16S рибосомальная РНК
В этой статье отсутствует информация о Rfam SSU_rRNA_bacteria, SSU_rRNA_archaea. ( декабрь 2020 г. ) |
16 S рибосомальная РНК (или 16 S рРНК является компонентом РНК субъединицы 30S прокариотической ) рибосомы ( SSU рРНК ). Он связывается с последовательностью Шайна-Дальгарно и обеспечивает большую часть структуры SSU.
Гены, кодирующие его, называются генами 16S рРНК и используются при реконструкции филогений из-за медленных темпов эволюции этой области гена. [2] Карл Вёзе и Джордж Э. Фокс были пионерами использования 16S рРНК в филогенетике в 1977 году. [3] могут существовать несколько последовательностей гена 16S рРНК В одной бактерии . [4]
Функции [ править ]
- Как и большая (23S) рибосомальная РНК , она играет структурную роль, действуя как каркас, определяющий положения рибосомальных белков .
- конец 3' - Шайна содержит последовательность анти- -Дальгарно , которая связывается выше стартового кодона AUG на мРНК . 3' - конец 16S РНК связывается с белками S1 и S21, которые, как известно, участвуют в инициации синтеза белка. [5]
- Взаимодействует с 23S, способствуя связыванию двух субъединиц рибосомы ( 50S и 30S ).
- Стабилизирует правильное спаривание кодон-антикодон в А-сайте за счет образования водородной связи между атомом N1 остатков аденина 1492 и 1493 и 2' - ОН-группой основной цепи мРНК.
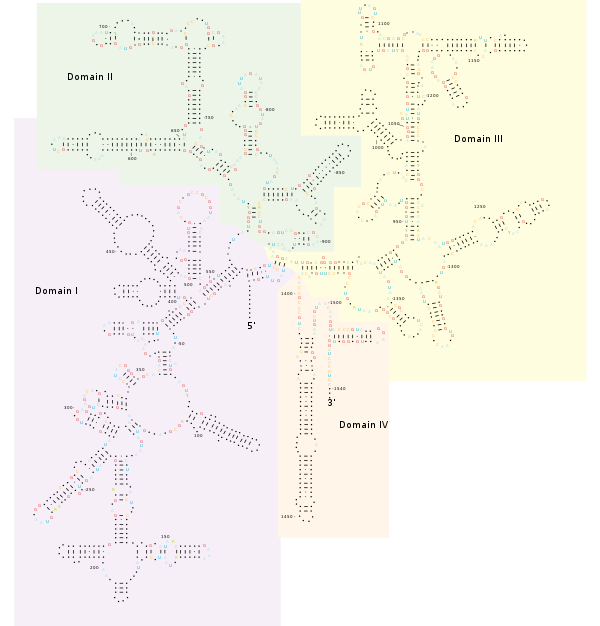
Структура [ править ]
Универсальные праймеры [ править ]
Ген 16S рРНК используется для филогенетических исследований. [7] поскольку он высоко консервативен между различными видами бактерий и архей. [8] Карл Везе впервые использовал 16S рРНК в 1977 году. [2] Предполагается, что ген 16S рРНК можно использовать в качестве надежных молекулярных часов , поскольку показано, что последовательности 16S рРНК из отдаленно родственных бактериальных линий имеют схожие функциональные возможности. [9] Некоторые термофильные археи (например, порядок Thermoproteales ) содержат интроны гена 16S рРНК , которые расположены в высококонсервативных областях и могут влиять на отжиг «универсальных» праймеров . [10] Митохондриальные и хлоропластические рРНК также амплифицируются. [11]
Наиболее распространенная пара праймеров была разработана Weisburg et al. (1991) [7] и в настоящее время обозначается как 27F и 1492R; однако для некоторых применений могут потребоваться более короткие ампликоны , например, для секвенирования 454 с использованием титановой химии пара праймеров 27F-534R, охватывающая V1-V3. [12] Часто используется 8F, а не 27F. Эти два праймера почти идентичны, но 27F имеет букву M вместо C. AGAGTTTTGATC M TGGCTCAG по сравнению с 8F. [13]
| Имя праймера | Последовательность (5 ′ –3 ′ ) | Ссылка. |
|---|---|---|
| 8F | AGA GTT TGA TCC TGG CTC AG | [14] [15] |
| 27F | AGA GTT TGA TC M TGG CTC AG | [13] |
| 336Р | ACT GCT GCS YCC CGT AGG AGT CT | [16] |
| 337F | GAC TCC TAC GGG AGG CWG CAG | [17] |
| 518Р | GTA TTA CCG CGG CTG CTG G | |
| 533F | GTG CCA GCM GCC GCG GTA A | |
| 785Ф | GGA TTA GAT ACC CTG GTA | |
| 806Р | GGA CTA CVS GGG TAT CTA AT | [18] [19] |
| 907Р | CCG TCA ATT CCT TTR AGT TT | |
| 928Ф | ТАА ААК ТИА ААК ГАА ТТГ АКГ ГГ | [16] |
| 1100F | ДА CGA GCG CAA CCC | |
| 1100р | ГГГ ТТГ КГК TCG ТТГ | |
| U1492R | GGT TAC CTT GTT ACG ACT T | [14] [15] |
| 1492р | CGG TTA CCT TGT TAC GAC TT | [20] |
Приложения ПЦР и NGS [ править ]
В дополнение к высококонсервативным сайтам связывания праймеров, последовательности генов 16S рРНК содержат гипервариабельные области , которые могут обеспечивать видоспецифичные сигнатурные последовательности, полезные для идентификации бактерий. [21] [22] В результате секвенирование гена 16S рРНК стало превалировать в медицинской микробиологии как быстрая и дешевая альтернатива фенотипическим методам идентификации бактерий. [23] Хотя первоначально секвенирование 16S использовалось для идентификации бактерий, впоследствии было обнаружено, что оно способно реклассифицировать бактерии в совершенно новые виды . [24] или даже роды . [7] [25] Его также использовали для описания новых видов, которые никогда не были успешно культивированы. [26] [27] Благодаря секвенированию третьего поколения, которое появится во многих лабораториях, одновременная идентификация тысяч последовательностей 16S рРНК станет возможной в течение нескольких часов, что позволит проводить метагеномные исследования, например, кишечной флоры . [28] В образцах, собранных у пациентов с подтвержденными инфекциями, секвенирование 16S рРНК нового поколения (NGS) продемонстрировало улучшенное обнаружение в 40% случаев по сравнению с традиционными методами культивирования; более того, прием антибиотиков перед отбором проб не оказал существенного влияния на чувствительность 16S NGS. [29]
Гипервариабельные регионы [ править ]
Бактериальный ген 16S содержит девять гипервариабельных областей (V1–V9) длиной от 30 до 100 пар оснований , которые участвуют во вторичной структуре малой рибосомальной субъединицы . [30] Степень сохранности широко варьируется между гипервариабельными регионами: более консервативные регионы соответствуют таксономии более высокого уровня, а менее консервативные регионы - более низким уровням, таким как род и вид. [31] Хотя вся последовательность 16S позволяет сравнивать все гипервариабельные области, ее длина примерно в 1500 пар оснований может быть непомерно дорогой для исследований, направленных на идентификацию или характеристику различных бактериальных сообществ. [31] В этих исследованиях обычно используется платформа Illumina , которая производит считывания со скоростью в 50 и 12 000 раз дешевле, чем пиросеквенирование 454 и секвенирование Сэнгера соответственно. [32] Хотя секвенирование Illumina дешевле и позволяет более глубоко охватить сообщество, оно дает только считывания длиной 75–250 пар оснований (до 300 пар оснований с Illumina MiSeq) и не имеет установленного протокола для надежной сборки полного гена в образцах сообщества. [33] Однако полные гипервариабельные области можно собрать за один запуск Illumina, что делает их идеальными объектами для платформы. [33]
Хотя гипервариабельные области 16S могут существенно различаться у разных бактерий, ген 16S в целом сохраняет большую однородность длины, чем его эукариотический аналог ( рибосомальная РНК 18S ), что может облегчить выравнивание . [34] Кроме того, ген 16S содержит высококонсервативные последовательности между гипервариабельными областями, что позволяет создавать универсальные праймеры, которые могут надежно производить одни и те же участки последовательности 16S в разных таксонах . [35] Хотя ни одна гипервариабельная область не может точно классифицировать все бактерии от домена к виду, некоторые из них могут надежно предсказать конкретные таксономические уровни. [31] По этой причине во многих общественных исследованиях выбираются полуконсервативные гипервариабельные области, такие как V4, поскольку они могут обеспечить разрешение на уровне типа так же точно, как и полный ген 16S. [31] В то время как менее консервативные регионы изо всех сил пытаются классифицировать новые виды, когда таксономия более высокого порядка неизвестна, они часто используются для обнаружения присутствия конкретных патогенов. В одном исследовании Chakravorty et al. в 2007 году авторы охарактеризовали регионы V1–V8 различных патогенов, чтобы определить, какие гипервариабельные регионы было бы наиболее полезно включить в специфичные для заболевания и широкие анализы . [36] Среди других результатов они отметили, что область V3 лучше всего подходит для идентификации рода всех протестированных патогенов, а V6 является наиболее точным для дифференциации видов между всеми проверенными патогенами, наблюдаемыми CDC , включая сибирскую язву . [36]
Хотя анализ гипервариабельной области 16S является мощным инструментом для таксономических исследований бактерий, он с трудом позволяет дифференцировать близкородственные виды. [35] В семействах Enterobacteriaceae , Clostridiaceae и Peptostreptococcaceae виды могут иметь сходство последовательностей до 99% по полному гену 16S. [37] В результате последовательности V4 могут отличаться всего на несколько нуклеотидов , в результате чего справочные базы данных не могут надежно классифицировать эти бактерии на более низких таксономических уровнях. [37] Ограничивая анализ 16S выборкой гипервариабельных регионов, эти исследования могут не обнаружить различий в близкородственных таксонах и сгруппировать их в единые таксономические единицы, тем самым недооценивая общее разнообразие выборки. [35] Кроме того, бактериальные геномы могут содержать несколько генов 16S, при этом области V1, V2 и V6 содержат наибольшее внутривидовое разнообразие. [8] Хотя это и не самый точный метод классификации видов бактерий, анализ гипервариабельных областей остается одним из наиболее полезных инструментов, доступных для изучения бактериального сообщества. [37]
генов Неразборчивость 16S рРНК
При предположении, что эволюция управляется вертикальной передачей , долгое время считалось, что гены 16S рРНК являются видоспецифичными и безошибочными в качестве генетических маркеров, определяющих филогенетические отношения между прокариотами . Однако все большее число наблюдений позволяет предположить наличие горизонтального переноса этих генов. Помимо наблюдений за естественным возникновением, переносимость этих генов подтверждается экспериментально с использованием специализированной генетической системы Escherichia coli . Было показано, что при использовании нулевого мутанта в E. coli качестве хозяина рост мутантного штамма дополняется чужеродными генами 16S рРНК, которые филогенетически отличались от E. coli на уровне типа. [38] [39] Подобная функциональная совместимость наблюдалась и у Thermus thermophilus . [40] Кроме того, у T. thermophilus наблюдался как полный, так и частичный перенос генов. Частичный перенос привел к спонтанному образованию явно случайных химер между генами хозяина и чужеродных бактериальных генов. Таким образом, гены 16S рРНК могли развиваться посредством множества механизмов, включая вертикальное наследование и горизонтальный перенос генов ; частота последних может быть намного выше, чем считалось ранее. [41]
Базы данных 16S рибосом [ править ]
Ген 16S рРНК используется в качестве стандарта для классификации и идентификации микробов, поскольку он присутствует у большинства микробов и демонстрирует соответствующие изменения. [42] Типовые штаммы последовательностей генов 16S рРНК для большинства бактерий и архей доступны в общедоступных базах данных, таких как NCBI . Однако качество последовательностей, найденных в этих базах данных, часто не проверяется. Поэтому широко используются вторичные базы данных, в которых собраны только последовательности 16S рРНК. Ниже перечислены наиболее часто используемые базы данных:
МИМт [ править ]
MIMt — это компактная неизбыточная база данных 16S для быстрой идентификации метагеномных образцов. Он состоит из 39 940 полных последовательностей 16S, принадлежащих 17 625 хорошо классифицированным видам бактерий и архей. Все последовательности были получены из полных геномов, депонированных в NCBI, и для каждой из последовательностей представлена полная таксономическая иерархия. Он не содержит избыточности, поэтому рассматривался только один представитель для каждого вида, избегая одинаковых последовательностей из разных штаммов, изолятов или патоваров, что привело к созданию очень быстрого инструмента для идентификации микроорганизмов, совместимого с любым классификационным программным обеспечением (QIIME, Mothur, DADA и т. д.). [43]
EzBioCloud [ править ]
База данных EzBioCloud, ранее известная как EzTaxon , состоит из полной иерархической таксономической системы, содержащей 62 988 видов/филотипов бактерий и архей, которая включает 15 290 действительных опубликованных названий по состоянию на сентябрь 2018 года. На основе филогенетических отношений, таких как максимальное правдоподобие и OrthoANI, все виды/ подвиды представлены по крайней мере одной последовательностью гена 16S рРНК. База данных EzBioCloud систематически обновляется и включает в себя новые виды-кандидаты. Кроме того, на веб-сайте представлены инструменты биоинформатики, такие как калькулятор ANI, база данных рРНК ContEst16S и 16S для QIIME и конвейера Mothur. [44] ^^
рибосомальной Проект данных базы
Проект рибосомальной базы данных (RDP) — это курируемая база данных, которая предлагает данные о рибосомах, а также соответствующие программы и услуги. Предложения включают филогенетически упорядоченные выравнивания последовательностей рибосомальных РНК (рРНК), производные филогенетические деревья, диаграммы вторичной структуры рРНК и различные пакеты программного обеспечения для обработки, анализа и отображения выравниваний и деревьев. Данные доступны по FTP и электронной почте. Некоторые аналитические услуги также предоставляются сервером электронной почты. [45] Из-за своего большого размера база данных RDP часто используется в качестве основы для разработки биоинформатических инструментов и создания баз данных, курируемых вручную. [46]
SILVA[editСИЛЬВА
SILVA предоставляет комплексные, проверенные на качество и регулярно обновляемые наборы данных выровненных последовательностей малых (16S/ 18S , SSU ) и больших субъединиц ( 23S / 28S , LSU ) рибосомальных РНК (рРНК) для всех трех областей жизни, а также набор средств для поиска, инструменты для создания праймеров и выравнивания (Бактерии, Археи и Эукарии). [47]
Зеленые Гены [ править ]
GreenGenes — это комплексная справочная база данных генов 16S рРНК с контролируемым качеством и таксономия, основанная на филогении de novo , которая предоставляет стандартные рабочие наборы таксономических единиц. Помните, что здесь используются таксономические термины, предложенные на основе филогенетических методов, применявшихся много лет назад, в период с 2012 по 2013 год. С тех пор для архей и бактерий было предложено множество новых филогенетических методов. [48] [49]
Ссылки [ править ]
- ^ Шлюенцен Ф., Тоцил А., Заривач Р., Хармс Дж., Глюманн М., Джанелл Д. и др. (сентябрь 2000 г.). «Структура функционально активированной малой рибосомальной субъединицы с разрешением 3,3 ангстрема» . Клетка . 102 (5): 615–623. дои : 10.1016/S0092-8674(00)00084-2 . ПМИД 11007480 . S2CID 1024446 .
- ^ Jump up to: Перейти обратно: а б Woese CR , Fox GE (ноябрь 1977 г.). «Филогенетическая структура прокариотического домена: первичные царства» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (11): 5088–5090. Бибкод : 1977PNAS...74.5088W . дои : 10.1073/pnas.74.11.5088 . ПМК 432104 . ПМИД 270744 .

- ^ Вёзе Ч.Р. , Кандлер О., Уилис М.Л. (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–4579. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Дело Р.Дж., Баучер И., Даллёф И., Хольмстрем С., Дулиттл В.Ф., Кьеллеберг С. (январь 2007 г.). «Использование генов 16S рРНК и rpoB в качестве молекулярных маркеров для исследований микробной экологии» . Прикладная и экологическая микробиология . 73 (1): 278–288. Бибкод : 2007ApEnM..73..278C . дои : 10.1128/АЕМ.01177-06 . ПМК 1797146 . ПМИД 17071787 .
- ^ Черниловский А.П., Курланд К.Г. , Штёффлер Г. (октябрь 1975 г.). «30S рибосомальные белки, связанные с 3'-концом 16S РНК» . Письма ФЭБС . 58 (1): 281–284. Бибкод : 1975FEBSL..58..281C . дои : 10.1016/0014-5793(75)80279-1 . ПМИД 1225593 . S2CID 22941368 .
- ^ Вёзе CR (июнь 1987 г.). «Бактериальная эволюция» . Микробиологические обзоры . 51 (2): 221–271. дои : 10.1128/MR.51.2.221-271.1987 . ПМК 373105 . ПМИД 2439888 .
- ^ Jump up to: Перейти обратно: а б с Вайсбург В.Г., Барнс С.М., Пеллетье Д.А., Лейн DJ (январь 1991 г.). «Амплификация рибосомальной ДНК 16S для филогенетических исследований» . Журнал бактериологии . 173 (2): 697–703. дои : 10.1128/jb.173.2.697-703.1991 . ПМК 207061 . ПМИД 1987160 .
- ^ Jump up to: Перейти обратно: а б Коэнье Т., Вандамм П. (ноябрь 2003 г.). «Внутригеномная гетерогенность между несколькими оперонами 16S рибосомальной РНК в секвенированных бактериальных геномах» . Письма FEMS по микробиологии . 228 (1): 45–49. дои : 10.1016/S0378-1097(03)00717-1 . ПМИД 14612235 .
- ^ Цукуда М., Китахара К., Миядзаки К. (август 2017 г.). «Сравнительный анализ функций РНК выявил высокое функциональное сходство между отдаленно родственными бактериальными 16S рРНК» . Научные отчеты . 7 (1): 9993. Бибкод : 2017НатСР...7.9993Т . дои : 10.1038/s41598-017-10214-3 . ПМЦ 5577257 . ПМИД 28855596 .
- ^ Джей ЗиДжей, Inskeep WP (июль 2015 г.). «Распределение, разнообразие и важность интронов гена 16S рРНК в отряде Thermoproteales» . Биология Директ . 10 (35): 35. дои : 10.1186/s13062-015-0065-6 . ПМЦ 4496867 . ПМИД 26156036 .
- ^ Уокер, Сидни П.; Барретт, Морис; Хоган, Гленн; Флорес Буэсо, Йенси; Классон, Маркус Дж.; Тэнгни, Марк (01 октября 2020 г.). «Неспецифическая амплификация ДНК человека является серьезной проблемой для анализа последовательности гена 16S рРНК» . Научные отчеты . 10 (1): 16356. doi : 10.1038/s41598-020-73403-7 . ISSN 2045-2322 . ПМЦ 7529756 . ПМИД 33004967 .
- ^ «Проект микробиома человека DACC - Дом» . www.hmpdacc.org . Архивировано из оригинала 30 октября 2010 г.
- ^ Jump up to: Перейти обратно: а б «Праймеры, рибосомальная ДНК 16S - Лаборатория Франсуа Луцони» . lutzonilab.net . Архивировано из оригинала 27 декабря 2012 г.
- ^ Jump up to: Перейти обратно: а б Иден П.А., Шмидт Т.М., Блейкмор Р.П., Пейс Н.Р. (апрель 1991 г.). «Филогенетический анализ Aquaspirillum Magneticotacticum с использованием ДНК, специфичной для 16S рРНК, амплифицированной полимеразной цепной реакцией» . Международный журнал систематической бактериологии . 41 (2): 324–325. дои : 10.1099/00207713-41-2-324 . ПМИД 1854644 .
- ^ Jump up to: Перейти обратно: а б Джеймс, Грег (15 мая 2018 г.). «Универсальная идентификация бактерий с помощью ПЦР и секвенирования ДНК гена 16S рРНК». ПЦР для клинической микробиологии . Спрингер, Дордрехт. стр. 209–214. дои : 10.1007/978-90-481-9039-3_28 . ISBN 978-90-481-9038-6 .
- ^ Jump up to: Перейти обратно: а б Вайднер С., Арнольд В., Пулер А. (март 1996 г.). «Разнообразие некультивируемых микроорганизмов, связанных с морской травой Halophila stipulacea, оценено с помощью анализа полиморфизма длины рестрикционных фрагментов генов 16S рРНК, амплифицированных ПЦР» (PDF) . Прикладная и экологическая микробиология . 62 (3): 766–771. Бибкод : 1996ApEnM..62..766W . дои : 10.1128/АЕМ.62.3.766-771.1996 . ПМК 167844 . ПМИД 8975607 . Архивировано (PDF) из оригинала 15 июля 2011 г.
- ^ Парк, Чанву; Ким, Сын Бом; Чой, Сан Хо; Ким, Сейл (2021). «Сравнение микробного профилирования на основе гена 16S рРНК с использованием пяти секвенаторов следующего поколения и различных праймеров» . Границы микробиологии . 12 . дои : 10.3389/fmicb.2021.715500 . ISSN 1664-302X . ПМК 8552068 . ПМИД 34721319 .
- ^ Элоэ-Фадрош Е.А., Иванова Н.Н., Войк Т., Кирпидес Н.К. (февраль 2016 г.). «Метагеномика обнаруживает пробелы в обнаружении микробного разнообразия на основе ампликонов». Природная микробиология . 1 (4): 15032. doi : 10.1038/nmicrobiol.2015.32 . ОСТИ 1379258 . ПМИД 27572438 . S2CID 27232975 .
- ^ Бергманн Г.Т., Бейтс С.Т., Эйлерс К.Г., Лаубер К.Л., Капорасо Дж.Г., Уолтерс В.А. и др. (июль 2011 г.). «Недооцененное доминирование Verrucomicrobia в почвенных бактериальных сообществах» . Биология и биохимия почвы . 43 (7): 1450–1455. Бибкод : 2011SBiBi..43.1450B . doi : 10.1016/j.soilbio.2011.03.012 . ПМК 3260529 . ПМИД 22267877 .
- ^ Цзян Х., Донг Х., Чжан Г., Ю Б., Чепмен Л.Р., Филдс М.В. (июнь 2006 г.). «Микробное разнообразие в воде и отложениях озера Чака, аталассохалинного озера на северо-западе Китая» . Прикладная и экологическая микробиология . 72 (6): 3832–3845. Бибкод : 2006ApEnM..72.3832J . дои : 10.1128/АЕМ.02869-05 . ПМЦ 1489620 . ПМИД 16751487 .
- ^ Перейра Ф, Карнейру Дж, Маттисен Р, ван Аш Б, Пинту Н, Гужман Л, Аморим А (декабрь 2010 г.). «Идентификация видов методом мультиплексного анализа последовательностей переменной длины» . Исследования нуклеиновых кислот . 38 (22): е203. дои : 10.1093/nar/gkq865 . ПМК 3001097 . ПМИД 20923781 .
- ^ Кольберт КП, Персинг Д.Х. (июнь 1999 г.). «Секвенирование рибосомальной ДНК как инструмент идентификации бактериальных патогенов». Современное мнение в микробиологии . 2 (3): 299–305. дои : 10.1016/S1369-5274(99)80052-6 . ПМИД 10383862 .
- ^ Кларидж Дж. Э. (октябрь 2004 г.). «Влияние анализа последовательности гена 16S рРНК для идентификации бактерий на клиническую микробиологию и инфекционные заболевания» . Обзоры клинической микробиологии . 17 (4): 840–62, оглавление. doi : 10.1128/CMR.17.4.840-862.2004 . ПМК 523561 . ПМИД 15489351 .
- ^ Лу Т, Строот П.Г., Оертер Д.Б. (июль 2009 г.). «Обратная транскрипция 16S рРНК для мониторинга популяций бактерий, синтезирующих рибосомы, в окружающей среде» . Прикладная и экологическая микробиология . 75 (13): 4589–4598. Бибкод : 2009ApEnM..75.4589L . дои : 10.1128/АЕМ.02970-08 . ПМК 2704851 . ПМИД 19395563 .
- ^ Бретт П.Дж., ДеШазер Д., Вудс Д.Э. (январь 1998 г.). «Burkholderia thailandensis sp. nov., вид, похожий на Burkholderia pseudomallei» . Международный журнал систематической бактериологии . 48 Ч. 1 (1): 317–320. дои : 10.1099/00207713-48-1-317 . ПМИД 9542103 .
- ^ Шмидт Т.М., Рельман Д.А. (1994). «Филогенетическая идентификация некультивируемых патогенов с использованием последовательностей рибосомальной РНК» . Бактериальный патогенез. Часть A: Идентификация и регулирование факторов вирулентности . Методы энзимологии. Том. 235. С. 205–222 . дои : 10.1016/0076-6879(94)35142-2 . ISBN 978-0-12-182136-4 . ПМИД 7520119 .
- ^ Грей Дж.П., Хервиг Р.П. (ноябрь 1996 г.). «Филогенетический анализ бактериальных сообществ морских отложений» . Прикладная и экологическая микробиология . 62 (11): 4049–4059. Бибкод : 1996ApEnM..62.4049G . дои : 10.1128/АЕМ.62.11.4049-4059.1996 . ПМК 168226 . ПМИД 8899989 .
- ^ Саншагрин С., Йерго Э. (август 2014 г.). «Секвенирование следующего поколения ампликонов гена рибосомальной РНК 16S» . Журнал визуализированных экспериментов (90). дои : 10.3791/51709 . ПМЦ 4828026 . ПМИД 25226019 .
- ^ Ботан, Александру; Кампишано, Джузеппина; Зербато, Верена; Ди Белла, Стефано; Симонетти, Омар; Бусетти, Марина; Ток, Дэн Александру; Луццати, Роберто; Комар, Манола (21 июня 2024 г.). «Эффективность секвенирования гена 16S рРНК следующего поколения и метода культивирования при обнаружении бактерий в клинических образцах» . Диагностика . 14 (13): 1318. doi : 10.3390/diagnostics14131318 . ISSN 2075-4418 .
- ^ Грей М.В., Санкофф Д., Седергрен Р.Дж. (июль 1984 г.). «Об эволюционном происхождении организмов и органелл: глобальная филогения, основанная на высококонсервативном структурном ядре небольшой субъединицы рибосомальной РНК» . Исследования нуклеиновых кислот . 12 (14): 5837–5852. дои : 10.1093/нар/12.14.5837 . ПМК 320035 . ПМИД 6462918 .
- ^ Jump up to: Перейти обратно: а б с д Ян Б., Ван Ю, Цянь П.Ю. (март 2016 г.). «Чувствительность и корреляция гипервариабельных участков генов 16S рРНК в филогенетическом анализе» . БМК Биоинформатика . 17 (1): 135. дои : 10.1186/s12859-016-0992-y . ПМЦ 4802574 . ПМИД 27000765 .
- ^ Бартрам А.К., Линч, доктор медицинских наук, Стернс Дж.К., Морено-Хагелзиб Г., Нойфельд Дж.Д. (июнь 2011 г.). «Создание библиотек генов 16S рРНК с несколькими миллионами последовательностей из сложных микробных сообществ путем сборки парных концевых прочтений Ilumina» . Прикладная и экологическая микробиология . 77 (11): 3846–3852. Бибкод : 2011ApEnM..77.3846B . дои : 10.1128/АЕМ.02772-10 . ПМК 3127616 . ПМИД 21460107 .
- ^ Jump up to: Перейти обратно: а б Берк CM, Дарлинг AE (20 сентября 2016 г.). «Метод высокоточного секвенирования почти полноразмерных генов 16S рРНК на аппарате Illumina MiSeq» . ПерДж . 4 : е2492. дои : 10.7717/peerj.2492 . ПМК 5036073 . ПМИД 27688981 .
- ^ Ван де Пер Ю., Шапель С., Де Вахтер Р. (сентябрь 1996 г.). «Количественная карта скорости замен нуклеотидов в бактериальной рРНК» . Исследования нуклеиновых кислот . 24 (17): 3381–3391. дои : 10.1093/нар/24.17.3381 . ПМК 146102 . ПМИД 8811093 .
- ^ Jump up to: Перейти обратно: а б с Ветровский Т, Балдриан П (27 февраля 2013 г.). «Изменчивость гена 16S рРНК в бактериальных геномах и ее последствия для анализа бактериального сообщества» . ПЛОС ОДИН . 8 (2): e57923. Бибкод : 2013PLoSO...857923V . дои : 10.1371/journal.pone.0057923 . ПМЦ 3583900 . ПМИД 23460914 .
- ^ Jump up to: Перейти обратно: а б Чакраворти С., Хелб Д., Бердей М., Коннелл Н., Алланд Д. (май 2007 г.). «Детальный анализ сегментов гена 16S рибосомальной РНК для диагностики патогенных бактерий» . Журнал микробиологических методов . 69 (2): 330–339. дои : 10.1016/j.mimet.2007.02.005 . ПМК 2562909 . ПМИД 17391789 .
- ^ Jump up to: Перейти обратно: а б с Джовел Дж., Паттерсон Дж., Ван В., Хотте Н., О'Киф С., Митчел Т. и др. (01.01.2016). «Характеристика кишечного микробиома с использованием 16S или метагеномики дробовика» . Границы микробиологии . 7 : 459. дои : 10.3389/fmicb.2016.00459 . ПМЦ 4837688 . ПМИД 27148170 .
- ^ Китахара К., Ясутаке Ю., Миядзаки К. (ноябрь 2012 г.). «Мутационная устойчивость 16S рибосомальной РНК, показанная экспериментальным горизонтальным переносом генов в Escherichia coli» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (47): 19220–19225. Бибкод : 2012PNAS..10919220K . дои : 10.1073/pnas.1213609109 . ПМК 3511107 . ПМИД 23112186 .
- ^ Цукуда М., Китахара К., Миядзаки К. (август 2017 г.). «Сравнительный анализ функций РНК выявил высокое функциональное сходство между отдаленно родственными бактериальными 16S рРНК» . Научные отчеты . 7 (1): 9993. Бибкод : 2017НатСР...7.9993Т . дои : 10.1038/s41598-017-10214-3 . ПМЦ 5577257 . ПМИД 28855596 .
- ^ Миядзаки К., Томаригути Н. (август 2019 г.). «Появление случайно рекомбинированных функциональных генов 16S рРНК у Thermus thermophilus предполагает генетическую совместимость и беспорядочность бактериальных 16S рРНК» . Научные отчеты . 9 (1): 11233. Бибкод : 2019НатСР...911233М . дои : 10.1038/s41598-019-47807-z . ПМК 6677816 . ПМИД 31375780 .
- ^ Миядзаки, Кентаро; Томаригути, Нацуки (2 августа 2019 г.). «Появление случайно рекомбинированных функциональных генов 16S рРНК у Thermus thermophilus предполагает генетическую совместимость и беспорядочность бактериальных 16S рРНК» . Научные отчеты . 9 (1): 11233. Бибкод : 2019НатСР...911233М . дои : 10.1038/s41598-019-47807-z . ISSN 2045-2322 . ПМК 6677816 . ПМИД 31375780 .
- ^ Ярза П., Йилмаз П., Прюссе Э., Глёкнер Ф.О., Людвиг В., Шлейфер К.Х. и др. (сентябрь 2014 г.). «Объединение классификации культивируемых и некультивируемых бактерий и архей с использованием последовательностей гена 16S рРНК». Обзоры природы. Микробиология . 12 (9): 635–645. дои : 10.1038/nrmicro3330 . ПМИД 25118885 . S2CID 21895693 .
- ^ «МИМт – (Массовая идентификация метагеномных тестов)» . mimt.bu.biopolis.pt . Проверено 11 февраля 2024 г.
- ^ Юн, Ш., Ха, СМ, Квон, С., Лим, Дж., Ким, Ю., Со, Х. и Чун, Дж. (2017). Представляем EzBioCloud: таксономически объединенную базу данных 16S рРНК и целых геномных сборок. Int J Syst Evol Microbiol. 67: 1613–1617
- ^ Ларсен Н., Олсен Г.Дж., Майдак Б.Л., МакКоги М.Дж., Овербик Р., Маке Т.Дж., Марш Т.Л., Вёзе Ч.Р. (1993) Проект рибосомальной базы данных. Нуклеиновые кислоты Рез. 1 июля;21(13):3021-3.
- ^ Аллард Дж., Райан Ф.Дж., Джеффри И.Б., Клаессон М.Дж. (октябрь 2015 г.). «SPINGO: быстрый видовой классификатор последовательностей микробных ампликонов» . БМК Биоинформатика . 16 (1): 324. дои : 10.1186/s12859-015-0747-1 . ПМК 4599320 . ПМИД 26450747 .
- ^ Элмар Прюссе, Кристиан Кваст, Катрин Книттель, Бернхард М. Фукс, Вольфганг Людвиг, Йорг Пеплиес, Фрэнк Оливер Глёкнер (2007) Nucleic Acids Res. SILVA: комплексный онлайн-ресурс для проверенных и согласованных по качеству данных о последовательностях рибосомальных РНК, совместимых с ARB. декабрь; 35(21): 7188–7196.
- ^ ДеСантис Т.З., Хугенхольц П., Ларсен Н., Рохас М., Броди Э.Л., Келлер К. и др. (июль 2006 г.). «Greengenes, проверенная химерами база данных генов 16S рРНК и инструментальные средства, совместимые с ARB» . Прикладная и экологическая микробиология . 72 (7): 5069–5072. Бибкод : 2006ApEnM..72.5069D . дои : 10.1128/aem.03006-05 . ПМЦ 1489311 . ПМИД 16820507 .
- ^ Макдональд Д., Прайс М.Н., Гудрич Дж., Навроцкий Е.П., ДеСантис Т.З., Пробст А. и др. (март 2012 г.). «Улучшенная таксономия Greengenes с четкими рангами для экологического и эволюционного анализа бактерий и архей» . Журнал ISME . 6 (3): 610–618. Бибкод : 2012ISMEJ...6..610M . дои : 10.1038/ismej.2011.139 . ПМК 3280142 . ПМИД 22134646 .
Внешние ссылки [ править ]
- Лабораторная медицина Вашингтонского университета: молекулярная диагностика | Бактериальное секвенирование
- База данных МИМт 16С
- Проект рибосомальной базы данных , заархивированный 19 августа 2020 г. на Wayback Machine.
- Рибосомы и рибосомальная РНК: (рРНК)
- База данных рРНК SILVA
- Зеленые гены: данные и инструменты по 16S рДНК
- EzBioCloud