Комплементарность (молекулярная биология)

В молекулярной биологии комплементарность описывает отношения между двумя структурами, каждая из которых соответствует принципу «замок и ключ». В природе комплементарность является основным принципом репликации и транскрипции ДНК, поскольку это свойство, общее для двух последовательностей ДНК или РНК , например, когда они выровнены антипараллельно друг другу, нуклеотидные основания в каждой позиции в последовательностях будут комплементарны , во многом это как смотреть в зеркало и видеть обратное. Эта дополнительная пара оснований позволяет клеткам копировать информацию из одного поколения в другое и даже находить и восстанавливать повреждения информации, хранящейся в последовательностях.
Степень комплементарности между двумя цепями нуклеиновой кислоты может варьироваться от полной комплементарности (каждый нуклеотид находится напротив своей противоположности) до отсутствия комплементарности (каждый нуклеотид не находится напротив своей противоположности) и определяет стабильность последовательностей , находящихся вместе. Более того, различные функции репарации ДНК, а также регуляторные функции основаны на комплементарности пар оснований. В биотехнологии принцип комплементарности пар оснований позволяет создавать гибриды ДНК между РНК и ДНК и открывает двери для современных инструментов, таких как библиотеки кДНК . Хотя наибольшая комплементарность наблюдается между двумя отдельными цепочками ДНК или РНК, последовательность также может иметь внутреннюю комплементарность, приводящую к связыванию последовательности сама с собой в свернутой конфигурации.
пар оснований ДНК Комплементарность и РНК


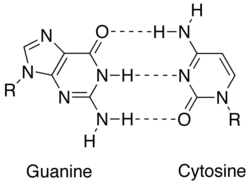
Комплементарность достигается за счет различных взаимодействий между азотистыми основаниями : аденином , тимином ( урацил в РНК ), гуанином и цитозином . Аденин и гуанин являются пуринами , а тимин, цитозин и урацил — пиримидинами . Пурины крупнее пиримидинов. Оба типа молекул дополняют друг друга и могут образовывать пары оснований только с азотистым основанием противоположного типа. В нуклеиновой кислоте нуклеиновые основания удерживаются вместе за счет водородной связи , которая эффективно работает только между аденином и тимином, а также между гуанином и цитозином. Комплемент оснований A = T имеет две водородные связи, а пара оснований G ≡ C имеет три водородные связи. Все другие конфигурации нуклеиновых оснований будут препятствовать образованию двойной спирали. Нити ДНК ориентированы в противоположных направлениях, их называют антипараллельными . [1]
| Нуклеиновая кислота | Нуклеиновые основания | Базовый комплект |
| ДНК | аденин(А), тимин(Т), гуанин(G), цитозин(С) | А = Т, Г ≡ С |
| РНК | аденин(А), урацил(U), гуанин(G), цитозин(С) | А = U, G ≡ C |
Комплементарная цепь ДНК или РНК может быть построена на основе комплементарности нуклеиновых оснований. [2] Каждая пара оснований, A = T или G ≡ C, занимает примерно одинаковое пространство, тем самым обеспечивая образование скрученной двойной спирали ДНК без каких-либо пространственных искажений. Водородные связи между азотистыми основаниями также стабилизируют двойную спираль ДНК. [3]
Комплементарность цепей ДНК в двойной спирали позволяет использовать одну цепь в качестве матрицы для построения другой. Этот принцип играет важную роль в репликации ДНК , закладывая основу наследственности , объясняя, как генетическая информация может передаваться следующему поколению. Комплементарность также используется в транскрипции ДНК , которая генерирует цепь РНК из матрицы ДНК. [4] Кроме того, вирус иммунодефицита человека , одноцепочечный РНК-вирус , кодирует РНК-зависимую ДНК-полимеразу ( обратную транскриптазу ), которая использует комплементарность для катализа репликации генома. Обратная транскриптаза может переключаться между двумя родительскими геномами выбора копии РНК путем рекомбинации во время репликации. [5]
Механизмы восстановления ДНК, такие как корректура, основаны на комплементарности и позволяют исправлять ошибки во время репликации ДНК путем удаления несовпадающих нуклеиновых оснований. [1] В общем, повреждения в одной цепи ДНК можно устранить путем удаления поврежденного участка и его замены с использованием комплементарности для копирования информации из другой цепи, как это происходит в процессах репарации ошибочных спариваний , эксцизионной репарации нуклеотидов и эксцизионной репарации оснований . [6]
Цепи нуклеиновых кислот также могут образовывать гибриды , в которых одноцепочечная ДНК может легко отжигаться с комплементарной ДНК или РНК. Этот принцип лежит в основе широко применяемых лабораторных методов, таких как полимеразная цепная реакция или ПЦР. [1]
Две цепи комплементарной последовательности называются смысловой и антисмысловой . Смысловая цепь обычно представляет собой транскрибируемую последовательность ДНК или РНК, которая образовалась при транскрипции, тогда как антисмысловая цепь представляет собой цепь, комплементарную смысловой последовательности.
Самодополняемость и шпильки [ править ]

Самокомплементарность относится к тому факту, что последовательность ДНК или РНК может сворачиваться сама в себя, создавая структуру, подобную двухцепочечной. В зависимости от того, насколько близко друг к другу расположены самодополняющие части последовательности, нить может образовывать шпильки, соединения, выпуклости или внутренние петли. [1] РНК с большей вероятностью образует подобные структуры из-за связывания пар оснований, не наблюдаемого в ДНК, например, связывания гуанина с урацилом. [1]

Регуляторные функции [ править ]
Комплементарность может быть обнаружена между короткими участками нуклеиновой кислоты и кодирующей областью или транскрибируемым геном и приводит к спариванию оснований. Эти короткие последовательности нуклеиновых кислот обычно встречаются в природе и выполняют регуляторные функции, такие как подавление генов. [1]
транскрипты Антисмысловые
Антисмысловые транскрипты представляют собой участки некодирующей мРНК, комплементарные кодирующей последовательности. [7] Полногеномные исследования показали, что антисмысловые транскрипты РНК часто встречаются в природе. Обычно считается, что они увеличивают кодирующий потенциал генетического кода и усложняют регуляцию генов. На данный момент известно, что 40% генома человека транскрибируется в обоих направлениях, что подчеркивает потенциальную значимость обратной транскрипции. [8] Было высказано предположение, что комплементарные области между смысловыми и антисмысловыми транскриптами позволят создавать гибриды двухцепочечной РНК, которые могут играть важную роль в регуляции генов. Например, мРНК фактора 1α, индуцированная гипоксией , и β-секретазы мРНК транскрибируются двунаправленно, и было показано, что антисмысловой транскрипт действует как стабилизатор смыслового сценария. [9]
микроРНК и миРНК [ править ]

микроРНК , микроРНК, представляют собой короткие последовательности РНК, которые комплементарны областям транскрибируемого гена и выполняют регуляторные функции. Текущие исследования показывают, что циркулирующая микроРНК может использоваться в качестве новых биомаркеров и, следовательно, демонстрирует многообещающие доказательства, которые можно использовать в диагностике заболеваний. [10] МикроРНК образуются из более длинных последовательностей РНК, которые ферментом Dicer отделяются от последовательности РНК, принадлежащей регуляторному гену. Эти короткие нити связываются с комплексом RISC . Они совпадают с последовательностями в верхней области транскрибируемого гена из-за их комплементарности и действуют как сайленсеры для гена тремя способами. Один из них заключается в предотвращении связывания рибосомы и инициации трансляции. Во-вторых, это разрушение мРНК, с которой связан комплекс. И третий — это создание новой последовательности двухцепочечной РНК (дцРНК), на которую Дайсер может воздействовать, создавая больше микроРНК для поиска и разрушения большего количества копий гена. Малые интерферирующие РНК (миРНК) по функциям аналогичны микроРНК; они происходят из других источников РНК, но служат той же цели, что и микроРНК. [1] Учитывая их небольшую продолжительность, правила взаимодополняемости означают, что они по-прежнему могут быть очень разборчивыми в выборе целей. Учитывая, что существует четыре варианта выбора для каждого основания в цепи и длина ми/миРНК составляет 20–22 п.н., это приводит к более чем 1 × 10 12 возможные комбинации . Учитывая, что длина генома человека составляет ~3,1 миллиарда оснований, [11] это означает, что каждая микроРНК должна случайно найти совпадение во всем геноме человека только один раз.
Целующиеся заколки [ править ]
Целующиеся шпильки образуются, когда одна цепь нуклеиновой кислоты дополняет сама себя, образуя петли РНК в форме шпильки. [12] Когда две шпильки вступают в контакт друг с другом in vivo , комплементарные основания двух нитей формируются и начинают раскручивать шпильки до тех пор, пока не образуется комплекс двухцепочечной РНК (дцРНК) или комплекс не раскручивается обратно на две отдельные нити за счет несоответствия в шпильках. Вторичная структура шпильки до поцелуя обеспечивает стабильную структуру с относительно фиксированным изменением энергии. [13] Целью этих структур является баланс между стабильностью петли шпильки и силой связывания с дополнительной нитью. Слишком сильная первоначальная привязка к неудачному месту и пряди не будут раскручиваться достаточно быстро; слишком слабое первоначальное связывание и пряди никогда полностью не сформируют желаемый комплекс. Эти шпильковые структуры позволяют выявить достаточное количество оснований, чтобы обеспечить достаточно сильную проверку первоначального связывания, и достаточно слабое внутреннее связывание, чтобы обеспечить развертывание после того, как будет найдено благоприятное совпадение. [13]
---C G---
C G ---C G---
U A C G
G C U A
C G G C
A G C G
A A A G
C U A A
U CUU ---CCUGCAACUUAGGCAGG---
A GAA ---GGACGUUGAAUCCGUCC---
G A U U
U U U C
U C G C
G C C G
C G A U
A U G C
G C ---G C---
---G C---
Kissing hairpins meeting up at the top of the loops. The complementarity
of the two heads encourages the hairpin to unfold and straighten out to
become one flat sequence of two strands rather than two hairpins.
Биоинформатика [ править ]
Комплементарность позволяет хранить информацию, содержащуюся в ДНК или РНК, в одной цепи. Комплементарную цепь можно определить по матрице и наоборот, как в библиотеках кДНК. Это также позволяет проводить анализ, например сравнивать последовательности двух разных видов. Были разработаны сокращения для записи последовательностей при наличии несоответствий (коды неоднозначности) или для ускорения чтения противоположной последовательности в дополнении (амбиграммы).
Библиотека кДНК [ править ]
Библиотека кДНК представляет собой коллекцию экспрессируемых генов ДНК, которые рассматриваются как полезный справочный инструмент в процессах идентификации и клонирования генов. Библиотеки кДНК конструируются из мРНК с использованием обратной транскриптазы (RT) РНК-зависимой ДНК-полимеразы, которая транскрибирует матрицу мРНК в ДНК. Следовательно, библиотека кДНК может содержать только вставки, предназначенные для транскрипции в мРНК. Этот процесс основан на принципе комплементарности ДНК/РНК. Конечным продуктом библиотек является двухцепочечная ДНК, которую можно вставлять в плазмиды. Следовательно, библиотеки кДНК являются мощным инструментом в современных исследованиях. [1] [14]
Коды неоднозначности [ править ]
При написании последовательностей для систематической биологии может потребоваться код IUPAC , означающий «любой из двух» или «любой из трех». Код IUPAC R (любой пурин ) комплементарен Y (любому пиримидину ) и M (амино) - K (кето). W (слабый) и S (сильный) обычно не меняются местами. [15] но в прошлом они были заменены некоторыми инструментами. [16] W и S обозначают «слабый» и «сильный» соответственно и указывают на количество водородных связей, которые нуклеотид использует для образования пары со своим комплементирующим партнером. Партнер использует одинаковое количество связей, чтобы составить дополняющую пару. [17]
Код ИЮПАК, специально исключающий один из трех нуклеотидов, может быть комплементарен коду ИЮПАК, исключающему комплементарный нуклеотид. Например, V (A, C или G — «не T») может быть дополнительным к B (C, G или T — «не A»).
| Символ [18] | Описание | Представленные базы | ||||
|---|---|---|---|---|---|---|
| А | Денин | А | 1 | |||
| С | цитозин | С | ||||
| Г | г уанин | Г | ||||
| Т | т химин | Т | ||||
| В | ты расовый | В | ||||
| В | слабый | А | Т | 2 | ||
| С | сильный | С | Г | |||
| М | а мино | А | С | |||
| К | к это | Г | Т | |||
| Р | чистота | А | Г | |||
| И | пи римидин | С | Т | |||
| Б | не А ( Б идет после А) | С | Г | Т | 3 | |
| Д | не C ( D идет после C) | А | Г | Т | ||
| ЧАС | не G ( H идет после G) | А | С | Т | ||
| V | не T ( V идет после T и U) | А | С | Г | ||
| Ни - | любая база ( не пробел) | А | С | Г | Т | 4 |
Амбиграммы [ править ]
Конкретные символы могут использоваться для создания подходящей ( двусмысленной ) нотации нуклеиновой кислоты для дополнительных оснований (например, гуанин = b , цитозин = q , аденин = n и тимин = u ), что позволяет дополнять целые последовательности ДНК простым вращение текста «вверх ногами». [19] Например, в предыдущем алфавите buqn (GTCA) читался бы как ubnq (TGAC, обратное дополнение), если его перевернуть.
- qqubqnnquunbbqnbb
- bbnqbuubnnuqqbuqq
Амбиграфические обозначения легко визуализируют дополнительные участки нуклеиновой кислоты, такие как палиндромные последовательности. [20] Эта функция расширяется при использовании пользовательских шрифтов или символов вместо обычных символов ASCII или даже символов Юникода. [20]
См. также [ править ]
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с д и ж г час Уотсон, Джеймс, Лаборатория Колд-Спринг-Харбор, Таня А. Бейкер, Массачусетский технологический институт, Стивен П. Белл, Массачусетский технологический институт, Александр Ганн, Лаборатория Колд-Спринг-Харбор, Майкл Левин, Калифорнийский университет, Беркли, Ричард Лосик, Гарвард Университет; со Стивеном К. Харрисоном, Гарвардский медицинский университет (2014). Молекулярная биология гена (Седьмое изд.). Бостон: Издательская компания Бенджамина-Каммингса. ISBN 978-0-32176243-6 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Молись, Лесли (2008). «Открытие структуры и функции ДНК: Уотсон и Крик» . Природное образование . 1 (1):100 . Проверено 27 ноября 2013 г.
- ^ Шанкар, А; Джагота, А; Миттал, Дж. (11 октября 2012 г.). «Димеры оснований ДНК стабилизируются за счет взаимодействий водородных связей, включая спаривание не-Уотсона-Крика вблизи поверхностей графита». Журнал физической химии Б. 116 (40): 12088–94. дои : 10.1021/jp304260t . ПМИД 22967176 .
- ^ Худ, Л; Галас, Д. (23 января 2003 г.). «Цифровой код ДНК» . Природа . 421 (6921): 444–8. Бибкод : 2003Natur.421..444H . дои : 10.1038/nature01410 . ПМИД 12540920 .
- ^ Роусон JMO, Николаичик О.А., Кил Б.Ф., Патак В.К., Ху WS. Рекомбинация необходима для эффективной репликации ВИЧ-1 и поддержания целостности вирусного генома. Нуклеиновые кислоты Рез. 2018;46(20):10535-10545. DOI: 10.1093/nar/gky910 PMID 30307534
- ^ Флек О., Нильсен О. Восстановление ДНК. J Cell Sci. 2004;117(Часть 4):515-517. DOI:10.1242/jcs.00952
- ^ Привет; Фогельштейн, Б; Велкулеску, ВЕ; Пападопулос, Н.; Кинзлер, К.В. (19 декабря 2008 г.). «Антисмысловые транскриптомы клеток человека» . Наука . 322 (5909): 1855–7. Бибкод : 2008Sci...322.1855H . дои : 10.1126/science.1163853 . ПМК 2824178 . ПМИД 19056939 .
- ^ Катаяма, С; Томару, Ю; Касукава, Т; Некоторые, К; Наканиши, М; Накамура, М; Нисида, Х; Яп, CC; Сузуки, М; Каваи, Дж.; Сузуки, Х; Карнинчи, П; Хаясидзаки, Ю; Уэллс, К.; Фрит, М; Раваси, Т; Панг, К.К.; Халлинан, Дж; Мэттик, Дж.; Хьюм, Д.А.; Липович Л ; Баталов С; Энгстрем, П.Г.; Мизуно, Ю; Фагихи, Массачусетс; Санделин, А; Мел, AM; Моттаги-Табар, С; Лян, З; Ленхард, Б; Валестедт, К.; Группа исследований генома RIKEN; Геномная научная группа (Основная группа проекта геномной сети); Консорциум ФАНТОМ (2 сентября 2005 г.). «Антисмысловая транскрипция в транскриптоме млекопитающих». Наука 309 (5740): 1564–6. Бибкод : 2005Наука... 309.1564R дои : 10.1126/science.1112009 . ПМИД 16141073 . S2CID 34559885 .
- ^ Фагихи, Массачусетс; Чжан, М; Хуанг, Дж; Модарреси, Ф; Ван дер Бруг, член парламента; Ноллс, Массачусетс; Куксон, MR; Сен-Лоран Ж, 3-й; Валестедт, К. (2010). «Доказательства естественного ингибирования функции микроРНК, опосредованного антисмысловыми транскриптами» . Геномная биология . 11 (5): R56. дои : 10.1186/gb-2010-11-5-r56 . ПМК 2898074 . ПМИД 20507594 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Косака, Н; Ёсиока, Ю; Хагивара, К; Томинага, Н.; Кацуда, Т; Очия, Т. (5 сентября 2013 г.). «Мусор или сокровище: внеклеточные микроРНК и межклеточная коммуникация» . Границы генетики . 4 : 173. дои : 10.3389/fgene.2013.00173 . ПМК 3763217 . ПМИД 24046777 .
- ^ «Браузер генома Ensembl 73: Homo sapiens - Сборка и создание генов» . Ensembl.org . Архивировано из оригинала 15 февраля 2013 года . Проверено 27 ноября 2013 г.
- ^ Марино, Япония; Грегориан Р.С. младший; Чанковски, Г; Кротерс, DM (9 июня 1995 г.). «Образование изогнутой спирали между шпильками РНК с комплементарными петлями». Наука . 268 (5216): 1448–54. Бибкод : 1995Sci...268.1448M . дои : 10.1126/science.7539549 . ПМИД 7539549 .
- ^ Jump up to: Перейти обратно: а б Чанг, Кентукки; Тиноко I-младший (30 мая 1997 г.). «Структура шпильочного комплекса РНК, «целующего» шпильку петли TAR ВИЧ, и его комплемент». Журнал молекулярной биологии . 269 (1): 52–66. дои : 10.1006/jmbi.1997.1021 . ПМИД 9193000 .
- ^ Ван, К.Х.; Ю, С; Джордж, РА; Карлсон, Дж.В.; Хоскинс, РА; Свирскас, Р; Стэплтон, М; Цельникер, SE (2006). «Высокопроизводительный скрининг библиотеки кДНК плазмид» . Протоколы природы . 1 (2): 624–32. дои : 10.1038/нпрот.2006.90 . ПМИД 17406289 . S2CID 205463694 .
- ^ Иеремия Фейт (2011) , таблица преобразования
- ^ arep.med.harvard.edu Страница инструмента с примечанием о примененном патче для преобразования WS.
- ^ Страница инструмента обратного дополнения с документированным преобразованием кода IUPAC, доступен исходный код.
- ^ Номенклатурный комитет Международного биохимического союза (NC-IUB) (1984). «Номенклатура не полностью определенных оснований в последовательностях нуклеиновых кислот» . Проверено 4 февраля 2008 г.
- ^ Розак Д.А. (2006). «Практические и педагогические преимущества амбиграфической записи нуклеиновых кислот». Нуклеозиды Нуклеотиды Нуклеиновые кислоты . 25 (7): 807–13. дои : 10.1080/15257770600726109 . ПМИД 16898419 . S2CID 23600737 .
- ^ Jump up to: Перейти обратно: а б Розак, Д.А.; Розак, AJ (май 2008 г.). «Простота, функциональность и разборчивость в расширенной амбиграфической записи нуклеиновых кислот» . БиоТехники . 44 (6): 811–3. дои : 10.2144/000112727 . ПМИД 18476835 .