Фрагменты Оказаки

Фрагменты Окадзаки представляют собой короткие последовательности ДНК нуклеотидов (длиной примерно от 150 до 200 пар оснований у эукариот ), которые синтезируются прерывисто и позже соединяются вместе с помощью фермента ДНК-лигазы , образуя отстающую цепь во время репликации ДНК . [1] Они были открыты в 1960-х годах японскими молекулярными биологами Рейджи и Цунэко Окадзаки вместе с некоторыми из их коллег.
Во время репликации ДНК двойная спираль раскручивается, и комплементарные цепи разделяются ферментом ДНК-хеликазой , создавая так называемую вилку репликации ДНК . После этого разветвления ДНК-примаза и ДНК-полимераза начинают действовать, создавая новую комплементарную цепь. Поскольку эти ферменты могут работать только в направлении от 5’ к 3’, две развернутые цепи матрицы реплицируются по-разному. [2] Одна цепь, ведущая цепь , подвергается непрерывному процессу репликации, поскольку ее матричная цепь имеет направленность от 3' до 5', что позволяет полимеразе, собирающей ведущую цепь, следовать за репликационной вилкой без перерыва. Однако отстающая цепь не может создаваться непрерывно, поскольку ее матричная цепь имеет направленность от 5' к 3', что означает, что полимераза должна действовать в обратном направлении от репликационной вилки. Это вызывает периодические перерывы в процессе создания отстающей пряди. Примаза и полимераза движутся в направлении, противоположном вилке, поэтому ферментам приходится неоднократно останавливаться и запускаться снова, в то время как ДНК-хеликаза разрывает цепи на части. После образования фрагментов ДНК-лигаза соединяет их в одну непрерывную цепь. [3] Весь процесс репликации считается «полунепрерывным», поскольку одна из новых цепей формируется непрерывно, а другая — нет. [4]
[2] В 1960-е годы Рейджи и Цунэко Оказаки проводили эксперименты по репликации ДНК в бактерии Escherichia coli . До этого времени считалось, что репликация представляет собой непрерывный процесс для обеих цепей, но открытия, связанные с E. coli, привели к новой модели репликации. Ученые обнаружили, что процесс репликации имеет прерывистый характер, путем импульсного мечения ДНК и наблюдения за изменениями, указывающими на несмежную репликацию.
Эксперименты [ править ]
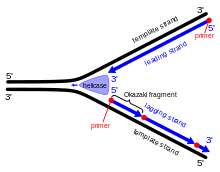
Работа Кивако Сакабе, Рейджи Окадзаки и Цунэко Окадзаки предоставила экспериментальные доказательства, подтверждающие гипотезу о том, что репликация ДНК является прерывистым процессом. Ранее было общепринято, что репликация является непрерывной как в направлении от 3' к 5', так и от 5' к 3'. 3' и 5' представляют собой атомы углерода с конкретными номерами в дезоксирибозном кольце нуклеиновых кислот и относятся к ориентации или направленности цепи. В 1967 году Цунэко Оказаки и Тору Огава предположили, что не существует механизма, который бы демонстрировал непрерывную репликацию в направлении от 3’ к 5’, а только от 5’ к 3’ с использованием ДНК-полимеразы , фермента репликации. Команда предположила, что если использовать прерывистую репликацию, короткие цепи ДНК , синтезированные в точке репликации, могут быть прикреплены в направлении от 5’ к 3’ к более старой цепи. [5]
Чтобы отличить метод репликации, используемый ДНК в эксперименте, команда импульсно пометила вновь реплицированные участки хромосом Escherichia coli , денатурировала и извлекла ДНК. Большое количество радиоактивных коротких единиц означало, что метод репликации, вероятно, был прерывистым. Гипотеза была дополнительно подтверждена открытием полинуклеотидлигазы — фермента, который связывает вместе короткие цепи ДНК. [6]
В 1968 году Рейджи и Цунэко Окадзаки собрали дополнительные доказательства зарождающихся нитей ДНК. Они предположили, что если прерывистая репликация, включающая короткие цепи ДНК, связанные вместе полинуклеотидлигазой, является механизмом синтеза ДНК, то «вновь синтезированные короткие цепи ДНК будут накапливаться в клетке в условиях, когда функция лигазы временно нарушается». E. coli были инфицированы бактериофагом Т4 , который продуцирует термочувствительную полинуклеотидлигазу. Клетки, инфицированные фагами Т4, при воздействии высоких температур накапливали большое количество коротких вновь синтезированных цепей ДНК, как и предсказывалось в гипотезе. Этот эксперимент дополнительно подтвердил гипотезу Оказакиса о прерывистой репликации и связывании с помощью полинуклеотидлигазы. Это опровергло представление о том, что короткие цепочки образуются и в процессе экстракции. [7]
Эксперименты Окадзаки предоставили обширную информацию о процессе репликации ДНК и существовании коротких, вновь синтезированных цепочек ДНК, которые позже стали известны как фрагменты Оказаки.
Пути [ править ]
Для обработки фрагментов Оказаки были предложены два пути: путь короткого лоскута и путь длинного лоскута.
путь лоскута Короткий
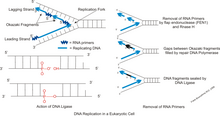
В пути короткого лоскута у эукариот отстающая цепь ДНК затравливается через короткие промежутки времени. нуклеаза FEN1 Только в коротком пути участвует . Pol δ часто сталкивается с нижележащим праймированным фрагментом Оказаки и смещает праймер-инициатор РНК/ДНК в 5'-лоскут. 5'-3' эндонуклеаза FEN1 распознает смещение 5'-лоскута и расщепляет его, создавая субстрат для лигирования. В этом методе праймер, синтезированный Pol a, удаляется. Исследования [ который? ] покажите, что в FEN1 предлагается «отслеживание; модель, в которой нуклеаза перемещается от 5'-клапана к его основанию для расщепления преформы. Pol δ не обрабатывает нуклеазную активность для расщепления смещенного лоскута. FEN1 расщепляет короткий лоскут сразу после его формирования. Расщепление ингибируется, когда 5'-конец лоскута ДНК блокируется либо комплементарным праймером, либо фрагментом стрептавидина, конъюгированным с биотином. ДНК-лигаза запечатывает разрыв, сделанный FEN1, и создает функциональную непрерывную двойную цепь ДНК. PCNA моделирует ферментативные функции белков как для FEN1, так и для ДНК-лигазы. Взаимодействие имеет решающее значение для создания правильного лигирования отстающей цепи ДНК. Последовательное смещение и расщепление цепи с помощью Pol δ и FEN1 соответственно помогает удалить всю инициаторную РНК перед лигированием. Для удаления инициаторного праймера необходимо осуществить множество смещений и провести реакции расщепления. Лоскут, который создается, обрабатывается и созревает по короткому пути лоскута.
путь лоскута Длинный
В некоторых случаях FEN1 существует лишь короткий период времени и отделяется от репликационного комплекса. Это вызывает задержку расщепления, из-за которой створки, смещенные Pol δ, становятся длинными. Когда RPA достигает достаточно большой длины, он может стабильно связываться. Когда лоскуты, связанные с RPA, рефакторизуются для расщепления FEN1, для обработки требуется другая нуклеаза, она была идентифицирована как альтернативная нуклеаза, DNA2. ДНК2 имеет дефекты сверхэкспрессии DEN1. Было показано, что DNA2 работает с FEN1 для обработки длинных лоскутов. ДНК2 может отделить RPA от длинного лоскута, используя механизм, подобный FEN1. Он связывает лоскут и продевает 5-футовый конец лоскута. Нуклеаза расщепляет лоскут, делая его слишком коротким для связывания с RPA; слишком короткий лоскут означает, что он доступен для FEN1 и лигирования. Это известно как метод длинного лоскута. ДНК2 может действовать как FEN1 в качестве резерва для активности нуклеазы, но это неэффективный процесс.
Альтернативный путь [ править ]
До недавнего времени было известно только два пути обработки фрагментов Оказаки. Однако текущие исследования пришли к выводу, что существует новый путь фрагментации Оказаки и репликации ДНК. Этот альтернативный путь включает ферменты Pol δ с Pif1, которые выполняют тот же процесс удаления лоскута, что и Pol δ и FEN1. [8]
участвующие в образовании фрагментов , Ферменты
Primase[editПримас
Примаза добавляет праймеры РНК к отстающей цепи, что позволяет синтезировать фрагменты Оказаки от 5' до 3'. Однако примаза создает праймеры РНК с гораздо меньшей скоростью, чем та, с которой ДНК-полимераза синтезирует ДНК на ведущей цепи. ДНК-полимераза на отстающей цепи также должна постоянно перерабатываться для построения фрагментов Оказаки после праймеров РНК. Это делает скорость синтеза отстающей цепи значительно ниже, чем скорость синтеза ведущей цепи. Чтобы решить эту проблему, примаза действует как временный сигнал остановки, ненадолго останавливая движение репликационной вилки во время репликации ДНК. Этот молекулярный процесс не позволяет ведущей цепи догнать отстающую. [9]
ДНК-полимераза δ [ править ]
На этом этапе новая ДНК создается ферментами, которые синтезируют ДНК в направлении от 5’ к 3’. ДНК-полимераза необходима как для ведущей цепи, которая создается в виде непрерывной цепи, так и для отстающей цепи, которая состоит из небольших частей при синтезе ДНК. Этот процесс происходит для удлинения вновь синтезированного фрагмента и изгнания сегмента РНК и ДНК. Синтез происходит в 3 фазы с участием двух разных полимераз: ДНК-полимеразы α-примазы и ДНК-полимеразы δ. Этот процесс начинается с вытеснения полимеразной α-примазы из праймера РНК и ДНК за счет эффекта репликации загрузчика зажима, этот эффект приводит к скользящему зажиму на ДНК. После этого ДНК-полимераза δ начинает переходить в голоферментную форму, из которой затем начинается синтез. Процесс синтеза будет продолжаться до тех пор, пока не достигнет 5'-конца предыдущего фрагмента Окадзаки. После прибытия обработка фрагмента Оказаки продолжается для присоединения вновь синтезированного фрагмента к отстающей цепи. Последняя функция ДНК-полимеразы δ - служить дополнением к активности 5'-флап-эндонуклеазы FEN1/RAD27. Аллель rad27-p летален в большинстве комбинаций, но жизнеспособен с полимеразой rad27-p и exo1. И полимераза rad27-p, и exo1 демонстрируют сильное синергетическое увеличение мутаций дупликации CAN 1. Единственная причина, по которой эта мутация жизнеспособна, связана с генами репарации двухцепочечных разрывов RAD50, RAD51 и RAD52. RAD27/FEN1 создает разрывы между соседними фрагментами Оказаки, сводя к минимуму количество вытеснения цепи в отстающей цепи.
ДНК-лигаза I [ править ]
Во время синтеза отстающей цепи ДНК-лигаза I соединяет фрагменты Оказаки после замены праймеров РНК нуклеотидами ДНК ДНК-полимеразой δ. Несвязанные фрагменты Оказаки могут вызвать двухцепочечные разрывы, которые расщепляют ДНК. [10] Поскольку допускается лишь небольшое количество двухцепочечных разрывов и лишь небольшое их количество можно восстановить, достаточное количество неудачных лигаций может быть фатальным для клетки.
Дальнейшие исследования предполагают дополнительную роль пролиферирующего клеточного ядерного антигена (PCNA) к функции ДНК-лигазы I по соединению фрагментов Оказаки. Когда сайт связывания PCNA на ДНК-лигазе I неактивен, способность ДНК-лигазы I соединять фрагменты Оказаки серьезно ухудшается. Таким образом, предполагаемый механизм следующий: после того, как комплекс PCNA-ДНК-полимераза δ синтезирует фрагменты Оказаки, ДНК-полимераза δ высвобождается. Затем ДНК-лигаза I связывается с PCNA, которая прикрепляется к зазубринам отстающей цепи, и катализирует образование фосфодиэфирных связей. [11] [12] [13]
Лоскутная эндонуклеаза 1
Эндонуклеаза лоскута 1 ( FEN1 ) отвечает за обработку фрагментов Оказаки. Он работает с ДНК-полимеразой для удаления праймера РНК фрагмента Оказаки и может удалять 5'-рибонуклеотид и 5'-флопы, когда ДНК-полимераза смещает цепи во время синтеза отстающей цепи. Удаление этих лоскутов включает процесс, называемый трансляцией разрыва, и создает разрыв для перевязки. Таким образом, функция FEN1 необходима для созревания фрагмента Окадзаки при формировании длинной непрерывной цепи ДНК. Аналогично, во время восстановления оснований ДНК поврежденный нуклеотид перемещается в лоскут и впоследствии удаляется FEN1. [14] [15]
Эндонуклеаза ДНК2 [ править ]
Эндонуклеаза Dna2 не имеет специфической структуры и ее свойства недостаточно изучены, но ее можно отнести к одноцепочечной ДНК со свободными концами (оцДНК). Эндонуклеаза Dna2 необходима для расщепления длинных лоскутов ДНК, которые покидают FEN1 во время процесса Оказаки. Эндонуклеаза Dna2 отвечает за удаление сегмента инициаторной РНК на фрагментах Оказаки. Кроме того, эндонуклеаза Dna2 играет ключевую роль в промежуточных продуктах, образующихся в ходе разнообразного метаболизма ДНК, и участвует в поддержании теломер. [16] [17] [18] [19] [20]
Эндонуклеаза Dna2 становится активной, когда терминальный сегмент РНК прикрепляется к 5'-концу, поскольку он перемещается в направлении от 5' к 3'. В присутствии одноцепочечного ДНК-связывающего белка RPA 5'-клапаны ДНК становятся слишком длинными, и разрывы больше не подходят в качестве субстрата для FEN1. Это не позволяет FEN1 удалить 5'-створки. Таким образом, роль Dna2 заключается в уменьшении 3'-конца этих фрагментов, что позволяет FEN1 разрезать лоскуты и делает созревание фрагментов Оказаки более эффективным. Во время процесса Окадзаки хеликаза и эндонуклеаза Dna2 неразделимы. Эндонуклеаза ДНК2 не зависит от структуры 5'-хвостой вилки своей активности. Известно, что непродуктивное связывание создает блоки расщепления и отслеживания FEN1. Известно, что АТФ снижает активность, но способствует высвобождению 3'-концевой метки. Исследования показали, что новая модель эндонуклеазы Dna2 и FEN1 частично ответственны за созревание фрагментов Оказаки. [19] [17] [16] [21]
Биологическая функция [ править ]
Вновь синтезированная ДНК, также известная как фрагменты Оказаки, связывается ДНК-лигазой, которая образует новую цепь ДНК. При синтезе ДНК образуются две нити. Ведущая цепь непрерывно синтезируется и во время этого процесса удлиняется, обнажая матрицу, которая используется для отстающей цепи (фрагменты Оказаки). В процессе репликации ДНК праймеры ДНК и РНК удаляются из отстающей цепи ДНК, чтобы обеспечить возможность связывания фрагментов Окадзаки. Поскольку этот процесс настолько распространен, созревание Оказаки будет происходить около миллиона раз за одно завершение репликации ДНК. Чтобы произошло созревание Окадзаки, праймеры РНК должны создать сегменты на лигируемых фрагментах. Он используется в качестве строительного блока для синтеза ДНК в отстающей цепи. На матричной цепи полимераза будет синтезироваться в направлении, противоположном репликационной вилке. Как только шаблон станет прерывистым, он создаст фрагмент Оказаки. Дефекты созревания фрагментов Окадзаки потенциально могут привести к разрыву нитей ДНК и вызвать различные формы хромосомных аномалий. Эти мутации в хромосомах могут влиять на внешний вид, количество наборов или количество отдельных хромосом. Поскольку хромосомы фиксированы для каждого конкретного вида, это также может изменить ДНК и вызвать дефекты в генофонде этого вида.
Различия прокариотов и эукариотов [ править ]
Фрагменты Окадзаки присутствуют как у прокариот , так и у эукариот . [22] Молекулы ДНК эукариот отличаются от кольцевых молекул прокариот тем, что они крупнее и обычно имеют несколько точек начала репликации. Это означает, что каждая эукариотическая хромосома состоит из множества реплицирующихся единиц ДНК с несколькими точками начала репликации. Для сравнения, прокариотическая ДНК имеет только один источник репликации. У эукариот эти реплицирующиеся вилки, многочисленные по всей длине ДНК, во время репликации образуют в ДНК «пузыри». Репликационная вилка формируется в определенной точке, называемой автономно реплицирующими последовательностями (ARS). У эукариотов есть комплекс зажима-загрузчика и шестизвенный зажим, называемый ядерным антигеном пролиферирующей клетки. [23] Эффективное движение репликационной вилки также критически зависит от быстрого размещения скользящих зажимов на вновь примированных участках отстающей цепи ДНК с помощью АТФ-зависимых комплексов-загрузчиков зажимов. Это означает, что кусочное генерирование фрагментов Оказаки может идти в ногу с непрерывным синтезом ДНК на ведущей цепи. Эти комплексы-зажимы-загрузчики характерны для всех эукариот и выделяют некоторые незначительные различия в синтезе фрагментов Оказаки у прокариот и эукариот. [24] Длина фрагментов Оказаки у прокариот и эукариот также различна. У прокариотов фрагменты Оказаки значительно длиннее, чем у эукариот. Эукариоты обычно имеют фрагменты Оказаки длиной от 100 до 200 нуклеотидов, тогда как фрагменты в прокариотической E. coli могут иметь длину 2000 нуклеотидов. Причина этого несоответствия неизвестна.
Каждая эукариотическая хромосома состоит из множества реплицирующихся единиц ДНК с несколькими точками начала репликации. Для сравнения, хромосома прокариот E. coli имеет только один источник репликации. Репликация у прокариот происходит внутри цитоплазмы, и при этом начинается репликация, в которой образуется примерно 100–200 и более нуклеотидов. Молекулы ДНК эукариот имеют значительно большее количество репликонов , около 50 000 и более; однако репликация не происходит одновременно на всех репликонах. У эукариот репликация ДНК происходит в ядре. Многосубъединичная форма репликации всего в одной реплицирующейся молекуле ДНК, начало репликации ДНК отодвигается многосубъединичным белком. Эта репликация медленная, иногда добавляется около 100 нуклеотидов в секунду.
Из этого мы исходим, что прокариотические клетки более просты по строению, у них нет ядра, органелл и очень мало ДНК в виде одной хромосомы. Эукариотические клетки имеют ядро с множеством органелл и большим количеством ДНК, расположенной в линейных хромосомах. Мы также видим, что размер — еще одно различие между этими прокариотическими и эукариотическими клетками. В средней эукариотической клетке примерно в 25 раз больше ДНК, чем в прокариотической клетке. Репликация в прокариотических клетках происходит гораздо быстрее, чем в эукариотических клетках; бактериям иногда требуется всего 40 минут, тогда как животным клеткам может потребоваться до 400 часов. У эукариотов также есть особая операция по репликации теломер на концах их последних хромосом. Прокариоты имеют кольцевые хромосомы, поэтому их концы не синтезируются. У прокариот есть короткий процесс репликации, который происходит непрерывно; эукариотические клетки, с другой стороны, осуществляют репликацию ДНК только во время S-фазы клеточного цикла .
Сходства — это этапы репликации ДНК. И у прокариот, и у эукариот репликация осуществляется путем раскручивания ДНК ферментом, называемым ДНК-хеликазой. Новые нити создаются ферментами, называемыми ДНК-полимеразами. Оба они следуют схожему шаблону, называемому полуконсервативной репликацией, при котором отдельные цепи ДНК образуются в разных направлениях, образуя ведущую и отстающую цепи. Эти отстающие цепи синтезируются путем образования фрагментов Окадзаки, которые вскоре соединяются. Оба этих организма начинают новые нити ДНК, которые также включают небольшие нити РНК.
Использование в технологии [ править ]
связанные с фрагментами Оказаки , Медицинские концепции

Хотя клетки проходят несколько этапов, чтобы гарантировать отсутствие мутаций в генетической последовательности, иногда специфические делеции и другие генетические изменения во время созревания фрагмента Окадзаки остаются незамеченными. Поскольку фрагменты Окадзаки представляют собой набор нуклеотидов отстающей цепи, любое изменение, включая делеции, вставки или дупликации исходной цепи, может вызвать мутацию, если она не обнаружена и не исправлена. Другие причины мутаций включают проблемы с белками, которые помогают в репликации ДНК. Например, мутация, связанная с примазой, влияет на удаление праймера РНК и может сделать цепь ДНК более хрупкой и восприимчивой к разрывам. Другая мутация касается полимеразы α, которая нарушает редактирование последовательности фрагмента Оказаки и включение белка в генетический материал. Оба изменения могут привести к хромосомным аберрациям, непреднамеренным генетическим перестройкам и различным видам рака в более позднем возрасте. [25]
Чтобы проверить влияние мутаций белка на живые организмы, исследователи генетически изменили лабораторных мышей, чтобы они были гомозиготными по другой мутации в белке, связанном с репликацией ДНК, эндонуклеазой лоскута 1 или FEN1. Результаты варьировались в зависимости от конкретных изменений генов. У гомозиготных нокаут-мутантных мышей наблюдалась «неспособность пролиферации клеток» и «ранняя эмбриональная гибель» (27). Мыши с мутациями F343A и F344A (также известными как FFAA) умерли сразу после рождения из-за осложнений при рождении, включая панцитопению легких и гипоплазию . Это связано с тем, что мутация FFAA предотвращает взаимодействие FEN1 с PCNA (ядерным антигеном пролиферирующих клеток), что, следовательно, не позволяет ему выполнить свою задачу во время созревания фрагмента Оказаки. Взаимодействие с этим белком считается ключевой молекулярной функцией биологической функции FEN1. Мутация FFAA вызывает дефекты удаления праймера РНК и восстановления пар длинных оснований, что приводит к множеству разрывов в ДНК. При тщательном наблюдении клетки, гомозиготные по мутации FFAA FEN1, по-видимому, демонстрируют лишь частичные дефекты созревания, а это означает, что мыши, гетерозиготные по мутации, смогут выжить во взрослом возрасте, несмотря на наличие множества небольших дефектов в их геномах. Однако эти разрывы неизбежно предотвращают будущую репликацию ДНК, поскольку разрыв приводит к коллапсу репликационной вилки и вызывает двухцепочечные разрывы в фактической последовательности ДНК. Со временем эти разрывы также вызывают полные разрывы хромосом, что может привести к серьезным мутациям и раку. Другие мутации были реализованы с использованием измененных версий полимеразы α, что привело к аналогичным результатам. [25]
Ссылки [ править ]
- ^ Балакришнан Л., Бамбара Р.А. (февраль 2013 г.). «Фрагментный метаболизм Оказаки» . Перспективы Колд-Спринг-Харбор в биологии . 5 (2): а010173. doi : 10.1101/cshperspect.a010173 . ПМК 3552508 . ПМИД 23378587 .
- ^ Перейти обратно: а б Окадзаки Т (11 мая 2017 г.). «Дни, сплетающие синтез отстающей цепи ДНК - личные воспоминания об открытии фрагментов Оказаки и исследованиях механизма прерывистой репликации» . Труды Японской академии. Серия Б, Физические и биологические науки . 93 (5): 322–338. Бибкод : 2017PJAB...93..322O . дои : 10.2183/pjab.93.020 . ПМЦ 5489436 . ПМИД 28496054 .
- ^ Купер GM (2000). «Репликация ДНК» . Клетка: молекулярный подход (2-е изд.). Сандерленд (Массачусетс): Sinauer Associates.
- ^ MacNeill SA (октябрь 2001 г.). «Репликация ДНК: партнеры в двухэтапной схеме Окадзаки» . Современная биология . 11 (20): Р842–Р844. дои : 10.1016/s0960-9822(01)00500-0 . ПМИД 11676941 . S2CID 15853820 .
- ^ Огава Т., Окадзаки Т. (1980). «Прерывистая репликация ДНК». Ежегодный обзор биохимии . 49 : 421–457. дои : 10.1146/annurev.bi.49.070180.002225 . ПМИД 6250445 .
- ^ Оказаки Р., Окадзаки Т., Сакабе К., Сугимото К., Сугино А. (февраль 1968 г.). «Механизм роста цепей ДНК. I. Возможный разрыв и необычная вторичная структура вновь синтезированных цепей» . Труды Национальной академии наук Соединенных Штатов Америки . 59 (2): 598–605. Бибкод : 1968PNAS...59..598O . дои : 10.1073/pnas.59.2.598 . ПМК 224714 . ПМИД 4967086 .
- ^ Сугимото К., Оказаки Т., Оказаки Р. (август 1968 г.). «Механизм роста цепей ДНК, II. Накопление вновь синтезированных коротких цепей в E. coli, инфицированных фагами Т4, дефектными по лигазе» . Труды Национальной академии наук Соединенных Штатов Америки . 60 (4): 1356–1362. Бибкод : 1968PNAS...60.1356S . дои : 10.1073/pnas.60.4.1356 . ПМК 224926 . ПМИД 4299945 .
- ^ Пайк Дж. Э., Генри Р. А., Бургерс П. М., Кэмпбелл Дж. Л., Бамбара Р. А. (декабрь 2010 г.). «Альтернативный путь обработки фрагмента Оказаки: разрешение откидных лоскутов с помощью геликазы Pif1» . Журнал биологической химии . 285 (53): 41712–41723. дои : 10.1074/jbc.M110.146894 . ПМК 3009898 . ПМИД 20959454 .
- ^ Ли Дж.Б., Хайт Р.К., Хамдан С.М., Се XS, Ричардсон К.С., ван Ойен А.М. (февраль 2006 г.). «ДНК-примаза действует как молекулярный тормоз при репликации ДНК» (PDF) . Природа . 439 (7076): 621–624. Бибкод : 2006Natur.439..621L . дои : 10.1038/nature04317 . ПМИД 16452983 . S2CID 3099842 .
- ^ Соза С, Лева В, Ваго Р, Феррари Дж, Мадзини Дж, Биамонти Дж, Монтекукко А (апрель 2009 г.). «Дефицит ДНК-лигазы I приводит к репликационно-зависимому повреждению ДНК и влияет на морфологию клеток, не блокируя прогрессирование клеточного цикла» . Молекулярная и клеточная биология . 29 (8): 2032–2041. дои : 10.1128/MCB.01730-08 . ПМЦ 2663296 . ПМИД 19223467 .
- ^ Джин Ю.Х., Айягари Р., Резник М.А., Горденин Д.А., Бургерс П.М. (январь 2003 г.). «Созревание фрагмента Оказаки в дрожжах. II. Сотрудничество между полимеразной и 3'-5'-экзонуклеазной активностью Pol delta при создании лигируемого разрыва» . Журнал биологической химии . 278 (3): 1626–1633. дои : 10.1074/jbc.M209803200 . ПМИД 12424237 .
- ^ Левин Д.С., Бай В., Яо Н., О'Доннелл М., Томкинсон А.Е. (ноябрь 1997 г.). «Взаимодействие между ДНК-лигазой I и ядерным антигеном пролиферирующей клетки: значение для синтеза и соединения фрагментов Окадзаки» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (24): 12863–12868. Бибкод : 1997PNAS...9412863L . дои : 10.1073/pnas.94.24.12863 . ПМК 24229 . ПМИД 9371766 .
- ^ Левин Д.С., Маккенна А.Е., Мотычка Т.А., Мацумото Ю., Томкинсон А.Е. (2000). «Взаимодействие между PCNA и ДНК-лигазой I имеет решающее значение для соединения фрагментов Оказаки и восстановления длинных участков путем вырезания оснований» . Современная биология . 10 (15): 919–922. дои : 10.1016/S0960-9822(00)00619-9 . ПМИД 10959839 . S2CID 14089939 .
- ^ Джин Ю.Х., Оберт Р., Бургерс П.М., Кункель Т.А., Резник М.А., Горденин Д.А. (апрель 2001 г.). «3'→5'-экзонуклеаза ДНК-полимеразы дельта может заменить эндонуклеазу 5'-клапана Rad27/Fen1 при обработке фрагментов Оказаки и предотвращении нестабильности генома» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (9): 5122–5127. дои : 10.1073/pnas.091095198 . ПМК 33174 . ПМИД 11309502 .
- ^ Лю Ю, Као ХИ, Бамбара Р.А. (2004). «Эндонуклеаза лоскута 1: центральный компонент метаболизма ДНК». Ежегодный обзор биохимии . 73 : 589–615. doi : 10.1146/annurev.biochem.73.012803.092453 . ПМИД 15189154 .
- ^ Перейти обратно: а б Бэ Ш., Ким Д.В., Ким Дж., Ким Дж.Х., Ким Д.Х., Ким Х.Д. и др. (июль 2002 г.). «Сочетание активности ДНК-хеликазы и эндонуклеазы дрожжевой ДНК2 облегчает обработку фрагмента Оказаки» . Журнал биологической химии . 277 (29): 26632–26641. дои : 10.1074/jbc.M111026200 . ПМИД 12004053 .
- ^ Перейти обратно: а б Пэ Ш., Со Ю.С. (декабрь 2000 г.). «Описание ферментативных свойств дрожжевой хеликазы/эндонуклеазы dna2 предполагает новую модель обработки фрагментов Оказаки» . Журнал биологической химии . 275 (48): 38022–38031. дои : 10.1074/jbc.M006513200 . ПМИД 10984490 .
- ^ Кан Ю.Х., Ли Ч., Со Ю.С. (апрель 2010 г.). «Dna2 на пути к обработке фрагментов Окадзаки и стабильности генома эукариот». Критические обзоры по биохимии и молекулярной биологии . 45 (2): 71–96. дои : 10.3109/10409230903578593 . ПМИД 20131965 . S2CID 23897130 .
- ^ Перейти обратно: а б Стюарт Дж.А., Кэмпбелл Дж.Л., Бамбара Р.А. (март 2009 г.). «Значение диссоциации Dna2 эндонуклеазой лоскута 1 для обработки фрагмента Оказаки в Saccharomyces cerevisiae» . Журнал биологической химии . 284 (13): 8283–8291. дои : 10.1074/jbc.M809189200 . ПМК 2659186 . ПМИД 19179330 .
- ^ Дуксин Дж.П., Мур Х.Р., Сидорова Дж., Каранджа К., Хонакер Ю., Дао Б. и др. (июнь 2012 г.). «Роль фермента Dna2 человека во время репликации ДНК, независимая от обработки фрагмента Оказаки» . Журнал биологической химии . 287 (26): 21980–21991. дои : 10.1074/jbc.M112.359018 . ПМЦ 3381158 . ПМИД 22570476 .
- ^ Айягари Р., Гомес XV, Горденин Д.А., Бургерс П.М. (январь 2003 г.). «Созревание фрагмента Оказаки в дрожжах. I. Распределение функций между FEN1 И ДНК2» . Журнал биологической химии . 278 (3): 1618–1625. дои : 10.1074/jbc.M209801200 . ПМИД 12424238 .
- ^ Мюррей С., Мертенс Б. «Являются ли фрагменты Оказаки уникальными для эукариот? Или они универсальны и поэтому присутствуют также при репликации бактериальной ДНК?» . Часто задаваемые вопросы по MCB 150 . Школа молекулярной и клеточной биологии Иллинойского университета в Урбана-Шампейн. Архивировано из оригинала 3 августа 2014 года.
- ^ «Репликация эукариотической ДНК» . Молекулярно-растительная биотехнология . multilab.biz. 29 марта 2011 г. Архивировано из оригинала 22 августа 2011 г.
- ^ Мацунага Ф, Норайс К, Фортер П, Милликаллио Х (февраль 2003 г.). «Идентификация коротких« эукариотических »фрагментов Оказаки, синтезированных из прокариотического источника репликации» . Отчеты ЭМБО . 4 (2): 154–158. дои : 10.1038/sj.embor.embor732 . ПМЦ 1315830 . ПМИД 12612604 .
- ^ Перейти обратно: а б Чжэн Л., Шэнь Б. (февраль 2011 г.). «Созревание фрагмента Оказаки: нуклеазы занимают центральное место» . Журнал молекулярно-клеточной биологии . 3 (1): 23–30. дои : 10.1093/jmcb/mjq048 . ПМК 3030970 . ПМИД 21278448 .
Дальнейшее чтение [ править ]
- Инман Р.Б., Шнос М. (март 1971 г.). «Структура точек ветвления при репликации ДНК: наличие одноцепочечных связей в точках ветвления лямбда-ДНК». Журнал молекулярной биологии . 56 (2): 319–325. дои : 10.1016/0022-2836(71)90467-0 . ПМИД 4927949 .
- Томмес П., Хюбшер У (декабрь 1990 г.). «Репликация эукариотической ДНК. Ферменты и белки, действующие на вилке» . Европейский журнал биохимии . 194 (3): 699–712. дои : 10.1111/j.1432-1033.1990.tb19460.x . ПМИД 2269294 .
Внешние ссылки [ править ]
- Фрагменты Окадзаки + в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Статья McGraw Hill Higher Education, в которой обсуждается синтез ДНК