РНК-индуцированный комплекс молчания
Комплекс молчания, индуцированный РНК , или RISC , представляет собой мультибелковый комплекс , в частности рибонуклеопротеин , который участвует в подавлении генов посредством различных путей на транскрипционном и трансляционном уровнях. [1] Используя фрагменты одноцепочечной РНК (оцРНК), такие как микроРНК (миРНК) или двухцепочечные малые интерферирующие РНК (миРНК), комплекс действует как ключевой инструмент регуляции генов. [2] Одноцепочечная РНК действует как матрица для RISC для распознавания комплементарной информационной РНК (мРНК) транскрипта . После обнаружения один из белков RISC, Argonaute , активирует и расщепляет мРНК. Этот процесс называется РНК-интерференцией (РНКи) и обнаружен у многих эукариот ; это ключевой процесс защиты от вирусных инфекций , поскольку он запускается наличием двухцепочечной РНК (дцРНК). [3] [4] [1]
Открытие
[ редактировать ]Биохимическую и идентификацию RISC провели Грегори Хэннон его коллеги из лаборатории Колд-Спринг-Харбор . [5] Это произошло всего через пару лет после открытия РНК-интерференции в 1998 году Эндрю Файром и Крейгом Мелло , которые разделили Нобелевскую премию 2006 года по физиологии и медицине . [3]

Хэннон и его коллеги попытались идентифицировать механизмы РНКи, участвующие в подавлении генов с помощью дцРНК в клетках дрозофилы . дрозофилы Клетки S2 трансфицировали экспрессии вектором lacZ экспрессии для количественной оценки гена с активностью β-галактозидазы . Результаты показали, что совместная трансфекция дсРНК lacZ значительно снижает активность β-галактозидазы по сравнению с контрольной дсРНК. Следовательно, дцРНК контролируют экспрессию генов посредством комплементарности последовательностей .
Затем клетки S2 трансфицировали дрозофилы дсРНК циклина E . Циклин E является важным геном для клеточного цикла перехода в S-фазу . ДцРНК циклина E останавливала клеточный цикл на ( фазе G1 перед фазой S). Следовательно, RNAi может воздействовать на эндогенные гены.
Кроме того, дцРНК циклина E только уменьшала РНК циклина E - аналогичный результат был также показан при использовании дцРНК, соответствующей циклину А , который действует в фазах S, G 2 и M клеточного цикла. Это демонстрирует характерную особенность РНКи: пониженные уровни мРНК соответствуют уровням добавленной дцРНК.
Чтобы проверить, было ли обнаруженное ими снижение уровней мРНК результатом прямого воздействия на мРНК (как предполагают данные других систем), клетки S2 дрозофилы трансфицировали либо дрозофилы дцРНК циклина E , либо дцРНК lacZ , а затем инкубировали с синтетическими мРНК циклина E. или lacZ .
В клетках, трансфицированных дцРНК циклина E, наблюдалась деградация только транскриптов циклина E — транскрипты lacZ были стабильными. И наоборот, клетки, трансфицированные дцРНК lacZ , демонстрировали деградацию только транскриптов lacZ , но не транскриптов циклина E. Их результаты привели Хэннона и его коллег к предположению, что RNAi разрушает целевую мРНК посредством «специфической для последовательности нуклеазной активности». нуклеазу Они назвали фермент RISC. [5] Позже Девананд Саркар и его коллеги Прасанна К. Сантекадур и Бьюнг Квон Ю из Университета Содружества Вирджинии выяснили активность RISC и ее молекулярный механизм в раковых клетках, а также идентифицировали еще один новый компонент RISC, названный AEG-1 [47].
Функция в интерференции РНК
[ редактировать ]
Включение миРНК/миРНК
[ редактировать ]РНКаза III Dicer является важным членом RISC, который инициирует процесс РНК-интерференции путем производства двухцепочечной миРНК или одноцепочечной миРНК. Ферментативное расщепление дцРНК внутри клетки приводит к образованию коротких фрагментов миРНК длиной 21-23 нуклеотида с двухнуклеотидным 3'- выступающим выступом. [6] [7] Dicer также обрабатывает пре-миРНК, которая образует структуру шпильки для имитации дцРНК, аналогичным образом. Фрагменты дцРНК загружаются в RISC, причем каждая цепь имеет различную судьбу, основанную на явлении правила асимметрии: выбор одной цепи в качестве направляющей цепи по сравнению с другой основан на термодинамической стабильности. [8] [9] [10] [11] Вновь созданные микроРНК или миРНК действуют как одноцепочечные направляющие последовательности для RISC, направляя мРНК для деградации. [12] [13]
- Нить с менее термодинамически стабильным 5'-концом отбирается белком Argonaute и интегрируется в RISC. [11] [14] Эта цепь известна как направляющая цепь и нацелена на деградацию мРНК.
- Другая цепь, известная как пассажирская цепь, разрушается RISC. [15]

Генная регуляция
[ редактировать ]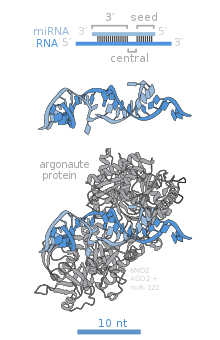
Основные белки RISC, Ago2, SND1 и AEG-1 вносят решающий вклад в функцию подавления генов комплекса. [16]
RISC использует направляющую цепь микроРНК или миРНК для нацеливания на комплементарные 3'-нетранслируемые области (3'UTR) транскриптов мРНК посредством спаривания оснований Уотсона-Крика , что позволяет ему регулировать экспрессию генов транскрипта мРНК несколькими способами. [17] [1]
деградация мРНК
[ редактировать ]Наиболее изученной функцией RISC является деградация целевой мРНК, которая снижает уровни транскриптов, доступных для трансляции рибосомами . Эндонуклеолитическое расщепление мРНК, комплементарной направляющей цепи RISC, белком Argonaute является ключом к инициации RNAi. [18] Для деградации мРНК необходимы два основных требования:
- почти идеальное комплементарное совпадение между направляющей цепью и последовательностью целевой мРНК, и
- каталитически активный белок Argonaute, называемый «срезом», расщепляющий целевую мРНК. [1]
Существует два основных пути деградации мРНК после того, как произошло расщепление. мРНК Оба инициируются путем деградации поли(А)-хвоста , что приводит к удалению 5'-кэпа мРНК.
- Деградация транскрипта с 5' на 3' происходит под действием экзонуклеазы XRN1 в цитоплазматических тельцах, называемых P-тельцами . [19]
- Деградация транскрипта с 3' на 5' осуществляется экзосомой и комплексом Ski . [18]
Трансляционная репрессия
[ редактировать ]RISC может модулировать загрузку рибосом и дополнительных факторов трансляции, экспрессию подавляя связанного транскрипта мРНК. Репрессия трансляции требует лишь частичного совпадения последовательностей между направляющей цепью и целевой мРНК. [1]
На начальном этапе перевод можно регулировать следующим образом:
- предотвращение связывания эукариотического фактора инициации трансляции (eIF) с 5'-кэпом . Было отмечено, что RISC может деаденилировать 3'- поли(А)-хвост , что может способствовать репрессии через 5'-кэп. [2] [17]
- предотвращение связывания рибосомальной субъединицы 60S с мРНК может подавлять трансляцию. [20]
Трансляцию можно регулировать на этапах после инициации путем:
- деградация пептидов,
- содействие преждевременному прекращению трансляции рибосом, [21] или,
- замедление удлинения. [22]
До сих пор ведутся предположения о том, являются ли репрессии трансляции посредством инициации и постинициации взаимоисключающими.
Образование гетерохроматина
[ редактировать ]Некоторые RISC способны напрямую воздействовать на геном , рекрутируя гистоновые метилтрансферазы для формирования гетерохроматина гена в локусе , подавляя ген. Эти RISC принимают форму РНК-индуцированного комплекса подавления транскрипции (RITS). Наиболее изученный пример – дрожжи RITS. [1] [23] [24]
Было показано, что RITS направляет образование гетерохроматина на центромерах посредством распознавания центромерных повторов. Посредством спаривания оснований миРНК (направляющей цепи) с целевыми последовательностями хроматина можно привлечь ферменты, модифицирующие гистоны. [25]
Механизм не совсем понятен; однако RITS разрушают возникающие транскрипты мРНК. Было высказано предположение, что этот механизм действует как «самоусиливающаяся петля обратной связи », поскольку деградированные возникающие транскрипты используются РНК-зависимой РНК-полимеразой (RdRp) для генерации большего количества миРНК. [26]
У Schizosaccharomyces pombe и Arabidopsis процессинг мишеней дцРНК в миРНК с помощью РНКаз Dicer может инициировать путь молчания генов путем образования гетерохроматина. Белок Argonaute, известный как AGO4, взаимодействует с небольшими РНК, которые определяют гетерохроматические последовательности. Гистонметилтрансфераза (HMT ) , H3K9 , метилирует гистон H3 и рекрутирует хромодоменные белки в сайты метилирования. Метилирование ДНК поддерживает молчание генов, поскольку последовательности гетерохроматина могут устанавливаться или распространяться. [27]
удаление ДНК
[ редактировать ]siRNA, генерируемая RISCs, по-видимому, играет роль в деградации ДНК во время соматических макронуклеусов развития у инфузорий рода Tetrahymena . Это похоже на эпигенетический контроль образования гетерохроматина и подразумевается как защита от вторжения генетических элементов. [27]
Подобно образованию гетерохроматина у S. pombe и Arabidopsis , белок Tetrahymena, относящийся к семейству Argonaute, Twi1p, катализирует удаление из ДНК целевых последовательностей, известных как последовательности внутреннего элиминирования (IES). С помощью метилтрансфераз и хромодоменных белков IES гетерохроматизируются и удаляются из ДНК. [27]
RISC-ассоциированные белки
[ редактировать ]Полная структура RISC до сих пор не определена. Во многих исследованиях сообщалось о различных размерах и компонентах RISC, но не совсем ясно, связано ли это с наличием ряда комплексов RISC или с разными источниками, которые используют разные исследования. [28]
| Сложный | Источник | Известные/очевидные компоненты | Предполагаемый размер | Очевидная функция пути РНКи |
|---|---|---|---|---|
| Дкр2-Р2Д2 [29] | D. melanogaster Клетки S2 | Дкр2 , Р2Д2 | ~250 кДа | Процессинг дсРНК, связывание миРНК |
| РЛК (А) [30] [31] | D. melanogaster Эмбрионы | Дкр2, Р2Д2 | Нет. | Процессинг дсРНК, связывание миРНК, предшественник RISC |
| Голо-РИСК [30] [31] | D. melanogaster Эмбрионы | Назад 2 , Дкр1, Дкр2, Фмр1 / Фхр , Р2Д2, Цн , Виг | ~80С | Связывание и расщепление целевой РНК |
| РИСК [5] [32] [33] [34] | D. melanogaster Клетки S2 | Назад2, Fmr1/Fxr, Цн, Виг | ~500 кДа | Связывание и расщепление целевой РНК |
| РИСК [35] | D. melanogaster Клетки S2 | Назад2 | ~140 кДа | Связывание и расщепление целевой РНК |
| Fmr1-ассоциированный комплекс [36] | D. melanogaster Клетки S2 | L5 , L11 , 5S рРНК , Fmr1/Fxr, Ago2, Dmp68 | Нет. | Возможное связывание и расщепление целевой РНК |
| Минимальный RISC [37] [38] [39] [40] | HeLa клетки | eIF2C1 (Ago1) или eIF2C2 (Ago2) | ~160 кДа | Связывание и расщепление целевой РНК |
| миРНП [41] [42] | клетки HeLa | eIF2C2 (назад2), Близнец3 , Близнец4 | ~550 кДа | Ассоциация микроРНК, связывание и расщепление целевой РНК |
Назад, Аргонавт; Дкр, Дайсер; Dmp68 — D. melanogaster ортолог униндазы РНК р68 млекопитающих ; eIF2C1 – эукариотический фактор инициации трансляции 2C1; eIF2C2 – эукариотический фактор инициации трансляции 2C2; Fmr1/Fxr — ортолог D. melanogaster белка хрупкой-X умственной отсталости; миРНП – комплекс миРНК-белок; НР, не сообщается; Цн – Тудор-стафилококковая нуклеаза; Vig, интронный ген vasa.

Тем не менее, очевидно, что белки Argonaute присутствуют и необходимы для функционирования. Кроме того, есть сведения о некоторых ключевых белках (помимо Argonaute) в составе комплекса, которые позволяют RISC выполнять свою функцию.
Аргонавтовые белки
[ редактировать ]Белки аргонавты — это семейство белков, обнаруженных у прокариот и эукариот. Их функция у прокариот неизвестна, но у эукариот они отвечают за РНКи. [43] У человека-аргонавтов восемь членов семейства, из которых только Argonaute 2 участвует исключительно в целевом расщеплении РНК в RISC. [40]

RISC-загрузочный комплекс
[ редактировать ]Комплекс загрузки RISC (RLC) представляет собой важную структуру, необходимую для загрузки фрагментов дцРНК в RISC для нацеливания на мРНК. RLC состоит из дайсера, РНК-связывающего белка трансактивирующего ответа ( TRBP ) и Argonaute 2.
- Dicer РНКазы III представляет собой эндонуклеазу , которая генерирует загружаемые фрагменты дцРНК, которые направляют РНКи.
- TRBP представляет собой белок с тремя двухцепочечными РНК-связывающими [белковый домен|домены].
- Аргонавт 2 представляет собой РНКазу и является каталитическим центром RISC.
Dicer связывается с TRBP и Argonaute 2, чтобы облегчить перенос фрагментов дцРНК, генерируемых Dicer, в Argonaute 2. [44] [45]
Более поздние исследования показали, что человеческая РНК-хеликаза А может помочь облегчить RLC. [46]
Другие белки
[ редактировать ]Недавно выявленными членами RISC являются SND1 и MTDH . [47] SND1 и MTDH являются онкогенами и регулируют экспрессию различных генов. [48]
| Белок | Виды, в которых встречается белок |
|---|---|
| Дкр1 [30] | Д. меланогастер |
| Дкр2 [29] [30] [31] | Д. меланогастер |
| Р2Д2 [30] [31] | Д. меланогастер |
| Назад2 [30] [32] [35] [36] | Д. меланогастер |
| Дмп68 [36] | Д. меланогастер |
| Фмр1/Фкр [30] [33] [36] | Д. меланогастер |
| ЦН [30] [34] | Д. меланогастер |
| Виг [30] [33] | Д. меланогастер |
| Полирибосомы , компоненты рибосом [5] [30] [32] [36] [49] | D. melanogaster , Т. brucei |
| eIF2C1 (1 августа) [37] | Х. сапиенс |
| eIF2C2 (Назад2) [37] [38] [40] [42] | Х. сапиенс |
| Близнецы3 [41] [42] | Х. сапиенс |
| Близнецы4 [41] [42] | Х. сапиенс |
Назад, Аргонавт; Дкр, Дайсер; Dmp68 — D. melanogaster ортолог униндазы РНК р68 млекопитающих ; eIF2C1 – эукариотический фактор инициации трансляции 2C1; eIF2C2 – эукариотический фактор инициации трансляции 2C2; Fmr1/Fxr — ортолог D. melanogaster белка хрупкой-X умственной отсталости; Цн – Тудор-стафилококковая нуклеаза; Vig, интронный ген vasa.
Связывание мРНК
[ редактировать ]
Пока неясно, как активированный комплекс RISC находит мишени мРНК в клетке, хотя было показано, что этот процесс может происходить и в ситуациях, не связанных с продолжающейся трансляцией белка с мРНК. [50]
Эндогенно экспрессируемая микроРНК у многоклеточных животных обычно не является полностью комплементарной большому числу генов и, таким образом, модулирует экспрессию посредством репрессии трансляции. [51] [52] Однако у растений этот процесс имеет гораздо большую специфичность в отношении мРНК-мишени, и обычно каждая микроРНК связывается только с одной мРНК. Более высокая специфичность означает, что более вероятна деградация мРНК. [53]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж Пратт А.Дж., Макрей И.Дж. (2009). «Комплекс молчания, индуцированный РНК: универсальная машина для подавления генов» . Журнал биологической химии . 284 (27): 17897–17901. дои : 10.1074/jbc.R900012200 . ПМК 2709356 . ПМИД 19342379 .
- ^ Jump up to: а б Филипович В., Бхаттачария С.Н., Соненбер Н. (2008). «Механизмы посттранскрипционной регуляции микроРНК: ответы видны?» . Обзоры природы Генетика . 9 (2): 102–114. дои : 10.1038/nrg2290 . ПМИД 18197166 . S2CID 11824239 .
- ^ Jump up to: а б Файер А, Сюй С, Монтгомери МК, Костас С.А., Драйвер С.Е., Мелло CC (1998). «Мощное и специфическое генетическое вмешательство двухцепочечной РНК в Caenorhabditis elegans » . Природа . 391 (6669): 806–811. Бибкод : 1998Natur.391..806F . дои : 10.1038/35888 . ПМИД 9486653 . S2CID 4355692 .
- ^ Уотсон, Джеймс Д. (2008). Молекулярная биология гена . Сан-Франциско, Калифорния: Лабораторное издательство Колд-Спринг-Харбор. стр. 641–648. ISBN 978-0-8053-9592-1 .
- ^ Jump up to: а б с д Хаммонд С.М., Бернштейн Э. , Бич Д., Хэннон Г.Дж. (2000). «РНК-направленная нуклеаза опосредует посттранскрипционное молчание генов в клетках дрозофилы ». Природа . 404 (6775): 293–296. Бибкод : 2000Natur.404..293H . дои : 10.1038/35005107 . ПМИД 10749213 . S2CID 9091863 .
- ^ Замор П.Д., Тушл Т., Шарп П.А., Бартель Д.П. (2000). «РНКи: двухцепочечная РНК направляет АТФ-зависимое расщепление мРНК с интервалами от 21 до 23 нуклеотидов» . Клетка . 101 (1): 25–33. дои : 10.1016/S0092-8674(00)80620-0 . ПМИД 10778853 .
- ^ Вермюлен А., Белен Л., Рейнольдс А., Вольфсон А., Маршалл В., Карпилов Дж., Хворова А. (2005). «Вклад структуры дцРНК в специфичность и эффективность Dicer» . РНК . 11 (5): 674–682. дои : 10.1261/rna.7272305 . ПМК 1370754 . ПМИД 15811921 .
- ^ Хутвагнер, Дьёрдь (2005). «Асимметрия малых РНК в РНКи: функция сборки RISC и регуляции генов» . Письма ФЭБС . 579 (26): 5850–5857. doi : 10.1016/j.febslet.2005.08.071 . hdl : 10453/15313 . ISSN 1873-3468 . ПМИД 16199039 .
- ^ Шварц Д.С., Хутвагнер Г., Ду Т, Сюй З, Аронин Н, Замор П.Д. (2003). «Асимметрия сборки ферментного комплекса РНКи» . Клетка . 115 (2): 199–208. дои : 10.1016/S0092-8674(03)00759-1 . ПМИД 14567917 .
- ^ Хворова А, Рейнольдс А, Джаясена С.Д. (2003). «Функциональные миРНК и микроРНК демонстрируют смещение цепи» . Клетка . 115 (2): 209–216. дои : 10.1016/S0092-8674(03)00801-8 . ПМИД 14567918 . S2CID 2500175 .
- ^ Jump up to: а б Сиоми Х, Сиоми MC (2009). «На пути к чтению кода РНК-интерференции». Природа . 457 (7228): 396–404. Бибкод : 2009Natur.457..396S . дои : 10.1038/nature07754 . ПМИД 19158785 . S2CID 205215974 .
- ^ Преалл, Джонатан Б.; Сонтхаймер, Эрик Дж. (18 ноября 2005 г.). «RNAi: RISC загружается» . Клетка . 123 (4): 543–545. дои : 10.1016/j.cell.2005.11.006 . ISSN 0092-8674 . ПМИД 16286001 .
- ^ «Обзор РНК-интерференции | Abcam» . www.abcam.com . Проверено 7 марта 2021 г.
- ^ Преалл, Джонатан Б.; Он, Чжэнъин; Горра, Джеффри М.; Сонтхаймер, Эрик Дж. (7 марта 2006 г.). «Выбор короткой цепи интерферирующей РНК не зависит от полярности процессинга дцРНК во время РНКи у дрозофилы» . Современная биология . 16 (5): 530–535. Бибкод : 2006CBio...16..530P . дои : 10.1016/j.cub.2006.01.061 . ISSN 0960-9822 . ПМИД 16527750 .
- ^ Грегори Р.И., Чендримада Т.П., Куч Н., Шихаттар Р. (2005). «Человеческий RISC сочетает биогенез микроРНК и посттранскрипционное молчание генов» . Клетка . 123 (4): 631–640. дои : 10.1016/j.cell.2005.10.022 . ПМИД 16271387 .
- ^ Сантекадур, Прасанна К.; Кумар, Дивья П. (01.06.2020). «Сборка RISC и посттранскрипционная регуляция генов при гепатоцеллюлярной карциноме» . Гены и болезни . 7 (2): 199–204. дои : 10.1016/j.gendis.2019.09.009 . ISSN 2352-3042 . ПМК 7083748 . ПМИД 32215289 .
- ^ Jump up to: а б Вакияма М., Такимото К., Охара О., Ёкояма С. (2007). «Let-7-микроРНК-опосредованное деаденилирование мРНК и репрессия трансляции в бесклеточной системе млекопитающих» . Гены и развитие . 21 (15): 1857–1862. дои : 10.1101/gad.1566707 . ПМК 1935024 . ПМИД 17671087 .
- ^ Jump up to: а б ОРБАН, ТАМАС И.; ИЗАУРРАЛДЕ, ИФА (апрель 2005 г.). «Распад мРНК, на которые нацелен RISC, требует XRN1, комплекса Ski и экзосомы» . РНК . 11 (4): 459–469. дои : 10.1261/rna.7231505 . ISSN 1355-8382 . ПМК 1370735 . ПМИД 15703439 .
- ^ Сен Г.Л., Блау Х.М. (2005). «Argonaute2/RISC находится в местах распада мРНК млекопитающих, известных как цитоплазматические тельца». Природная клеточная биология . 7 (6): 633–636. дои : 10.1038/ncb1265 . ПМИД 15908945 . S2CID 6085169 .
- ^ Чендримада Т.П., Финн К.Дж., Джи Икс, Байлат Д., Грегори Р.И., Либхабер С.А., Паскинелли А.Е., Шихаттар Р. (2007). «Замалчивание микроРНК посредством рекрутирования RISC eIF6». Природа . 447 (7146): 823–828. Бибкод : 2007Natur.447..823C . дои : 10.1038/nature05841 . ПМИД 17507929 . S2CID 4413327 .
- ^ Петерсен К.П., Бордело М.Э., Пеллетье Дж., Шарп П.А. (2006). «Короткие РНК подавляют трансляцию после инициации в клетках млекопитающих» . Молекулярная клетка . 21 (4): 533–542. doi : 10.1016/j.molcel.2006.01.031 . ПМИД 16483934 .
- ^ Марони П.А., Ю Ю., Фишер Дж., Нильсен Т.В. (2006). «Доказательства того, что микроРНК связаны с трансляцией информационных РНК в клетках человека». Структурная и молекулярная биология природы . 13 (12): 1102–1107. дои : 10.1038/nsmb1174 . ПМИД 17128271 . S2CID 19106463 .
- ^ Вердель А., Цзя С., Гербер С., Сугияма Т., Гиги С., Гревал С.И., Моазед Д. (2004). «РНКи-опосредованное нацеливание на гетерохроматин комплексом RITS» . Наука . 303 (5658): 672–676. Бибкод : 2004Sci...303..672V . дои : 10.1126/science.1093686 . ПМК 3244756 . ПМИД 14704433 .
- ^ Вердель А., Цзя С., Гербер С., Сугияма Т., Гиги С., Гревал С.И., Моазед Д. (2004). «RITS действует в цис-цис-режиме , способствуя транскрипции, опосредованной интерференцией РНК, и посттранскрипционному молчанию» . Природная генетика . 36 (11): 1174–1180. дои : 10.1038/ng1452 . ПМИД 15475954 .
- ^ Симада, Юкико; Мон, Фабио; Бюлер, Марк (1 декабря 2016 г.). «Комплекс подавления транскрипции, индуцированный РНК, нацелен исключительно на хроматин посредством взаимодействия с возникающими транскриптами» . Гены и развитие . 30 (23): 2571–2580. дои : 10.1101/gad.292599.116 . ISSN 0890-9369 . ПМК 5204350 . ПМИД 27941123 .
- ^ Сугияма Т., Кэм Х., Вердель А., Моазед Д., Гревал С.И. (2005). «РНК-зависимая РНК-полимераза является важным компонентом самоподдерживающейся петли, соединяющей сборку гетерохроматина с производством миРНК» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (1): 152–157. дои : 10.1073/pnas.0407641102 . ПМК 544066 . ПМИД 15615848 .
- ^ Jump up to: а б с Мочизуки К., Горовский М.А. (2004). «Маленькие РНК в расположении генома тетрахимены ». Текущее мнение в области генетики и развития . 14 (2): 181–187. дои : 10.1016/j.где.2004.01.004 . ПМИД 15196465 .
- ^ Jump up to: а б с Зонтхаймер Э.Дж. (2005). «Сборка и функция комплексов молчания РНК». Nature Reviews Молекулярно-клеточная биология . 6 (2): 127–138. дои : 10.1038/nrm1568 . ПМИД 15654322 . S2CID 27294007 .
- ^ Jump up to: а б Лю Кью, Рэнд Т.А., Калидас С., Ду Ф, Ким Х.Э., Смит Д.П., Ван Х (2003). «R2D2, мост между инициирующим и эффекторным этапами пути RNAi дрозофилы ». Наука . 301 (5641): 1921–1925. Бибкод : 2003Sci...301.1921L . дои : 10.1126/science.1088710 . ПМИД 14512631 . S2CID 41436233 .
- ^ Jump up to: а б с д и ж г час я дж Фам Дж.В., Пеллио Дж.Л., Ли Ю.С., Картью Р.В., Сонтхаймер Э.Дж. (2004). «Зависимый от Dicer-2 80S комплекс расщепляет целевые мРНК во время РНКи у дрозофилы » . Клетка . 117 (1): 83–94. дои : 10.1016/S0092-8674(04)00258-2 . ПМИД 15066284 .
- ^ Jump up to: а б с д Томари Ю, Ду Т, Хейли Б, Шварц Д.С., Беннетт Р., Кук Х.А., Коппетш Б.С., Теуркауф В.Е., Замор П.Д. (2004). «Дефекты сборки RISC в RNAi дрозофилы мутантном армитаже » . Клетка . 116 (6): 831–841. дои : 10.1016/S0092-8674(04)00218-1 . ПМИД 15035985 .
- ^ Jump up to: а б с Хаммонд С.М., Бетчер С., Коди А.А., Кобаяши Р., Хэннон Г.Дж. (2001). «Argonaute2, связь между генетическим и биохимическим анализом РНКи». Наука . 293 (5532): 1146–1150. дои : 10.1126/science.1064023 . ПМИД 11498593 . S2CID 5271290 .
- ^ Jump up to: а б с Коди А.А., Майерс М., Хэннон Дж.Дж., Хаммонд С.М. (2002). «Хрупкий Х-родственный белок и VIG связываются с механизмом РНК-интерференции» . Гены и развитие . 16 (19): 2491–2496. дои : 10.1101/gad.1025202 . ПМК 187452 . ПМИД 12368260 .
- ^ Jump up to: а б Коди А.А., Кеттинг Р.Ф., Хаммонд С.М., Денли А.М., Батхорн А.М., Топс Б.Б., Сильва Дж.М., Майерс М.М., Хэннон Г.Дж., Пластерк Р.Х. (2003). «Гомолог микрококковой нуклеазы в эффекторных комплексах РНКи». Природа . 425 (6956): 411–414. Бибкод : 2003Natur.425..411C . дои : 10.1038/nature01956 . ПМИД 14508492 . S2CID 4410688 .
- ^ Jump up to: а б Рэнд Т.А., Гинальский К., Гришин Н.В., Ван Х (2004). «Биохимическая идентификация Argonaute 2 как единственного белка, необходимого для активности комплекса молчания, индуцированного РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (40): 14385–14389. Бибкод : 2004PNAS..10114385R . дои : 10.1073/pnas.0405913101 . ПМК 521941 . ПМИД 15452342 .
- ^ Jump up to: а б с д и Исидзука А, Сиоми MC, Сиоми Х (2002). « Хрупкий X-белок дрозофилы взаимодействует с компонентами РНКи и рибосомальными белками» . Гены и развитие . 16 (19): 2497–2508. дои : 10.1101/gad.1022002 . ПМК 187455 . ПМИД 12368261 .
- ^ Jump up to: а б с Мартинес Дж., Патканиовска А., Урлауб Х., Лурманн Р., Тушль Т. (2002). «Одноцепочечные антисмысловые миРНК направляют расщепление целевой РНК в РНКи». Клетка . 110 (5): 563–574. дои : 10.1016/S0092-8674(02)00908-X . hdl : 11858/00-001M-0000-0012-F2FD-2 . ПМИД 12230974 . S2CID 10616773 .
- ^ Jump up to: а б Лю Дж., Кармелл М.А., Ривас Ф.В., Марсден К.Г., Томсон Дж.М., Сонг Дж.Дж., Хаммонд С.М., Джошуа-Тор Л., Хэннон Дж.Дж. (2004). «Argonaute2 — каталитический двигатель РНКи млекопитающих» . Наука . 305 (5689): 1437–1441. Бибкод : 2004Sci...305.1437L . дои : 10.1126/science.1102513 . ПМИД 15284456 . S2CID 2778088 .
- ^ Мартинес Дж., Тушл Т. (2004). «RISC представляет собой эндонуклеазу РНК, продуцирующую 5'-фосфомоноэфир» . Гены и развитие . 18 (9): 975–980. дои : 10.1101/gad.1187904 . ПМК 406288 . ПМИД 15105377 .
- ^ Jump up to: а б с Мейстер Г., Ландталер М., Патканёвска А., Дорсетт Ю., Тенг Г., Тушль Т. (2004). «Человеческий Argonaute2 опосредует расщепление РНК, на которое направлены микроРНК и миРНК» . Молекулярная клетка . 15 (2): 1403–1408. doi : 10.1016/j.molcel.2004.07.007 . ПМИД 15260970 .
- ^ Jump up to: а б с Мурелатос З., Дости Дж., Паушкин С., Шарма А., Шарру Б., Абель Л., Раппсилбер Дж. , Манн М., Дрейфус Г. (2002). «МиРНП: новый класс рибонуклеопротеинов, содержащий многочисленные микроРНК» . Гены и развитие . 16 (6): 720–728. дои : 10.1101/gad.974702 . ПМК 155365 . ПМИД 11914277 .
- ^ Jump up to: а б с д Хутвангнер Г., Замор П.Д. (2002). «МикроРНК в многооборотном ферментном комплексе РНКи». Наука . 297 (5589): 2056–2060. Бибкод : 2002Sci...297.2056H . дои : 10.1126/science.1073827 . ПМИД 12154197 . S2CID 16969059 .
- ^ Холл ТМ (2005). «Структура и функции белков аргонавтов» . Клетка . 13 (10): 1403–1408. дои : 10.1016/j.str.2005.08.005 . ПМИД 16216572 .
- ^ Чендримада Т.П., Грегори Р.И., Кумарасвами Э., Норман Дж., Куч Н., Нисикура К., Шикхаттер Р. (2005). «TRBP рекрутирует комплекс Dicer к Ago2 для процессинга микроРНК и подавления генов» . Природа . 436 (7051): 740–744. Бибкод : 2005Natur.436..740C . дои : 10.1038/nature03868 . ПМЦ 2944926 . ПМИД 15973356 .
- ^ Ван Х.В., Ноланд С., Сиричадилок Б., Тейлор Д.В., Ма Э., Фельдерер К., Дудна Дж.А., Ногалес Е (2009). «Структурные данные об обработке РНК комплексом RISC-загрузки человека» . Структурная и молекулярная биология природы . 16 (11): 1148–1153. дои : 10.1038/nsmb.1673 . ПМЦ 2845538 . ПМИД 19820710 .
- ^ Фу Ц, Юань Я. (2013). «Структурное понимание сборки RISC, чему способствуют дцРНК-связывающие домены спиралей человеческой РНК A (DHX9)» . Исследования нуклеиновых кислот . 41 (5): 3457–3470. дои : 10.1093/нар/gkt042 . ПМЦ 3597700 . ПМИД 23361462 .
- ^ Ю Б.К., Сантекадур П.К., Гредлер Р., Чен Д., Эмдад Л., Бхутиа С., Паннелл Л., Фишер П.Б., Саркар Д. (2011). «Повышенная активность РНК-индуцированного комплекса молчания (RISC) способствует развитию гепатоцеллюлярной карциномы» . Гепатология . 53 (5): 1538–1548. дои : 10.1002/hep.24216 . ПМК 3081619 . ПМИД 21520169 .
- ^ Ю Б.К., Эмдад Л., Ли С.Г., Су З., Сантекадур П., Чен Д., Гредлер Р., Фишер П.Б., Саркар Д. (2011). «Ген повышенного уровня астроцитов (AEG-1): многофункциональный регулятор нормальной и аномальной физиологии» . Фармакология и терапия . 130 (1): 1–8. doi : 10.1016/j.pharmthera.2011.01.008 . ПМК 3043119 . ПМИД 21256156 .
- ^ Джикенг А, Ши Х, Чуди С, Шен С, Уллу Э (2003). «Рибонуклеопротеин siRNA обнаружен связанный с полирибосомами Trypanosoma brucei » . РНК . 9 (7): 802–808. дои : 10.1261/rna.5270203 . ПМК 1370447 . ПМИД 12810914 .
- ^ Сен ГЛ, Верман ТС, Блау ХМ (2005). «Трансляция мРНК не является обязательным условием для расщепления мРНК, опосредованного малой интерферирующей РНК». Дифференциация . 73 (6): 287–293. дои : 10.1111/j.1432-0436.2005.00029.x . ПМИД 16138829 .
- ^ Саумет А, Леселье CH (2006). «Антивирусное подавление РНК: мы похожи на растения?» . Ретровирусология . 3 :3. дои : 10.1186/1742-4690-3-3 . ПМЦ 1363733 . ПМИД 16409629 .
- ^ Бартель Д.П. (2009). «МикроРНК: целевое распознавание и регуляторные функции» . Клетка . 136 (2): 215–233. дои : 10.1016/j.cell.2009.01.002 . ПМЦ 3794896 . ПМИД 19167326 .
- ^ Джонс-Роудс М.В., Бартель Д.П., Бартель Б. (2006). «МикроРНК и их регуляторная роль в растениях». Ежегодный обзор биологии растений . 57 : 19–53. doi : 10.1146/annurev.arplant.57.032905.105218 . ПМИД 16669754 .
Дальнейшее чтение
[ редактировать ]- Сонтхаймер, Э.Дж. (2005). «Сборка и функция комплексов молчания РНК». Nature Reviews Молекулярно-клеточная биология . 6 (2): 127–138. дои : 10.1038/nrm1568 . ПМИД 15654322 . S2CID 27294007 .
- Фу Ц, Юань Я. (март 2013 г.). «Структурное понимание сборки RISC, чему способствуют дцРНК-связывающие домены человеческой РНК-хеликазы A (DHX9)» . Исследования нуклеиновых кислот . 41 (5): 3457–70. дои : 10.1093/нар/gkt042 . ПМЦ 3597700 . ПМИД 23361462 .
- Шварц Д.С., Томари Й., Замор П.Д. (2004). «Комплекс молчания, индуцированный РНК, представляет собой Mg 2+ -зависимая эндонуклеаза» . Current Biology . 14 (9): 787–91. Bibcode : 2004CBio...14..787S . doi : 10.1016/j.cub.2004.03.008 . PMID 15120070 .
Внешние ссылки
[ редактировать ]- РНК-индуцированный + глушитель + комплекс в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)