5S рибосомальная РНК
| 5S рибосомальная РНК | |
|---|---|
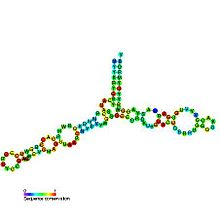 Предсказанная вторичная структура и консервативность последовательности 5S рибосомальной РНК | |
| Идентификаторы | |
| Символ | 5S_рРНК |
| Рфам | RF00001 CL00113 |
| Другие данные | |
| РНК Тип | ген ; рРНК |
| Домен(ы) | Эукариоты ; Бактерии ; Архея |
| ИДТИ | ПОЙДИТЕ: 0005840 ПОЙДИТЕ: 0003735 |
| ТАК | ТАК: 0000652 |
| PDB Структуры | ПДБе |
5S рибосомальная РНК ( 5S рРНК длиной примерно 120 нуклеотидов ) представляет собой молекулу рибосомной РНК и массой 40 кДа . Это структурный и функциональный компонент большой субъединицы рибосомы во всех сферах жизни ( бактерии , археи и эукариоты ), за исключением митохондриальных рибосом грибов и животных. молекулы Обозначение 5S относится к коэффициенту седиментации в ультрацентрифуге, который измеряется в единицах Сведберга (S). [ 1 ]

Биосинтез
[ редактировать ]У прокариот ген 5S рРНК обычно расположен в оперонах рРНК ниже малой и большой субъединицы рРНК и совместно транскрибируется в полицистронный предшественник . [ 3 ] Особенностью ядерных геномов эукариот является наличие множественных копий гена 5S рРНК (5S рДНК), сгруппированных в тандемные повторы, причем количество копий варьируется от вида к виду. [ 4 ] [ 5 ] Эукариотическая 5S рРНК синтезируется РНК-полимеразой III , тогда как другие эукариотические рРНК отщепляются от предшественника 45S, транскрибируемого I. РНК- полимеразой В Xenopus ооцитах было показано, что пальцы 4–7 девятицинковых пальцев транскрипционного фактора TFIIIA могут связываться с центральной областью 5S РНК. [ 6 ] [ 7 ] Связывание между 5S рРНК и TFIIIA служит как для подавления дальнейшей транскрипции 5S РНК гена , так и для стабилизации транскрипта 5S РНК до тех пор, пока он не понадобится для сборки рибосомы. [ 8 ]
Структура
[ редактировать ]Вторичная структура 5S рРНК состоит из пяти спиралей (обозначенных римскими цифрами I–V ), четырех петель (BE) и одного шарнира (А), которые вместе образуют Y-подобную структуру. Петли C и D являются концевыми шпильками, а петли B и E — внутренними. [ 4 ] Согласно филогенетическим исследованиям, спирали I и III, вероятно, являются предковыми. [ 9 ] Helix III включает два высококонсервативных аденозина. [ 10 ] Считается, что спираль V с ее шпилечной структурой взаимодействует с TFIIIA. [ 4 ]
Расположение внутри рибосомы
[ редактировать ]
С помощью различных молекулярных методов, включая иммуноэлектронную микроскопию , криоэлектронную микроскопию , межмолекулярную химическую сшивку и рентгеновскую кристаллографию , расположение 5S рРНК внутри большой субъединицы рибосомы было определено с большой точностью. У бактерий и архей большая рибосомальная субъединица (LSU) сама по себе состоит из двух фрагментов РНК, 5S рРНК и другой более крупной РНК, известной как 23S рРНК , а также многочисленных связанных с ними белков. [ 3 ] [ 12 ]
У эукариот LSU содержит 5S, 5.8S и 28S рРНК и еще больше белков. [ 13 ] [ 14 ] Структура LSU в трех измерениях показывает одну относительно гладкую поверхность и противоположную поверхность, имеющую три выступа, а именно выступ L1, центральный выступ (CP) и ножку L7/L12. Выступ L1 и ножка L7/L12 расположены латерально вокруг CP. 5S рРНК расположена в ЦП и участвует в формировании и строении этой проекции. Другие основные составляющие центрального выступа включают 23S рРНК (или, альтернативно, 28S у эукариот) и несколько белков, включая L5, L18, L25 и L27. [ 15 ]
Рибосомальные функции
[ редактировать ]Точная функция 5S рРНК пока не ясна. У Escherichia coli делеции гена 5S рРНК снижают скорость синтеза белка и оказывают более глубокое вредное воздействие на приспособленность клеток, чем делеции сопоставимого количества копий других генов ( 16S и 23S ) рРНК. [ 16 ] Кристаллографические исследования показывают, что 5S рРНК-связывающие белки и другие белки центрального выступа LSU играют роль в связывании тРНК. [ 15 ] Также топографическая и физическая близость 5S рРНК и 23S рРНК, образующей пептидилтрансферазу и центр, ассоциированный с ГТФазой, позволяет предположить, что 5S рРНК выступает посредником между двумя функциональными центрами рибосомы, образуя вместе с 5S рРНК-связывающим центром рибосому белки и другие компоненты центрального выступа, межсубъединичных мостиков и сайтов связывания тРНК. [ 15 ]
Роли в сборке рибосом
[ редактировать ]У эукариот цитозольная рибосома состоит из четырех рРНК и более 80 белков. [ 14 ] [ 17 ] После транскрипции 3'-концы 5S рРНК могут быть обрезаны до зрелой длины только функциональными гомологами РНКазы Т , например Rex1p в Saccharomyces cerevisiae . [ 18 ] Субъединицы рибосом 60S и 40S экспортируются из ядра в цитоплазму, где они соединяются, образуя зрелую и компетентную к трансляции 80S рибосому. Когда именно 5S рРНК интегрируется в рибосому, остается спорным. [ 4 ] но общепринято, что 5S рРНК включена в частицу 90S, которая является предшественником частицы 60S, как часть небольшого независимого от рибосомы комплекса РНП, образованного 5S рРНК и рибосомальным белком L5. [ 17 ]
Взаимодействие с белками
[ редактировать ]Ниже перечислены несколько важных белков, которые взаимодействуют с 5S рРНК.
Белок
[ редактировать ]Взаимодействие 5S рРНК с белком La предотвращает деградацию РНК экзонуклеазами в клетке. [ 19 ] Белок La обнаружен в ядре всех эукариотических организмов и связан с несколькими типами РНК, транскрибируемыми РНК pol III. Белок La взаимодействует с этими РНК (включая 5S рРНК) через их 3'-олигоуридиновый тракт, способствуя стабильности и сворачиванию РНК. [ 4 ] [ 20 ]
белок L5
[ редактировать ]В эукариотических клетках рибосомальный белок L5 связывает и стабилизирует 5S рРНК, образуя прерибосомальную рибонуклеопротеиновую частицу (РНП), которая обнаруживается как в цитозоле, так и в ядре. Дефицит L5 предотвращает транспорт 5S рРНК в ядро и приводит к снижению сборки рибосом. [ 4 ]
Другие рибосомальные белки
[ редактировать ]У прокариот 5S рРНК связывается с рибосомными белками L5, L18 и L25, тогда как у эукариотов 5S рРНК, как известно, связывается только с рибосомальным белком L5 . [ 21 ] У T. brucei , возбудителя сонной болезни , 5S рРНК взаимодействует с двумя близкородственными РНК-связывающими белками, P34 и P37, потеря которых приводит к снижению глобального уровня 5S рРНК. [ 4 ]
Присутствие в рибосомах органелл
[ редактировать ]| Пермутированный митохондриальный геном, кодирующий 5S рРНК | |
|---|---|
| Идентификаторы | |
| Символ | мтПермь-5С |
| Рфам | RF02547 CL00113 |
| Другие данные | |
| РНК Тип | ген ; рРНК |
| Домен(ы) | Эукариоты ; |
| ИДТИ | ПОЙДИТЕ: 0005840 ПОЙДИТЕ: 0003735 |
| ТАК | ТАК: 0000652 |
| PDB Структуры | ПДБе |

Механизмы трансляции митохондрий и пластид (органелл эндосимбиотического бактериального происхождения) и их бактериальных родственников имеют много общих черт, но также демонстрируют заметные различия. Геномы органелл без исключения кодируют рРНК SSU и LSU, однако распределение генов 5S рРНК ( rrn5 ) крайне неравномерно. Rrn5 легко идентифицируется и часто встречается в геномах большинства пластид. Напротив, митохондриальный rrn5 первоначально, по-видимому, был ограничен растениями и небольшим количеством простейших. [ 22 ] [ 23 ] Дополнительные, более дивергентные органелларные 5S рРНК были идентифицированы только с помощью специализированных ковариационных моделей, которые включают информацию о выраженной систематической ошибке состава последовательностей и структурных вариациях. [ 24 ] Этот анализ выявил дополнительные гены 5S рРНК не только в митохондриальных геномах большинства линий протистов , но также и в геномах некоторых апикопластов (нефотосинтезирующих пластид патогенных простейших, таких как Toxoplasma gondii и Eimeria tenella ).

Митохондриальные 5S рРНК большинства страменопилей содержат наибольшее разнообразие вторичных структур. [ 24 ] Перестановленные митохондриальные 5S рРНК в бурых водорослях представляют собой наиболее нетрадиционный случай, когда замыкающаяся спираль I, которая в противном случае объединяет 5'- и 3'-концы молекулы, заменяется (закрытой) шпилькой, что приводит к открытому трехстороннему соединению.
Современные данные указывают на то, что в митохондриальной ДНК лишь нескольких групп, особенно животных , грибов , альвеолятов и эвгленозойных генов отсутствует этот ген. [ 24 ] Центральный выступ , в противном случае занятый 5S рРНК и связанными с ней белками (см. рисунок 2 ), был ремоделирован различными способами. В митохондриальных рибосомах грибов 5S рРНК заменена последовательностями расширения LSU рРНК. [ 25 ] У кинетопластид (эвгленозоев) центральный выступ полностью состоит из эволюционно новых митохондриальных рибосомальных белков. [ 26 ] Наконец, митохондриальные рибосомы животных внедрили специфическую митохондриальную тРНК (Val у позвоночных), чтобы заменить недостающую 5S рРНК. [ 27 ] [ 28 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Шиманский М., Барчишевска М.З., Эрдманн В.А., Барцишевский Дж. (январь 2002 г.). «База данных 5S рибосомальных РНК» . Исследования нуклеиновых кислот . 30 (1): 176–178. дои : 10.1093/нар/30.1.176 . ПМК 99124 . ПМИД 11752286 .
- ^ Мюллер Ф., Зоммер И., Баранов П., Матадин Р., Столдт М., Вёнерт Дж. и др. (апрель 2000 г.). «Трехмерное расположение 23 S и 5 S рРНК в 50 S рибосомальной субъединице Escherichia coli на основе криоэлектронной микроскопической реконструкции с разрешением 7,5 А». Журнал молекулярной биологии . 298 (1): 35–59. дои : 10.1006/jmbi.2000.3635 . ПМИД 10756104 .
- ^ Jump up to: а б Качановска М., Риден-Аулин М. (сентябрь 2007 г.). «Биогенез рибосом и процесс трансляции в Escherichia coli» . Обзоры микробиологии и молекулярной биологии . 71 (3): 477–494. дои : 10.1128/MMBR.00013-07 . ПМК 2168646 . ПМИД 17804668 .
- ^ Jump up to: а б с д и ж г Сиганда М., Уильямс Н. (2011). «Биогенез эукариотической 5S рРНК» . Междисциплинарные обзоры Wiley. РНК . 2 (4): 523–533. дои : 10.1002/wrna.74 . ПМЦ 3278907 . ПМИД 21957041 .
- ^ Дуэ Ж, Турменте С (июль 2007 г.). «Транскрипция гетерохроматических генов 5S рРНК эпигенетически контролируется у Arabidopsis thaliana и Xenopus laevis» . Наследственность . 99 (1): 5–13. дои : 10.1038/sj.hdy.6800964 . ПМИД 17487217 .
- ^ МакБрайант С.Дж., Вельдхоэн Н., Гедулин Б., Лереш А., Фостер М.П., Райт П.Е. и др. (апрель 1995 г.). «Взаимодействие РНК-связывающих пальцев транскрипционного фактора IIIA Xenopus со специфическими участками 5S рибосомальной РНК». Журнал молекулярной биологии . 248 (1): 44–57. дои : 10.1006/jmbi.1995.0201 . ПМИД 7731045 .
- ^ Сирлз М.А., Лу Д., Клуг А. (август 2000 г.). «Роль центральных цинковых пальцев транскрипционного фактора IIIA в связывании с 5S РНК». Журнал молекулярной биологии . 301 (1): 47–60. дои : 10.1006/jmbi.2000.3946 . ПМИД 10926492 .
- ^ Пелхэм Х.Р., Браун Д.Д. (июль 1980 г.). «Специфический фактор транскрипции, который может связывать либо ген 5S РНК, либо ген 5S РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (7): 4170–4174. Бибкод : 1980PNAS...77.4170P . дои : 10.1073/pnas.77.7.4170 . ПМК 349792 . ПМИД 7001457 .
- ^ Сан Ф.Дж., Каэтано-Аноллес Дж. (ноябрь 2009 г.). «Эволюционная история структуры 5S рибосомальной РНК». Журнал молекулярной эволюции . 69 (5): 430–443. Бибкод : 2009JMolE..69..430S . дои : 10.1007/s00239-009-9264-z . ПМИД 19639237 . S2CID 21572917 .
- ^ ДиНитто Дж.П., Хубер П.В. (октябрь 2001 г.). «Роль ароматических аминокислот в связывании рибосомного белка L5 Xenopus с 5S рРНК». Биохимия . 40 (42): 12645–12653. дои : 10.1021/bi011439m . ПМИД 11601989 .
- ^ Бан Н., Ниссен П., Хансен Дж., Мур П.Б., Стейц Т.А. (август 2000 г.). «Полная атомная структура большой субъединицы рибосомы с разрешением 2,4 А». Наука . 289 (5481): 905–920. Бибкод : 2000Sci...289..905B . CiteSeerX 10.1.1.58.2271 . дои : 10.1126/science.289.5481.905 . ПМИД 10937989 .
- ^ Тирумалай М.Р., Кельбер Дж.Т., Парк Д.Р., Тран К., Fox GE (октябрь 2020 г.). «Визуализация криоэлектронной микроскопией большой вставки в 5S рибосомальной РНК чрезвычайно галофильной археи Halococcus morrhuae» . Открытая биография FEBS . 10 (10): 1938–1946. дои : 10.1002/2211-5463.12962 . ПМЦ 7530397 . PMID 32865340 .
- ^ Туровский Т.В., Толлервей Д. (2015). «Котранскрипционные события в синтезе эукариотических рибосом» . Междисциплинарные обзоры Wiley. РНК . 6 (1): 129–139. дои : 10.1002/wrna.1263 . hdl : 20.500.11820/983ebe02-f5a9-4b65-a3f6-c93ac8088fb2 . ПМИД 25176256 .
- ^ Jump up to: а б Юсупова Г, Юсупов М (февраль 2014 г.). «Структура высокого разрешения эукариотической рибосомы 80S». Ежегодный обзор биохимии . 83 : 467–486. doi : 10.1146/annurev-biochem-060713-035445 . ПМИД 24580643 .
- ^ Jump up to: а б с Гонгадзе Г.М. (декабрь 2011 г.). «5S рРНК и рибосома». Биохимия. Биохимия . 76 (13): 1450–1464. дои : 10.1134/S0006297911130062 . ПМИД 22339598 . S2CID 41669737 .
- ^ Аммонс Д., Рамперсад Дж., Фокс Дж. Е. (январь 1999 г.). «Делеции гена 5S рРНК вызывают неожиданно высокую потерю приспособленности у Escherichia coli» . Исследования нуклеиновых кислот . 27 (2): 637–642. дои : 10.1093/нар/27.2.637 . ПМК 148226 . ПМИД 9862991 .
- ^ Jump up to: а б Анрас А.К., Суде Ж., Жерус М., Лебарон С., Кайзерг-Феррер М., Мужен А., Анри Ю. (август 2008 г.). «Посттранскрипционные этапы биогенеза эукариотических рибосом» . Клеточные и молекулярные науки о жизни . 65 (15): 2334–2359. дои : 10.1007/s00018-008-8027-0 . ПМЦ 11131730 . ПМИД 18408888 . S2CID 1934099 .
- ^ ван Хуф А., Леннерц П., Паркер Р. (март 2000 г.). «Три консервативных члена семейства РНКазы D обладают уникальными и перекрывающимися функциями при процессинге РНК 5S, 5.8S, U4, U5, РНКазы MRP и РНКазы P в дрожжах» . Журнал ЭМБО . 19 (6): 1357–1365. дои : 10.1093/emboj/19.6.1357 . ПМК 305676 . ПМИД 10716935 .
- ^ Волин С.Л., Седервалл Т (2002). «Протеин Ла». Ежегодный обзор биохимии . 71 : 375–403. doi : 10.1146/annurev.biochem.71.090501.150003 . ПМИД 12045101 .
- ^ Марайя Р.Дж., Intine RV (2002). «Белок La и связанные с ним малые ядерные и ядрышковые РНК-предшественники» . Экспрессия генов . 10 (1–2): 41–57. ПМЦ 5977531 . ПМИД 11868987 .
- ^ Мур П.Б. (март 2001 г.). «Рибосома с атомным разрешением». Биохимия . 40 (11): 3243–3250. дои : 10.1021/bi0029402 . ПМИД 11258942 .
- ^ Bullerwell CE, Schnare MN, Grey MW (март 2003 г.). «Открытие и характеристика митохондриальной 5S рРНК Acanthamoeba castellanii» . РНК . 9 (3): 287–292. дои : 10.1261/rna.2170803 . ПМЦ 1370395 . ПМИД 12592002 .
- ^ Bullerwell CE, Burger G, Gott JM, Kourennaia O, Schnare MN, Grey MW (май 2010 г.). «Обильные 5S-рРНК-подобные транскрипты, кодируемые митохондриальным геномом амебозой» . Эукариотическая клетка . 9 (5): 762–773. дои : 10.1128/EC.00013-10 . ПМК 2863963 . ПМИД 20304999 .
- ^ Jump up to: а б с Валах М., Бургер Дж., Грей М.В., Ланг Б.Ф. (декабрь 2014 г.). «Широко распространенное распространение 5S рРНК, кодируемых геномом органелл, включая перестановочные молекулы» . Исследования нуклеиновых кислот . 42 (22): 13764–13777. дои : 10.1093/nar/gku1266 . ПМЦ 4267664 . ПМИД 25429974 .
- ^ Амунтс А., Браун А., Бай XC, Лласер Дж.Л., Хуссейн Т., Эмсли П. и др. (март 2014 г.). «Строение большой рибосомальной субъединицы митохондрий дрожжей» . Наука . 343 (6178): 1485–1489. Бибкод : 2014Sci...343.1485A . дои : 10.1126/science.1249410 . ПМК 4046073 . ПМИД 24675956 .
- ^ Шарма М.Р., Бут Т.М., Симпсон Л., Маслов Д.А., Агравал Р.К. (июнь 2009 г.). «Структура митохондриальной рибосомы с минимальной РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (24): 9637–9642. Бибкод : 2009PNAS..106.9637S . дои : 10.1073/pnas.0901631106 . ПМК 2700991 . ПМИД 19497863 .
- ^ Браун А., Амунц А., Бай XC, Сугимото Ю., Эдвардс ПК., Муршудов Г. и др. (ноябрь 2014 г.). «Строение большой рибосомальной субъединицы митохондрий человека» . Наука . 346 (6210): 718–722. Бибкод : 2014Sci...346..718B . дои : 10.1126/science.1258026 . ПМК 4246062 . ПМИД 25278503 .
- ^ Гребер Б.Дж., Берингер Д., Лейбундгут М., Биери П., Лейтнер А., Шмитц Н. и др. (ноябрь 2014 г.). «Полная структура большой субъединицы митохондриальной рибосомы млекопитающих». Природа . 515 (7526): 283–286. Бибкод : 2014Natur.515..283G . дои : 10.1038/nature13895 . hdl : 20.500.11850/93239 . ПМИД 25271403 . S2CID 4402484 .
Внешние ссылки
[ редактировать ]- Страница 5S рибосомальной РНК в Rfam
- 5SData , заархивировано 27 апреля 2010 г. на Wayback Machine.
- 5S + рибосомальная + РНК в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Расположение человеческого гена 5S_rRNA в браузере генома UCSC .
- Halococcus morrhuae (архебактерия) 5S рРНК