Аргонавт
Семейство Argonaute белков , впервые обнаруженное из-за его эволюционно консервативной функции стволовых клеток, [1] играет центральную роль в процессах молчания РНК, являясь важным компонентом комплекса РНК-индуцированного молчания (RISC). RISC отвечает за феномен молчания генов, известный как РНК-интерференция (RNAi) . [2] Белки Argonaute связывают различные классы малых некодирующих РНК , включая микроРНК (миРНК), малые интерферирующие РНК (миРНК) и Piwi-взаимодействующие РНК (пиРНК). Малые РНК направляют белки Argonaute к их конкретным мишеням посредством комплементарности последовательностей (спаривания оснований), что затем приводит к расщеплению мРНК, ингибированию трансляции и/или инициированию распада мРНК. [3]
Название этого семейства белков происходит от мутантного фенотипа, возникшего в результате мутации AGO1 у Arabidopsis thaliana , которую сравнили Bohmert et al. к появлению пелагического осьминога Argonauta argo . [4]
| Домен Аргонавта Пиви | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | Пиви | ||
| Пфам | PF02171 | ||
| ИнтерПро | ИПР003165 | ||
| PROSITE | ПС50822 | ||
| CDD | cd02826 | ||
| |||
| Домен Аргонавта Паса | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | Мир | ||
| Пфам | ПФ12212 | ||
| ИнтерПро | ИПР021103 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | b.34.14.1 / ОБЛАСТЬ ПРИМЕНЕНИЯ / СУПФАМ | ||
| |||

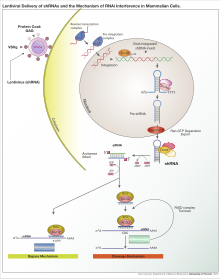
РНК-интерференция
[ редактировать ]РНК-интерференция (РНКи) — это биологический процесс, при котором молекулы РНК ингибируют экспрессию генов либо путем разрушения определенных молекул мРНК, либо путем подавления трансляции. [5] РНКи играет важную роль в защите клеток от паразитных нуклеотидных последовательностей. [ нужна ссылка ] . У эукариот, включая животных, РНКи инициируется ферментом Dicer . Dicer расщепляет длинные молекулы двухцепочечной РНК (дцРНК, часто встречающаяся в вирусах и небольших интерферирующих РНК ) на короткие двухцепочечные фрагменты, состоящие примерно из 20 нуклеотидов siRNA. Затем дцРНК разделяется на две одноцепочечные РНК (оцРНК) — пассажирскую цепь и направляющую цепь. Впоследствии пассажирская цепь разрушается, а направляющая цепь включается в РНК-индуцированный комплекс молчания (RISC). Наиболее хорошо изученным результатом РНКи является посттранскрипционное молчание генов, которое происходит, когда направляющая цепь спаривается с комплементарной последовательностью в молекуле информационной РНК и индуцирует расщепление с помощью Argonaute, которое лежит в ядре РНК-индуцированного комплекса молчания.
Белки Argonaute являются активной частью РНК-индуцированного комплекса молчания, расщепляя целевую цепь мРНК, комплементарную связанной с ними миРНК. [6] Теоретически дайсер производит короткие двухцепочечные фрагменты, поэтому должны быть также произведены две функциональные одноцепочечные миРНК. Но здесь только одна из двух одноцепочечных РНК будет использоваться для образования пары оснований с целевой мРНК . Он известен как направляющая цепь, включенная в белок Argonaute и приводящая к молчанию генов. Другая одноцепочечная цепь-пассажир деградирует во время процесса РНК-индуцированного комплекса молчания. [7]
Как только Argonaute связывается с малой РНК, ферментативная активность, обеспечиваемая доменом PIWI, расщепляет только пассажирскую цепь малой интерферирующей РНК. Разделение нитей РНК и включение в белок Argonaute регулируются силой взаимодействия водородных связей на 5'-концах дуплекса РНК, известной как правило асимметрии. Также степень комплементарности между двумя нитями промежуточного дуплекса РНК определяет, как микроРНК сортируются на различные типы белков Argonaute.
У животных Argonaute, связанный с миРНК, связывается с 3'-нетранслируемой областью мРНК и различными способами предотвращает продукцию белков. Привлечение белков Argonaute к целевой мРНК может вызвать деградацию мРНК. Комплекс Argonaute-miRNA также может влиять на образование функциональных рибосом на 5'-конце мРНК. Комплекс здесь конкурирует с факторами инициации трансляции и/или аннулирует сборку рибосомы . Кроме того, комплекс Argonaute-miRNA может регулировать выработку белка путем привлечения клеточных факторов, таких как пептиды или посттрансляционные модифицирующие ферменты, которые ухудшают рост полипептидов. [8]
У растений, как только de novo образуются дуплексы двухцепочечной (дц) РНК с целевой мРНК, неизвестный фермент, подобный РНКазе-III, продуцирует новые миРНК, которые затем загружаются на белки Argonaute, содержащие домены PIWI, в которых отсутствует каталитическая аминокислота. остатки, которые могут индуцировать другой уровень специфического молчания генов.
Функциональные области и механизм
[ редактировать ]Семейство генов Argonaute (AGO) кодирует шесть характерных доменов: N-концевой (N), линкер-1 (L1), PAZ, линкер-2 (L2), Mid и C-концевой домен PIWI . [8]
Домен PAZ назван в честь Drosophila Piwi, Arabidopsis Argonaute-1 и Arabidopsis Zwille (также известного как булавочная головка, позже переименованного в argonaute-10), где домен впервые был признан консервативным. Домен PAZ представляет собой РНК-связывающий модуль, который распознает одноцепочечные 3'-концы миРНК , миРНК и пиРНК независимым от последовательности образом.
PIWI назван в честь белка Piwi дрозофилы. Домен PIWI, структурно напоминающий РНКазу H, необходим для целевого расщепления. Активный центр с триадой аспартат-аспартат-глутамат содержит ион двухвалентного металла, необходимый для катализа. Члены семейства AGO, утратившие эту консервативную особенность в ходе эволюции, лишены активности расщепления. В AGO человека мотив PIWI также опосредует белок-белковое взаимодействие в PIWI-боксе, где он связывается с Dicer в домене РНКазы III. [9]
На границе доменов PIWI и Mid находится 5'-фосфат миРНК, миРНК или пиРНК, функциональность которого имеет важное значение. Внутри Mid находится мотив MC, гомологическая структура, предположительно имитирующая мотив структуры связывания кэпа, обнаруженный в eIF4E. Позже было обнаружено, что мотив MC не участвует в связывании кэпа мРНК. [8]
Члены семьи
[ редактировать ]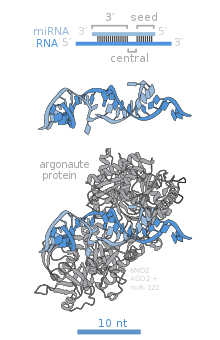
У человека имеется восемь членов семейства AGO, некоторые из которых интенсивно исследуются. Однако, хотя AGO1-4 способны загружать микроРНК, эндонуклеазная активность и, следовательно, RNAi-зависимое подавление генов принадлежит исключительно AGO2. Учитывая консервативность последовательностей доменов PAZ и PIWI во всем семействе, предполагается, что уникальность AGO2 обусловлена либо N-концом, либо спейсерной областью, соединяющей мотивы PAZ и PIWI. [9]
Некоторые представители семейства AGO у растений также привлекают внимание. AGO1 участвует в деградации РНК, связанной с микроРНК, и играет центральную роль в морфогенезе. У некоторых организмов это строго необходимо для эпигенетического молчания. Он регулируется самой микроРНК. AGO4 не участвует в деградации РНК, направленной РНКи, но участвует в метилировании ДНК и других эпигенетических регуляциях посредством пути малых РНК (мРНК). AGO10 участвует в разработке растений. AGO7 имеет функцию, отличную от AGO 1 и 10, и не обнаруживается при подавлении генов, индуцированном трансгенами. Вместо этого это связано со сроками развития растений. [10]
Болезни и терапевтические инструменты
[ редактировать ]Сообщалось, что белки аргонавты связаны с раком. [11] [12] Для заболеваний, которые связаны с селективной или повышенной экспрессией конкретных идентифицированных генов, таких как рак поджелудочной железы, высокая специфичность РНК-интерференции может сделать ее подходящим методом лечения, особенно подходящим для борьбы с раком, связанным с мутированными последовательностями эндогенных генов. Сообщалось, что несколько крошечных некодирующих РНК (микроРНК) связаны с раком человека, например, миР-15а и миР-16а часто удаляются и/или подавляются у пациентов. Несмотря на то, что биологические функции микроРНК до конца не изучены, роль микроРНК в координации клеточной пролиферации и гибели клеток во время развития и метаболизма была раскрыта. Считается, что микроРНК могут направлять негативную или позитивную регуляцию на разных уровнях, что зависит от взаимодействия конкретных микроРНК и целевой пары оснований, а также от кофакторов, которые их распознают. [13]
Поскольку широко известно, что многие вирусы имеют в качестве генетического материала РНК, а не ДНК, и проходят по крайней мере одну стадию своего жизненного цикла, когда они образуют двухцепочечную РНК, РНК-интерференция считается потенциально древним механизмом эволюции. защита организмов от вирусов. Небольшие интерферирующие РНК, продуцируемые Dicer , вызывают специфичное для последовательности посттранскрипционное молчание генов, направляя эндонуклеазу, РНК-индуцированный комплекс молчания (RISC), к мРНК. Этот процесс наблюдался у широкого спектра организмов, таких как гриб Neurospora (у которого он известен как подавление), растения (посттранскрипционное подавление генов) и клетки млекопитающих (RNAi). Если существует полная или почти полная комплементарность последовательностей между малой РНК и мишенью, белковый компонент Argonaute RISC опосредует расщепление целевого транскрипта, механизм включает преимущественно репрессию трансляции. [ нужна ссылка ] .
Биотехнологическое применение прокариотических белков Argonaute
[ редактировать ]В 2016 году группа из Хэбэйского университета науки и технологий сообщила о редактировании генома с использованием прокариотического белка Argonaute из Natronobacterium gregoryi . Однако доказательства применения белков Argonaute в качестве ДНК-ориентированных нуклеаз для редактирования генома были подвергнуты сомнению, а ведущий журнал отозвал это заявление. [14] В 2017 году группа из Университета Иллинойса сообщила об использовании прокариотического белка Argonaute, взятого из Pyrococcus Furiosus (PfAgo), вместе с направляющей ДНК для редактирования ДНК in vitro в качестве искусственных ферментов рестрикции . [15] Искусственные ферменты рестрикции на основе PfAgo также использовались для хранения данных о нативных последовательностях ДНК посредством ферментативного разрыва. [16]
Ссылки
[ редактировать ]- ^ Кокс Д.Н., Чао А., Бейкер Дж., Чанг Л., Цяо Д., Линь Х. (декабрь 1998 г.). «Новый класс эволюционно консервативных генов, определенный piwi, необходим для самообновления стволовых клеток» . Гены и развитие . 12 (23): 3715–3727. дои : 10.1101/gad.12.23.3715 . ПМК 317255 . ПМИД 9851978 .
- ^ Мауро М., Берретта М., Палермо Г., Кавальери В., Ла Рокка Г. (июнь 2022 г.). «Множественность комплексов Аргонавта в клетках млекопитающих» . J Pharmacol Exp Ther . дои : 10.1124/jpet.122.001158 . ПМЦ 9827513 . ПМИД 35667689 .
- ^ Йонас С., Изаурральде Э. (июль 2015 г.). «На пути к молекулярному пониманию молчания генов, опосредованного микроРНК». Обзоры природы. Генетика . 16 (7): 421–433. дои : 10.1038/nrg3965 . ПМИД 26077373 . S2CID 24892348 .
- ^ Бомерт К., Камю И., Беллини С., Буше Д., Кабош М., Беннинг С. (январь 1998 г.). «AGO1 определяет новый локус Arabidopsis, контролирующий развитие листьев» . Журнал ЭМБО . 17 (1): 170–180. дои : 10.1093/emboj/17.1.170 . ПМК 1170368 . ПМИД 9427751 .
- ^ Го Х, Инголия НТ, Вайсман Дж.С., Бартель Д.П. (август 2010 г.). «МикроРНК млекопитающих преимущественно снижают уровни целевой мРНК» . Природа . 466 (7308): 835–840. Бибкод : 2010Natur.466..835G . дои : 10.1038/nature09267 . ПМК 2990499 . ПМИД 20703300 .
- ^ Купфершмидт К. (август 2013 г.). «Смертельная доза РНК» . Наука . 341 (6147): 732–733. Бибкод : 2013Sci...341..732K . дои : 10.1126/science.341.6147.732 . ПМИД 23950525 .
- ^ Грегори Р.И., Чендримада Т.П., Куч Н., Шихаттар Р. (ноябрь 2005 г.). «Человеческий RISC сочетает биогенез микроРНК и посттранскрипционное молчание генов» . Клетка . 123 (4): 631–640. дои : 10.1016/j.cell.2005.10.022 . ПМИД 16271387 .
- ^ Перейти обратно: а б с Хутвагнер Г., Симард М.Дж. (январь 2008 г.). «Белки-аргонавты: ключевые игроки в подавлении РНК». Обзоры природы. Молекулярно-клеточная биология . 9 (1): 22–32. дои : 10.1038/nrm2321 . hdl : 10453/15429 . ПМИД 18073770 . S2CID 8822503 .
- ^ Перейти обратно: а б Мейстер Г., Ландталер М., Патканёвска А., Дорсетт Ю., Тенг Г., Тушль Т. (июль 2004 г.). «Человеческий Argonaute2 опосредует целенаправленное расщепление РНК с помощью микроРНК и миРНК» . Молекулярная клетка . 15 (2): 185–197. doi : 10.1016/j.molcel.2004.07.007 . ПМИД 15260970 .
- ^ Майнс Ф., Си-Аммур А., Блевинс Т. (2005). «Системы подавления РНК и их значение для развития растений». Ежегодный обзор клеточной биологии и биологии развития . 21 (1): 297–318. doi : 10.1146/annurev.cellbio.21.122303.114706 . ПМИД 16212497 .
- ^ Цяо Д., Зееман А.М., Дэн В., Лоойженга Л.Х., Линь Х. (июнь 2002 г.). «Молекулярная характеристика hiwi, человеческого члена семейства генов piwi, сверхэкспрессия которого коррелирует с семиномами». Онкоген . 21 (25): 3988–3999. дои : 10.1038/sj.onc.1205505 . ПМИД 12037681 . S2CID 6078065 .
- ^ Росс Р.Дж., Вайнер М.М., Лин Х. (январь 2014 г.). «Белки PIWI и PIWI-взаимодействующие РНК в соме» . Природа . 505 (7483): 353–359. дои : 10.1038/nature12987 . ПМЦ 4265809 . ПМИД 24429634 .
- ^ Хэннон Дж.Дж. (июль 2002 г.). «РНК-интерференция» . Природа . 418 (6894): 244–251. Бибкод : 2002Natur.418..244H . дои : 10.1038/418244a . ПМИД 12110901 .
- ^ Сираноски Д. (2017). «Авторы отказываются от спорного исследования редактирования генов NgAgo» . Природа . дои : 10.1038/nature.2017.22412 .
- ^ Энгиад Б., Чжао Х. (май 2017 г.). «Программируемые ферменты искусственной рестрикции, управляемые ДНК». ACS Синтетическая биология . 6 (5): 752–757. doi : 10.1021/acsynbio.6b00324 . ПМИД 28165224 . S2CID 3833124 .
- ^ Табатабаи С.К., Ван Б., Атрея Н.Б., Энгиад Б., Эрнандес А.Г., Филдс С.Дж. и др. (апрель 2020 г.). «Перфокарты ДНК для хранения данных о нативных последовательностях ДНК посредством ферментативного надреза» . Природные коммуникации . 11 (1): 1742. Бибкод : 2020NatCo..11.1742T . дои : 10.1038/s41467-020-15588-z . ПМК 7142088 . ПМИД 32269230 .
Внешние ссылки
[ редактировать ]- База данных starBase : база данных для изучения карт взаимодействия микроРНК-мРНК на основе данных Argonaute CLIP-Seq ( HITS-CLIP , PAR-CLIP ) и Degradome-Seq.