Направленность (молекулярная биология)
Эта статья нуждается в дополнительных цитатах для проверки . ( май 2023 г. ) |
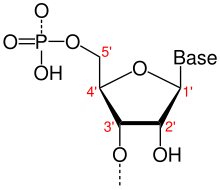
Направленность в молекулярной биологии и биохимии — это сквозная химическая ориентация одной цепи нуклеиновой кислоты . В одноцепочечной ДНК или РНК химическое соглашение об именовании атомов углерода в нуклеотидном пентозо-сахарном кольце означает, что будет 5'-конец (обычно произносится как «конец с пятью штрихами»), который часто содержит фосфатную группу. присоединен к 5'-углероду рибозного кольца и 3'-концу (обычно произносится как «трехштриховой конец»), который обычно не модифицирован заместителем рибозы -OH. В двойной спирали ДНК нити идут в противоположных направлениях, что позволяет спаривать основания между ними, что важно для репликации или транскрипции закодированной информации.
Нуклеиновые кислоты могут быть синтезированы только in vivo в направлении от 5'-к-3', поскольку полимеразы , которые собирают различные типы новых цепей, обычно полагаются на энергию, вырабатываемую при разрыве нуклеозидтрифосфатных связей для присоединения новых нуклеозидмонофосфатов к 3'-. гидроксильная (-OH) группа через фосфодиэфирную связь . Относительное расположение структур вдоль нитей нуклеиновой кислоты, включая гены белков и различные сайты связывания , обычно отмечают как расположенные либо выше (к 5'-концу), либо ниже (к 3'-концу). (См. также восходящий и нисходящий поток .)
Направленность связана со смыслом , но отличается от него . Транскрипция одноцепочечной РНК с двухцепочечной ДНК-матрицы требует выбора одной цепи ДНК-матрицы в качестве цепи матрицы, которая непосредственно взаимодействует с образующейся РНК за счет комплементарной последовательности . Другая цепь не копируется напрямую, но ее последовательность обязательно будет аналогична последовательности РНК. Сайты инициации транскрипции обычно встречаются на обеих цепях ДНК организма и определяют местоположение, направление и обстоятельства, при которых будет происходить транскрипция. Если транскрипт кодирует один или (реже) несколько белков , трансляция каждого белка рибосомой будет происходить в направлении от 5'-к-3' и будет расширять белок от его N-конца к его С-концу . Например, в типичном гене стартовый кодон (5'-ATG-3') представляет собой последовательность ДНК внутри смысловой цепи. Транскрипция начинается с сайта, расположенного выше (относительно смысловой цепи), и, проходя через этот участок, копирует 3'-TAC-5' из матричной цепи с образованием 5'-AUG-3' внутри информационная РНК (мРНК). мРНК сканируется рибосомой с 5'-конца, где стартовый кодон направляет включение метионина ( вместо этого бактерии , митохондрии и пластиды используют N -формилметионин ) на N-конце белка. По соглашению, одиночные цепи последовательностей ДНК и РНК записываются в направлении от 5' к 3', за исключением случаев, когда это необходимо для иллюстрации схемы спаривания оснований.
5'-конец
[ редактировать ]
5'-конец (произносится как «конец с пятью штрихами») обозначает конец цепи ДНК или РНК, которая имеет пятый атом углерода в сахарном кольце дезоксирибозы на своем или рибозы конце. Фосфатная группа , присоединенная к 5'-концу, обеспечивает лигирование двух нуклеотидов , т.е. ковалентное связывание 5'-фосфата с 3'-гидроксильной группой другого нуклеотида, с образованием фосфодиэфирной связи . Удаление 5'-фосфата предотвращает лигирование. Чтобы предотвратить нежелательное лигирование нуклеиновой кислоты (например, самолигирование плазмидного вектора при клонировании ДНК ), молекулярные биологи обычно удаляют 5'-фосфат с помощью фосфатазы .
5'-конец зарождающейся информационной РНК является местом, где происходит посттранскрипционное кэпирование , процесс, который жизненно важен для производства зрелой информационной РНК. Кэпирование увеличивает стабильность информационной РНК во время ее трансляции , обеспечивая устойчивость к деградационному воздействию экзонуклеаз . [1] Он состоит из метилированного нуклеотида ( метилгуанозина ), присоединенного к информационной РНК редкой 5'- и 5'-трифосфатной связью.
часто 5'- гена фланкирующий участок обозначает транскрибируется участок ДНК, который не в РНК. 5'-фланкирующая область содержит промотор гена , а также может содержать энхансеры или другие сайты связывания белка.
область 5'- нетранслируемая (5'-UTR) представляет собой область гена, которая транскрибируется в мРНК и расположена на 5'-конце мРНК. Эта область мРНК может транслироваться, а может и не транслироваться , но обычно участвует в регуляции трансляции. 5'-нетранслируемая область представляет собой часть ДНК, начинающуюся от кэп-сайта и простирающуюся до основания непосредственно перед кодоном инициации трансляции AUG основной кодирующей последовательности. Эта область может иметь последовательности, такие как сайт связывания рибосомы и последовательность Козака , которые определяют эффективность трансляции мРНК или могут влиять на стабильность мРНК.
3'-конец
[ редактировать ]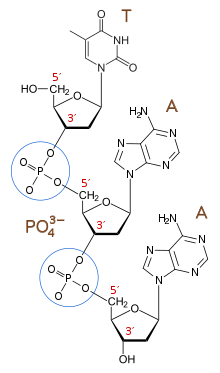
3'-конец (три штриховых конца) цепи назван так из-за того, что он оканчивается у гидроксильной группы третьего углерода сахарного кольца , и известен как хвостовой конец . 3'-гидроксил необходим для синтеза новых молекул нуклеиновой кислоты, поскольку он лигируется (присоединяется) к 5'-фосфату отдельного нуклеотида, позволяя образовывать цепи связанных нуклеотидов.
Молекулярные биологи могут использовать нуклеотиды , в которых отсутствует 3'-гидроксил (дидезоксирибонуклеотиды), для прерывания репликации ДНК . Этот метод известен как метод обрыва дидезокси-цепи или метод Сэнгера и используется для определения порядка нуклеотидов в ДНК .
3'-конец формирующейся информационной РНК является местом посттранскрипционного полиаденилирования , которое прикрепляет цепь из 50–250 остатков аденозина для образования зрелой информационной РНК. Эта цепочка помогает определить, как долго информационная РНК сохраняется в клетке, влияя на то, сколько белка вырабатывается из нее.
3'- фланкирующая область представляет собой область ДНК, которая не копируется в зрелую мРНК, но находится рядом с 3'-концом гена. Первоначально считалось, что 3'-фланкирующая ДНК вообще не транскрибируется, но было обнаружено, что она транскрибируется в РНК и быстро удаляется во время обработки первичного транскрипта с образованием зрелой мРНК. 3'-фланкирующая область часто содержит последовательности, влияющие на формирование 3'-конца сообщения. Он также может содержать энхансеры или другие сайты, с которыми могут связываться белки.
область 3'- нетранслируемая транскрибируется (3'-UTR) представляет собой область ДНК, которая в мРНК и становится 3'-концом сообщения, но не содержит последовательности, кодирующей белок. Все, что находится между стоп-кодоном и хвостом полиА, считается 3'-нетранслируемым. 3'-нетранслируемая область может влиять на эффективность трансляции мРНК или стабильность мРНК. Он также имеет последовательности, необходимые для добавления поли(А)-хвоста к сообщению, включая гексануклеотид AAAUAAA.
См. также
[ редактировать ]Дальнейшее чтение
[ редактировать ]- Харви Лодиш; Арнольд Берк; Пол Мацудайра; Крис А. Кайзер (2004). Молекулярно-клеточная биология (5-е изд.). Нью-Йорк: WH Freeman and Company. ISBN 978-0-7167-4366-8 .
- ^ Гао М., Фриц Д.Т., Форд Л.П., Вилуш Дж. (март 2000 г.). «Взаимодействие между поли(А)-специфической рибонуклеазой и 5'-кэпом влияет на скорость деаденилирования мРНК in vitro» . Молекулярная клетка . 5 (3): 479–488. дои : 10.1016/S1097-2765(00)80442-6 . ПМЦ 2811581 . ПМИД 10882133 .