Бета-пропеллерная фитаза
β-пропеллерные фитазы ( BPP ) представляют собой группу ферментов (т.е. суперсемейство белков ) с круглой структурой бета-пропеллера . BPP являются фитазами , а это означает, что они способны удалять ( гидролизовать ) фосфатные группы из фитиновой кислоты и ее фитатных солей. [2] Гидролиз происходит ступенчато и обычно заканчивается образованием мио -инозитолтрифосфатного продукта, с которым все еще связаны три фосфатные группы. [3] Фактическим субстратом BPP является кальция . фитат [4] и чтобы гидролизовать его, BPP должны содержать Ca 2+ ионы, связанные сами с собой. BPP являются наиболее широко встречающимся суперсемейством фитаз в окружающей среде, и считается, что они играют важную роль в круговороте фитатов и фосфора в почве и воде. [5] Как следует из альтернативного названия «щелочная фитаза» , BPP лучше всего работают в основной (или нейтральной) среде. Их оптимум pH составляет 6–9, [2] что является уникальным среди фитаз. [5]
Возможное использование
[ редактировать ]По состоянию на апрель 2018 года БПП не используются в коммерческих целях, но у них может быть потенциал для такого использования. Гистидиновые кислые фитазы (ГАК) — единственная группа фитаз, которые на данный момент используются в кормах для животных.
Корма для животных
[ редактировать ]Рекомбинантные фитазы обычно добавляют в сельском хозяйстве в корм животным с однокамерным желудком питательных веществ корма для повышения биодоступности . [6] Эти питательные вещества включают фосфор , который связан с фитатами в форме их фосфатных групп. В отличие от жвачных животных , таких как крупный рогатый скот , кишечные бактерии животных с однокамерным желудком, таких как свиньи и куры, не могут должным образом гидролизовать эти группы, чтобы пищеварительная система животного могла использовать фосфор. Таким образом, неабсорбированный фосфор тратится впустую и может попасть в окружающую среду с навозом животных через сельскохозяйственные стоки и вызвать эвтрофикацию . Фитиновая кислота также может действовать как антинутриент : она может хелатировать кальций из корма и снижать его биодоступность до 60–70% от общего содержания кальция в корме. Добавление фитазы улучшает доступность кальция, а также может улучшить биодоступность железа и цинка . Это также может увеличить доступность меди и марганца . Биодоступность аминокислот существенно не увеличивается. [7]
По сравнению с фитазами гистидиновой кислоты (HAP), которые часто нестабильны при температурах выше 65 ° C, BPP естественным образом могут выдерживать высокие температуры 80–85 ° C. Такие температуры обычно используются при гранулировании во корма для животных время его производства. В отличие от ГАП, БПП имеют нейтральный или щелочной оптимум pH, что позволяет использовать их в нейтральных или щелочных средах. Это расширяет потенциальные возможности применения фитаз. [2] BPP можно использовать в кормах для водных животных, поскольку многие из этих животных, такие как рыбы и креветки, имеют нейтральный или щелочной желудочно-кишечный тракт . [8] В отличие от HAP, BPP также специфичны для фитатов. [5] которые гидролизуют также другие фосфатсодержащие молекулы, такие как АДФ , ГТФ и НАДН . [9] Однако BPP каталитически более чем в 2–60 раз медленнее нынешних HAP. ГАП обладают удельной каталитической активностью 100–3000 ЕД мг. −1 . BPP обычно имеют удельную каталитическую активность менее 50 U мг. −1 . [2] Из-за такой низкой активности практическое использование БПП требует гораздо большего количества исследований. [5]
Структура
[ редактировать ]По состоянию на апрель 2018 года было известно 7 кристаллических структур BPP : 3AMR , 3AMS , 1H6L , 1POO , 2POO , 1CVM и 1QLG .
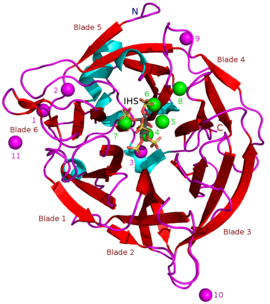
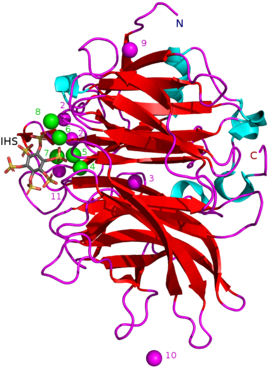
известных БПП составляют примерно 35–68 кДа . Массы [5] Их структура β-пропеллера в форме пончика состоит из 6 антипараллельных структур бета-листов или «лопастей». Один из этих клинков имеет 5 β-листов (лезвие номер 5 на рисунке в начале статьи), а остальные имеют 4 β-листа. Между этими лопастями существуют гидрофобные взаимодействия, которые, как полагают, удерживают конструкцию пропеллера вместе. Эти лезвия образуют туннельное отверстие в ферменте. Этот туннель связывает некоторые молекулы воды. Перед туннелем находится активный центр фермента , который в целом заряжен положительно благодаря Са. 2+ ионы, с которыми он связывается, и определенные положительные аминокислотные остатки. Этот сайт связывает отрицательно заряженный фитат кальция и гидролизует из него фосфаты. [2]
Узоры
[ редактировать ]
Исследование Huang et al., проведенное в 2008 году. BPP сравнили 66 пептидных последовательностей и обнаружили, что мотивы последовательности DA[A/T/E]DDPA[I/L/V]W и NN[V/I]D[I/L/V]R[Y/D/Q] были сохранены во всех изученных БПП. Например, R[Y/D/Q] означает, что R и Y , D или Q , т.е. RY или RD или RQ. в последовательности были обнаружены [10] Исследование Кумара и др., 2014 г. сравнили 44 BPP и обнаружили 10 мотивов. Два из них, DDPAIW[VI][HN]PK[DN]P[ESA]KS и NN[F/V]D[I/V/L], были обнаружены во всех изученных BPP. Было отмечено, что они аналогичны тем, которые были обнаружены в исследовании 2008 года Huang et al. [5]
Зависимость от кальция и ингибиторы
[ редактировать ]BPP представляют собой кальций-зависимые металлопротеины . Их активный центр содержит ряд катионов кальция (Ca 2+ ) связан с ним через отрицательно заряженные группы аминокислот карбоксилатные . Положительные ионы кальция необходимы для того, чтобы сделать связывание отрицательного фитата электрически выгодным. Связывание происходит через отрицательно заряженные фосфатные группы фитата вместе с определенными положительными аминокислотными остатками BPP, которые связываются непосредственно с фитатом. [1] Что 2+ концентрации также влияют на оптимум pH и термостабильность BPP: например, с Bacillus sp . ХУ-10 БПП активность наибольшая при добавлении 10 2 мМ CaCl при 60 °С и рН 6–9,5. Без добавления CaCl 2 наибольшая активность при pH наблюдается при 40 °C и pH 6,5–8,5. [5]
Удаление Ca 2+ приводит к потере каталитической активности, поэтому Ca 2+ Хелатирующая ЭДТА ингибирует BPP. Определенные точечные мутации аминокислот, связывающих кальций, также останавливают функцию фермента. Двухвалентные ионы, такие как Cd 2+ , Мн 2+ , С 2+ , Нет 2+ , ртуть 2+ , Зн 2+ , Ко 2+ и Fe 2+ ингибируют BPP путем замены Ca 2+ внутри фермента. [5] Вероятно, это связано с тем, что эти катионы слишком малы по сравнению с Ca. 2+ ионы, имеющие радиус Ван-дер-Ваальса (WDV) 0,99 Å . Ко 2+ например, имеет VDW 0,74 Å и поэтому, вероятно, слишком мал для выполнения тех же задач, что и Ca. 2+ может. Однако сэр 2+ ионы могут заменить Ca 2+ по крайней мере, в некоторых случаях, без полной потери каталитической функции. старший 2+ Радиус VDW составляет 1,12 Å и аналогичен радиусу Ca. 2+ . [4] Подобную совместимость между различными ионами можно увидеть и у некоторых других ферментов. [11]
Высокий Ca 2+ концентрация может повысить скорость катализа BPP до предела. Когда Ка 2+ концентрация превышает этот предел, дополнительный Ca 2+ ионы начинают действовать как конкурентные ингибиторы . Высокие концентрации свободного фитата, не связанного с Ca. 2+ также ингибирует BPP. Возможно, это происходит за счет хелатирования Са, опосредованного свободным фитатом. 2+ привязаны к BPP. [4]
Другие типы ингибиторов включают оксианионные (связывающие кислород) молибдат , вольфрамат и ванадат . Было высказано предположение, что ингибирование этими оксианионами происходит потому, что они образуют тригональные бипирамидальные комплексы внутри активного центра фермента, которые аналогичны переходному состоянию фосфатной группы фитата во время его гидролиза. Ортофосфат , высвобождаемый из фитиновой кислоты, действует как конкурентный ингибитор BPP. [9] Аналог фитиновой кислоты мио -инозитол-гексасульфат (IHS) ингибирует BPP, и его использовали в качестве вспомогательного средства в структурных исследованиях BPP из-за его сходства с фитиновой кислотой (см. 3AMR ). [2]
Механизм гидролиза
[ редактировать ]В механизме гидролиза, предложенном в 2001 году Shin et al. Ка 2+ Ионы, связанные с BPP, делятся на гидролизного и аффинного центра ионы . В месте гидролиза Ca 2+ помочь фосфату, который должен быть удален, связаться с активным центром . Они также активируют молекулу воды, которая участвует в гидролизе, превращая ее в ОН. − ион и стабилизируют переходное состояние в ходе катализа. Ионы области сродства увеличивают сродство фитата к активному центру и удерживают фитат в неподвижном состоянии во время гидролиза от других его фосфатных групп. Гидролиз поэтапно повторяют до мио - инозитолового продукта с тремя фосфатами. получения [3] Другие исследования подтверждают наличие в продукте трех фосфатов, [12] [13] [14] [15] но больше фосфатов можно удалить в экстремальных условиях, таких как высокая концентрация BPP и удлиненное время инкубации. [13]

Фактический гидролиз, предложенный Shin et al. происходит в два этапа. Второй медленнее и ограничивает общую скорость реакции. На первом этапе карбонильная группа отбирает электроны у фосфора фосфата, делая его бедным электронами, то есть оставляя его с положительным зарядом. Одновременно ОН − отдает электронную пару номинально положительному фосфору в фосфате. состояние . Формируется тригонально-бипирамидальное промежуточное На втором этапе фосфоэфирная связь разрывается, когда кислый аминокислотный остаток (BH + ) отдает протон кислороду в связи. Фосфатная группа таким образом отщепляется. [3]
Маршруты гидролиза
[ редактировать ]Существует несколько предполагаемых путей гидролиза, но неизвестно, какой из них правильный или существует ли несколько путей. Эти пути гидролиза кратко описаны ниже.

См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Цзэн Ю.Ф., Ко Т.П., Лай Х.Л., Ченг Ю.С., Ву Т.Х., Ма Ю., Чен К.С., Ян К.С., Ченг К.Дж., Хуан Ч.Х., Го RT, Лю-младший (июнь 2011 г.). «Кристаллические структуры щелочной фитазы Bacillus в комплексе с ионами двухвалентных металлов и гексасульфатом инозитола» (PDF) . Журнал молекулярной биологии . 409 (2): 214–24. дои : 10.1016/j.jmb.2011.03.063 . ПМИД 21463636 .
- ^ Перейти обратно: а б с д и ж Чен CC, Ченг К.Дж., Ко Т.П., Го RT (01.04.2015). «Текущий прогресс в исследованиях фитазы: трехмерная структура и белковая инженерия» . ХимБиоИнж Обзоры . 2 (2): 76–86. дои : 10.1002/cben.201400026 .
- ^ Перейти обратно: а б с д Шин С., Ха NC, Oh BC, Oh TK, Oh BH (сентябрь 2001 г.). «Ферментный механизм и каталитические свойства бета-пропеллерной фитазы» . Структура . 9 (9): 851–8. дои : 10.1016/S0969-2126(01)00637-2 . ПМИД 11566134 .
- ^ Перейти обратно: а б с Ха NC, Oh BC, Shin S, Kim HJ, Oh TK, Kim YO, Choi KY, Oh BH (февраль 2000 г.). «Кристаллические структуры новой термостабильной фитазы в частично и полностью нагруженных кальцием состояниях». Структурная биология природы . 7 (2): 147–53. дои : 10.1038/72421 . ПМИД 10655618 . S2CID 29391671 .
- ^ Перейти обратно: а б с д и ж г час Кумар В., Ядав А.Н., Верма П., Сангван П., Саксена А., Кумар К., Сингх Б. (май 2017 г.). «β-Пропеллерные фитазы: разнообразие, каталитические свойства, текущие разработки и потенциальные биотехнологические применения». Международный журнал биологических макромолекул . 98 : 595–609. doi : 10.1016/j.ijbiomac.2017.01.134 . ПМИД 28174082 .
- ^ Гифре Л., Арис А., Бах А., Гарсия-Фруитос Э. (март 2017 г.). «Тенденции использования рекомбинантных белков в животноводстве» . Заводы по производству микробных клеток . 16 (1): 40. дои : 10.1186/s12934-017-0654-4 . ПМК 5336677 . ПМИД 28259156 .
- ^ Лей XG, Уивер Дж.Д., Маллани Э., Улла А.Х., Азаин М.Дж. (январь 2013 г.). «Фитаза, новая жизнь «старого» фермента». Ежегодный обзор биологических наук о животных . 1 (1): 283–309. doi : 10.1146/annurev-animal-031412-103717 . ПМИД 25387021 .
- ^ Виадер-Сальвадо Х.М., Гальегос-Лопес Х.А., Карреон-Тревиньо Х.Г., Кастильо-Гальван М., Рохо-Домингес А., Герреро-Олазаран М. (октябрь 2010 г.). «Создание термостабильных бета-пропеллерных фитаз с активностью в широком диапазоне pH и их сверхпродукция Pichia Pastoris» . Прикладная и экологическая микробиология . 76 (19): 6423–30. Бибкод : 2010ApEnM..76.6423V . дои : 10.1128/AEM.00253-10 . ПМЦ 2950461 . ПМИД 20693453 .
- ^ Перейти обратно: а б с Коницкий Ю, Грейнер Р. (2002). «Молекулярные и каталитические свойства ферментов, расщепляющих фитаты (фитаз)» . Международный журнал пищевой науки и технологий . 37 (7): 791–812. дои : 10.1046/j.1365-2621.2002.00617.x .
- ^ Перейти обратно: а б Хуан Х, Шао Н, Ван Ю, Луо Х, Ян П, Чжоу З, Чжан Цз, Яо Б (май 2009 г.). «Новая бета-пропеллерная фитаза из Pedobacter nyackensis MJ11 CGMCC 2503 с потенциалом использования в качестве кормовой добавки для водных животных». Прикладная микробиология и биотехнология . 83 (2): 249–59. дои : 10.1007/s00253-008-1835-1 . ПМИД 19139877 . S2CID 20225761 .
- ^ Дёберейнер А., Шмид А., Людвиг А., Гебель В., Бенц Р. (сентябрь 1996 г.). «Влияние кальция и других поливалентных катионов на образование каналов альфа-гемолизином Escherichia coli в эритроцитах и липидных бислойных мембранах» . Европейский журнал биохимии . 240 (2): 454–60. дои : 10.1111/j.1432-1033.1996.0454h.x . ПМИД 8841412 .
- ^ Перейти обратно: а б Керовуо Дж., Рувинен Дж., Хацак Ф. (декабрь 2000 г.). «Анализ гидролиза мио-инозитол-гексакисфосфата фитазой Bacillus: указание на новый механизм реакции» . Биохимический журнал . 352 Ч. 3 (Часть 3): 623–8. дои : 10.1042/bj3520623 . ПМЦ 1221497 . ПМИД 11104666 .
- ^ Перейти обратно: а б с д Грейнер Р., Ларссон Альмингер М., Карлссон Н.Г., Музкиз М., Бурбано С., Куадрадо С., Педроса М.М., Гойоага С. (2002). «Путь дефосфорилирования мио-инозитолгексакисфосфата фитазами семян бобовых». Журнал сельскохозяйственной и пищевой химии . 50 (23): 6865–6870. дои : 10.1021/jf025620t . ПМИД 12405789 .
- ^ Перейти обратно: а б О BC, Ким М.Х., Юн Б.С., Чой В.К., Парк СК, Бэ СК, О ТК (август 2006 г.). «Хелирование Са (2+)-инозитолфосфата опосредует субстратную специфичность бета-пропеллер-фитазы». Биохимия . 45 (31): 9531–9. дои : 10.1021/bi0603118 . ПМИД 16878987 .
- ^ Перейти обратно: а б Нам С.Дж., Ким Ё, Ко ТК, Кан Дж.К., Чун К.Х., Ау Дж.Х., Ли К.С., Ли И.К., Пак С., О, Британская Колумбия (октябрь 2014 г.). «Молекулярные и биохимические характеристики β-пропеллерной фитазы морских Pseudomonas sp. BS10-3 и ее потенциальное применение в кормовых добавках для животных» . Журнал микробиологии и биотехнологии . 24 (10): 1413–20. дои : 10.4014/jmb.1407.07063 . ПМИД 25112322 .