеря нескольких путей репарации ДНК в опухолях
[ редактировать ]Отсутствие MMR часто происходит в сочетании с потерей других генов репарации ДНК. [9] Например, гены MMR MLH1 и MLH3, а также 11 других генов репарации ДНК (таких как MGMT и многие гены пути NER ) были значительно снижены как в астроцитомах более низкой, так и в более высокой степени , в отличие от нормальной ткани головного мозга. [36] Более того, экспрессия MLH1 и MGMT тесно коррелировала в 135 образцах рака желудка, а потеря MLH1 и MGMT, по-видимому, синхронно ускорялась во время прогрессирования опухоли. [37]
Недостаточная экспрессия нескольких генов репарации ДНК часто обнаруживается при раке. [9] и может способствовать возникновению тысяч мутаций, обычно встречающихся при раке (см. Частота мутаций при раке ).
Старение
[ редактировать ]Популярной идеей, не получившей существенной экспериментальной поддержки, является идея о том, что мутация, в отличие от повреждения ДНК, является основной причиной старения. Мыши, дефектные по mutL гомологу Pms2, имеют примерно в 100 раз повышенную частоту мутаций во всех тканях, но, по-видимому, не стареют быстрее. [38] Эти мыши в основном развиваются и живут нормально, за исключением раннего канцерогенеза и мужского бесплодия.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Айер Р.Р., Плучиенник А., Бердетт В., Модрич П.Л. (февраль 2006 г.). «Репарация несоответствия ДНК: функции и механизмы». Химические обзоры . 106 (2): 302–23. дои : 10.1021/cr0404794 . ПМИД 16464007 .
- ^ Ларреа А.А., Лухан С.А., Кункель Т.А. (май 2010 г.). «SnapShot: восстановление несоответствия ДНК» . Клетка . 141 (4): 730–730.e1. дои : 10.1016/j.cell.2010.05.002 . ПМИД 20478261 . S2CID 26969788 .
- ^ Хеллер Р.К., Марианс К.Дж. (декабрь 2006 г.). «Сборка реплисом и прямой перезапуск остановившихся вилок репликации». Обзоры природы. Молекулярно-клеточная биология . 7 (12): 932–43. дои : 10.1038/nrm2058 . ПМИД 17139333 . S2CID 27666329 .
- ^ Плучиенник А, Дзантиев Л, Айер Р.Р., Константин Н, Кадыров Ф.А., Модрич П. (сентябрь 2010 г.). «Функция PCNA в активации и направлении цепи эндонуклеазы MutLα при репарации ошибочного спаривания» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (37): 16066–71. дои : 10.1073/pnas.1010662107 . ПМЦ 2941292 . ПМИД 20713735 .
- ^ Кадыров Ф.А., Дзантиев Л., Константин Н., Модрич П. (июль 2006 г.). «Эндонуклеолитическая функция MutLalpha в восстановлении несоответствия человека» . Клетка . 126 (2): 297–308. дои : 10.1016/j.cell.2006.05.039 . ПМИД 16873062 . S2CID 15643051 .
- ^ Флорес-Розас Х., Кларк Д., Колоднер Р.Д. (ноябрь 2000 г.). «Пролиферирующий клеточный ядерный антиген и Msh2p-Msh6p взаимодействуют, образуя активный комплекс распознавания неправильной пары». Природная генетика . 26 (3): 375–8. дои : 10.1038/81708 . ПМИД 11062484 . S2CID 20861705 .
- ^ Айер Р.Р., Польхаус Т.Дж., Чен С., Хура Г.Л., Дзантиев Л., Биз Л.С., Модрич П. (май 2008 г.). «Взаимодействие MutSalpha-пролиферирующих клеток с ядерным антигеном при восстановлении несоответствия ДНК человека» . Журнал биологической химии . 283 (19): 13310–9. дои : 10.1074/jbc.M800606200 . ПМЦ 2423938 . ПМИД 18326858 .
- ^ Интернет-менделевское наследование у человека (OMIM): 276300
- ^ Jump up to: а б с Бернштейн С., Бернштейн Х. (май 2015 г.). «Эпигенетическое снижение репарации ДНК при прогрессировании рака желудочно-кишечного тракта» . Всемирный журнал желудочно-кишечной онкологии . 7 (5): 30–46. дои : 10.4251/wjgo.v7.i5.30 . ПМЦ 4434036 . ПМИД 25987950 .
- ^ Jump up to: а б с Трунингер К., Менигатти М., Луз Дж., Рассел А., Хайдер Р., Гебберс Дж.О. и др. (май 2005 г.). «Иммуногистохимический анализ выявляет высокую частоту дефектов PMS2 при колоректальном раке» . Гастроэнтерология . 128 (5): 1160–71. дои : 10.1053/j.gastro.2005.01.056 . ПМИД 15887099 .
- ^ Валери Н., Гаспарини П., Фаббри М., Бракони С., Веронезе А., Ловат Ф. и др. (апрель 2010 г.). «Модуляция репарации ошибочных спариваний и стабильности генома с помощью миР-155» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (15): 6982–7. Бибкод : 2010PNAS..107.6982V . дои : 10.1073/pnas.1002472107 . ПМЦ 2872463 . ПМИД 20351277 .
- ^ Купчинскайте-Норейкене Р., Скицевичене Ю., Йонайтис Л., Угенскене Р., Купчинскас Ю., Маркелис Р. и др. (2013). «Метилирование CpG-островков генов MLH1, MGMT, DAPK и CASP8 в раковых и прилегающих к ним нераковых тканях желудка» . Лекарство . 49 (8): 361–6. дои : 10.3390/medicine49080056 . ПМИД 24509146 .
- ^ Ваки Т., Тамура Г., Цучия Т., Сато К., Нисидзука С., Мотояма Т. (август 2002 г.). «Статус метилирования промотора генов E-кадгерина, hMLH1 и p16 в неопухолевом эпителии желудка» . Американский журнал патологии . 161 (2): 399–403. дои : 10.1016/S0002-9440(10)64195-8 . ПМК 1850716 . ПМИД 12163364 .
- ^ Эндо Ю., Тамура Г., Аджиока Ю., Ватанабэ Х., Мотояма Т. (сентябрь 2000 г.). «Частое гиперметилирование промотора гена hMLH1 при дифференцированных опухолях желудка с желудочным фовеолярным фенотипом» . Американский журнал патологии . 157 (3): 717–22. дои : 10.1016/S0002-9440(10)64584-1 . ЧВК 1949419 . ПМИД 10980110 .
- ^ Вани М., Афроз Д., Махдуми М., Хамид И., Вани Б., Бхат Дж. и др. (2012). «Статус метилирования промотора гена репарации ДНК (hMLH1) у пациентов с карциномой желудка в Кашмирской долине» (PDF) . Азиатско-Тихоокеанский журнал профилактики рака . 13 (8): 4177–81. дои : 10.7314/apjcp.2012.13.8.4177 . ПМИД 23098428 .
- ^ Чанг З, Чжан В, Чанг З, Сун М, Цинь Ю, Чанг Ф и др. (январь 2015 г.). «Характеристики экспрессии FHIT , p53 , BRCA2 и MLH1 в семьях с историей рака пищевода в регионе с высокой заболеваемостью раком пищевода» . Письма об онкологии . 9 (1): 430–436. дои : 10.3892/ол.2014.2682 . ПМК 4246613 . ПМИД 25436004 .
- ^ Тауфик Х.М., Эль-Максуд Н.М., Хак Б.Х., Эль-Щербини Ю.М. (2011). «Плоскоклеточный рак головы и шеи: иммуногистохимия восстановления несоответствия и гиперметилирование промотора гена hMLH1». Американский журнал отоларингологии . 32 (6): 528–36. дои : 10.1016/j.amjoto.2010.11.005 . ПМИД 21353335 .
- ^ Цзо С., Чжан Х., Спенсер Х.Дж., Вурал Е., Суен Дж.Ю., Шичман С.А. и др. (октябрь 2009 г.). «Повышенная микросателлитная нестабильность и эпигенетическая инактивация гена hMLH1 при плоскоклеточном раке головы и шеи». Отоларингология – хирургия головы и шеи . 141 (4): 484–90. дои : 10.1016/j.otohns.2009.07.007 . ПМИД 19786217 . S2CID 8357370 .
- ^ Сафар А.М., Спенсер Х., Су Х, Коффи М., Куни К.А., Ратнасингхе Л.Д. и др. (июнь 2005 г.). «Профилирование метилирования архивного немелкоклеточного рака легкого: многообещающая прогностическая система» . Клинические исследования рака . 11 (12): 4400–5. дои : 10.1158/1078-0432.CCR-04-2378 . ПМИД 15958624 .
- ^ Рубин Х (март 2011 г.). «Поля и канцеризация полей: предопухолевые источники рака: бессимптомные гиперпластические поля являются предшественниками неоплазии, и их прогрессирование в опухоли можно отследить по плотности насыщения в культуре». Биоэссе . 33 (3): 224–31. doi : 10.1002/bies.201000067 . ПМИД 21254148 . S2CID 44981539 .
- ^ Цао Дж.Л., Ятабе Ю., Саловаара Р., Ярвинен Х.Дж., Меклин Дж.П., Аалтонен Л.А. и др. (февраль 2000 г.). «Генетическая реконструкция историй отдельных колоректальных опухолей» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (3): 1236–41. Бибкод : 2000PNAS...97.1236T . дои : 10.1073/pnas.97.3.1236 . ПМК 15581 . ПМИД 10655514 .
- ^ Фогельштейн Б., Пападопулос Н., Велкулеску В.Е., Чжоу С., Диас Л.А., Кинцлер К.В. (март 2013 г.). «Пейзажи генома рака» . Наука . 339 (6127): 1546–58. Бибкод : 2013Sci...339.1546V . дои : 10.1126/science.1235122 . ПМЦ 3749880 . ПМИД 23539594 .
- ^ Ризви, Найер; Хеллманн, Мэтью; Снайдер, Александра; Квистборг, Пиа; Макаров Владимир; Гавел, Джонатан; Ли, Уильям; Юань, Цзянда; Вонг, Филипп; Эй, Тереза; Миллер, Мартин; Рехтман, Наташа; Морейра, Андра; Ибрагим, Фавзия; Брюггеман, Кэмерон; Гасми, Биллель; Заппасоди, Роберта; Маэда, Юка; Сандер, Крис; Гарон, Эдвард; Мергуб, Таха; Волчок, Джедд; Шумахер, Тон; Тимоти, Чан (2015). «Мутационный ландшафт определяет чувствительность к блокаде PD-1 при немелкоклеточном раке легкого» . Наука . 6230 (348): 124–128. дои : 10.1126/science.aaa1348 . ПМЦ 4993154 . ПМИД 25765070 .
- ^ Центр оценки и исследования лекарств. «Одобренные лекарственные средства – FDA предоставляет ускоренное одобрение пембролизумабу для первых показаний, не зависящих от ткани/места» . www.fda.gov . Проверено 24 мая 2017 г.
- ^ Пал Т., Пермут-Вей Дж., Селлерс Т.А. (август 2008 г.). «Обзор клинической значимости дефицита репарации несоответствия при раке яичников» . Рак . 113 (4): 733–42. дои : 10.1002/cncr.23601 . ПМК 2644411 . ПМИД 18543306 .
- ^ Гёлльнер EM, Патнэм CD, Колоднер РД (август 2015 г.). «Экзонуклеаза 1-зависимая и независимая репарация ошибочного спаривания» . Восстановление ДНК . 32 : 24–32. дои : 10.1016/j.dnarep.2015.04.010 . ПМЦ 4522362 . ПМИД 25956862 .
- ^ Ли ГМ (январь 2008 г.). «Механизмы и функции восстановления несоответствий ДНК» . Клеточные исследования . 18 (1): 85–98. дои : 10.1038/cr.2007.115 . ПМИД 18157157 .
- ^ Ли ГМ (июль 2014 г.). «Новые идеи и проблемы в восстановлении несоответствий: преодоление хроматинового препятствия» . Восстановление ДНК . 19 : 48–54. дои : 10.1016/j.dnarep.2014.03.027 . ПМК 4127414 . ПМИД 24767944 .
- ^ Чахван Р., Эдельманн В., Шарфф, доктор медицинских наук, Роа С. (август 2012 г.). «Помощь разнообразию антител путем исправления ошибочных несоответствий» . Семинары по иммунологии . 24 (4): 293–300. дои : 10.1016/j.smim.2012.05.005 . ПМЦ 3422444 . ПМИД 22703640 .
- ^ Се П. (сентябрь 2012 г.). «Устранение несоответствия ДНК: доктор Джекилл и мистер Хайд?» . Молекулярная клетка . 47 (5): 665–6. doi : 10.1016/j.molcel.2012.08.020 . ПМК 3457060 . ПМИД 22980456 .
- ^ Jump up to: а б Супек Ф., Ленер Б. (июль 2017 г.). «Сигнатуры кластерных мутаций показывают, что склонная к ошибкам репарация ДНК нацелена на мутации в активных генах» . Клетка . 170 (3): 534–547.e23. дои : 10.1016/j.cell.2017.07.003 . hdl : 10230/35343 . ПМИД 28753428 .
- ^ Туна М., Амос С.И. (ноябрь 2013 г.). «Геномное секвенирование при раке» . Письма о раке . 340 (2): 161–70. дои : 10.1016/j.canlet.2012.11.004 . ПМЦ 3622788 . ПМИД 23178448 .
- ^ Супек Ф., Ленер Б. (май 2015 г.). «Дифференциальное восстановление несоответствий ДНК лежит в основе изменения скорости мутаций в геноме человека» . Природа . 521 (7550): 81–4. Бибкод : 2015Natur.521...81S . дои : 10.1038/nature14173 . ПМЦ 4425546 . ПМИД 25707793 .
- ^ Шустер-Беклер Б., Ленер Б. (август 2012 г.). «Организация хроматина оказывает большое влияние на уровень региональных мутаций в раковых клетках человека». Природа . 488 (7412): 504–7. Бибкод : 2012Natur.488..504S . дои : 10.1038/nature11273 . ПМИД 22820252 . S2CID 205229634 .
- ^ Ли Ф, Мао Г, Тонг Д, Хуан Дж, Гу Л, Ян В, Ли ГМ (апрель 2013 г.). «Гистоновая метка H3K36me3 регулирует восстановление несоответствия ДНК человека посредством взаимодействия с MutSα» . Клетка . 153 (3): 590–600. дои : 10.1016/j.cell.2013.03.025 . ПМЦ 3641580 . ПМИД 23622243 .
- ^ Цзян Цз, Ху Дж, Ли Х, Цзян Ю, Чжоу В, Лу Д (декабрь 2006 г.). «Анализ экспрессии 27 генов репарации ДНК в астроцитоме с помощью массива TaqMan низкой плотности». Письма по неврологии . 409 (2): 112–7. дои : 10.1016/j.neulet.2006.09.038 . ПМИД 17034947 . S2CID 54278905 .
- ^ Китадзима Ю, Миядзаки К, Мацукура С, Танака М, Секигути М (2003). «Потеря экспрессии ферментов репарации ДНК MGMT, hMLH1 и hMSH2 во время прогрессирования опухоли при раке желудка» . Рак желудка . 6 (2): 86–95. дои : 10.1007/s10120-003-0213-z . ПМИД 12861399 .
- ^ Нарайанан Л., Фритцелл Дж.А., Бейкер С.М., Лискай Р.М., Глейзер П.М. (апрель 1997 г.). «Повышенные уровни мутаций во многих тканях мышей с дефицитом гена репарации несоответствия ДНК Pms2» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (7): 3122–7. Бибкод : 1997PNAS...94.3122N . дои : 10.1073/pnas.94.7.3122 . ЧВК 20332 . ПМИД 9096356 .
Дальнейшее чтение
[ редактировать ]- Се П., Ямане К. (2008). «Репарация несоответствия ДНК: молекулярный механизм, рак и старение» . Механизмы старения и развития . 129 (7–8): 391–407. дои : 10.1016/j.mad.2008.02.012 . ПМК 2574955 . ПМИД 18406444 .
- Айер Р.Р., Плучиенник А., Бердетт В., Модрич П.Л. (февраль 2006 г.). «Репарация несоответствия ДНК: функции и механизмы». Химические обзоры . 106 (2): 302–23. дои : 10.1021/cr0404794 . ПМИД 16464007 .
- Джозеф Н., Дуппатла В., Рао Д.Н. (2006). Репарация несоответствия ДНК прокариот . Том. 81. стр. 1–49. дои : 10.1016/S0079-6603(06)81001-9 . ISBN 9780125400817 . ПМИД 16891168 .
{{cite book}}:|journal=игнорируется ( помогите ) - Ян В. (август 2000 г.). «Структура и функции белков репарации ошибочного спаривания» . Мутационные исследования . 460 (3–4): 245–56. дои : 10.1016/s0921-8777(00)00030-6 . ПМИД 10946232 .
- Гриффитс Дж. Ф., Гилберт В.М., Левонтин Р.К., Весслер С.Р., Сузуки Д.Т., Миллер Дж.Х. (2004). Введение в генетический анализ (8-е изд.). Нью-Йорк, штат Нью-Йорк: Фриман. ISBN 978-0-7167-4939-4 .
- Кункель Т.А., Эри Д.А. (2005). «Устранение несоответствия ДНК» . Ежегодный обзор биохимии . 74 : 681–710. doi : 10.1146/annurev.biochem.74.082803.133243 . ПМИД 15952900 .
- Фридберг ЕС, Уокер Г.К., Зиде В., Вуд Р.Д. , Шульц Р.А., Элленбергер (2005). Репарация ДНК и мутагенез (2-е изд.). Вашингтон, округ Колумбия: ASM Press. ISBN 978-1-55581-319-2 .
Внешние ссылки
[ редактировать ]- Восстановление ДНК. Архивировано 12 февраля 2018 г. в Wayback Machine.
- ДНК + несовпадение + восстановление в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)