Экзосомный комплекс

Экзосомальный комплекс (или комплекс PM/Scl , часто называемый просто экзосомой ) представляет собой многобелковый внутриклеточный комплекс , способный расщеплять различные типы молекул РНК (рибонуклеиновой кислоты). Экзосомальные комплексы обнаружены как в эукариотических клетках, так и в археях , тогда как у бактерий более простой комплекс, называемый деградосомой, выполняет аналогичные функции.
Ядро экзосомы содержит шестичленную кольцевую структуру, к которой прикреплены другие белки. В эукариотических клетках экзосомный комплекс присутствует в цитоплазме , ядре и особенно в ядрышке , хотя различные белки взаимодействуют с экзосомным комплексом в этих компартментах, регулируя активность деградации РНК комплекса до субстратов, специфичных для этих клеточных компартментов. Субстраты экзосомы включают информационную РНК , рибосомальную РНК и многие виды малых РНК . Экзосома выполняет экзорибонуклеолитическую функцию, то есть она расщепляет РНК, начиная с одного конца ( в данном случае 3'-конца ), а у эукариот также выполняет эндорибонуклеолитическую функцию, то есть расщепляет РНК в участках внутри молекулы.
Некоторые белки в экзосоме являются мишенью аутоантител у пациентов со специфическими аутоиммунными заболеваниями (особенно синдромом перекрытия PM/Scl ), а некоторые антиметаболические химиотерапевтические препараты при раке действуют путем блокирования активности экзосомы. Кроме того, мутации в компоненте 3 экзосомы вызывают понтоцеребеллярную гипоплазию и заболевание мотонейронов спинного мозга .
Открытие
[ редактировать ]Экзосома была впервые обнаружена как РНКаза в 1997 году в почкующихся дрожжах Saccharomyces cerevisiae , часто используемом модельном организме . [1] Вскоре после этого, в 1999 году, стало понятно, что экзосома на самом деле представляет собой дрожжевой эквивалент уже описанного комплекса в клетках человека, называемого комплексом PM/Scl был идентифицирован как аутоантиген у пациентов с некоторыми аутоиммунными заболеваниями , который несколько лет назад ( см . ниже ). [2] Очистка этого «комплекса PM/Scl» позволила идентифицировать больше белков экзосом человека и, в конечном итоге, охарактеризовать все компоненты комплекса. [3] [4] В 2001 году растущее количество данных о геноме , которые стали доступны, позволило предсказать белки экзосом у архей, хотя потребовалось еще два года, прежде чем был очищен первый комплекс экзосом из архейного организма. [5] [6]
Структура
[ редактировать ]Основные белки
[ редактировать ]
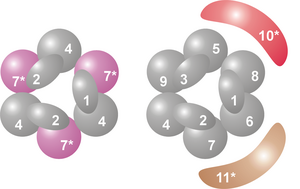
Ядро комплекса имеет кольцевую структуру, состоящую из шести белков, принадлежащих к одному классу РНКаз — РНКазным PH- подобным белкам. [7] У архей есть два разных PH-подобных белка (называемых Rrp41 и Rrp42), каждый из которых присутствует три раза в чередующемся порядке. Эукариотические экзосомальные комплексы содержат шесть различных белков, образующих кольцевую структуру. [8] [9] Из этих шести эукариотических белков три напоминают архейный белок Rrp41, а три других белка больше похожи на архейный белок Rrp42. [10]

На вершине этого кольца расположены три белка, которые имеют РНК-связывающий домен S1 (RBD). Два белка дополнительно имеют домен K-гомологии (KH) . [7] У эукариот с кольцом связаны три разных белка «S1», тогда как у архей в состав экзосомы могут входить либо один, либо два разных белка «S1» (хотя к комплексу всегда присоединены три субъединицы S1). [11]
Эта кольцевая структура очень похожа на структуру белков РНКазы PH и PNPase . У бактерий белок РНКаза PH, участвующий в процессинге тРНК , образует гексамерное кольцо, состоящее из шести идентичных белков РНКазы PH. [12] [13] В случае PNPазы, которая представляет собой фосфоролитический белок, расщепляющий РНК, обнаруженный в бактериях , хлоропластах и митохондриях некоторых эукариотических организмов, два домена РНКазы PH, а также РНК-связывающие домены S1 и KH являются частью одного белка, который образует тримерный комплекс, имеющий структуру, почти идентичную структуре экзосомы. [14] Из-за такого высокого сходства как белковых доменов, так и структуры считается, что эти комплексы эволюционно родственны и имеют общего предка . [15] семейству РНКаз РНКазы PH и являются фосфоролитическими экзорибонуклеазами , что означает, что они используют неорганический фосфат для удаления нуклеотидов с 3'-конца молекул РНК Белки экзосомы, подобные РНКазе PH, PNPase и RNase PH принадлежат к . [7]
Связанные белки
[ редактировать ]Помимо этих девяти основных белков экзосомы, в эукариотических организмах с комплексом часто связаны два других белка. Одним из них является Rrp44 , гидролитическая РНКаза, которая принадлежит к РНКазы R семейству гидролитических экзорибонуклеаз (нуклеаз, которые используют воду для расщепления нуклеотидных связей). Помимо того, что Rrp44 является экзорибонуклеолитическим ферментом, он также обладает эндорибонуклеолитической активностью, которая находится в отдельном домене белка. [16] [17] У дрожжей Rrp44 связан со всеми экзосомными комплексами и играет решающую роль в активности экзосомного комплекса дрожжей. [18] Хотя человеческий гомолог белка существует, в течение длительного времени не было обнаружено доказательств того, что его человеческий гомолог связан с экзосомным комплексом человека. [7] Однако в 2010 году было обнаружено, что у человека есть три гомолога Rrp44, и два из них могут быть связаны с экзосомным комплексом. Эти два белка, скорее всего, разрушают разные субстраты РНК из-за их разной клеточной локализации: один локализован в цитоплазме ( DIS3L1 ), а другой — в ядре ( DIS3 ). [19] [20]

Второй распространенный ассоциированный белок называется Rrp6 (у дрожжей) или PM/Scl-100 (у человека). Как и Rrp44, этот белок представляет собой гидролитическую экзорибонуклеазу, но в данном случае относится к РНКазы D. семейству белков [21] Белок PM/Scl-100 чаще всего является частью экзосомных комплексов в ядре клеток, но также может составлять часть цитоплазматического экзосомного комплекса. [22]
Регуляторные белки
[ редактировать ]Помимо этих двух прочно связанных белковых субъединиц, многие белки взаимодействуют с экзосомным комплексом как в цитоплазме, так и в ядре клеток. Эти слабо связанные белки могут регулировать активность и специфичность экзосомального комплекса. В цитоплазме экзосома взаимодействует со связывающими белками, богатыми AU-элементами (ARE) (например, KRSP и TTP), которые могут способствовать или предотвращать деградацию мРНК. Ядерная экзосома связана с РНК-связывающими белками (например, MPP6/Mpp6 и C1D/Rrp47 у человека/дрожжей), которые необходимы для обработки определенных субстратов. [7]
Помимо одиночных белков, с экзосомой взаимодействуют и другие белковые комплексы. Одним из них является цитоплазматический комплекс Ski , который включает РНК- геликазу (Ski2) и участвует в деградации мРНК. [23] В ядре процессинг рРНК и мякРНК экзосомой опосредован комплексом TRAMP , который обладает как РНК-хеликазной (Mtr4), так и полиаденилирующей (Trf4) активностью. [24]
Функция
[ редактировать ]Ферментативная функция
[ редактировать ]
Как указано выше, экзосомальный комплекс содержит множество белков с рибонуклеазными доменами. Точная природа этих рибонуклеазных доменов менялась в ходе эволюции от бактериальных комплексов к архейным и эукариотическим, поскольку различные виды активности приобретались и терялись. Экзосома представляет собой в первую очередь 3'-5' экзорибонуклеазу , что означает, что она расщепляет молекулы РНК с их 3'-конца . Экзорибонуклеазы, содержащиеся в экзосомных комплексах, являются либо фосфоролитическими (РНКазы PH-подобные белки), либо, у эукариот, гидролитическими (белки доменов РНКазы R и РНКазы D). Фосфоролитические ферменты используют неорганический фосфат для расщепления фосфодиэфирных связей , высвобождая нуклеотиддифосфаты . Гидролитические ферменты используют воду для гидролиза этих связей, высвобождая нуклеотидмонофосфаты .
У архей субъединица комплекса Rrp41 представляет собой фосфоролитическую экзорибонуклеазу. В кольце присутствуют три копии этого белка, отвечающие за активность комплекса. [9] У эукариот ни одна из субъединиц РНКазы PH не сохранила эту каталитическую активность, а это означает, что основная кольцевая структура человеческой экзосомы не имеет ферментативно активного белка. [25] Несмотря на эту потерю каталитической активности, структура сердцевинной экзосомы высоко консервативна от архей до человека, что позволяет предположить, что комплекс выполняет жизненно важную клеточную функцию. У эукариот отсутствие фосфоролитической активности компенсируется наличием гидролитических ферментов, ответственных за рибонуклеазную активность экзосомы у таких организмов. [26] [27] [28]
Как указано выше, гидролитические белки Rrp6 и Rrp44 связаны с экзосомой у дрожжей, а у человека, кроме Rrp6, в положении дрожжевого белка Rrp44 могут быть ассоциированы два разных белка, Dis3 и Dis3L1. [19] [20] Хотя первоначально считалось, что белки домена S1 также обладают 3'-5'-гидролитической экзорибонуклеазной активностью, существование этой активности недавно было поставлено под сомнение, и эти белки могли играть лишь роль в связывании субстратов до их деградации комплексом. [26]
Субстраты
[ редактировать ]Экзосома участвует в деградации и процессинге широкого спектра видов РНК. В цитоплазме клеток он участвует в обмене молекул информационной РНК (мРНК). Комплекс может разрушать молекулы мРНК, которые были помечены для деградации, поскольку они содержат ошибки, посредством взаимодействия с белками по бессмысленному или безостановочному распаду . Альтернативно, мРНК деградируют в ходе своего нормального оборота . Несколько белков, которые стабилизируют или дестабилизируют молекулы мРНК посредством связывания с AU-богатыми элементами в 3'-нетранслируемой области мРНК, взаимодействуют с экзосомным комплексом. [29] [30] [31] В ядре экзосома необходима для правильного процессинга нескольких небольших молекул ядерной РНК. [32] Наконец, ядрышко — это отсек, в котором находится большинство экзосомальных комплексов. Там он играет роль в процессинге 5.8S рибосомальной РНК (первая выявленная функция экзосомы) и нескольких малых ядрышковых РНК . [1] [32] [33]
Хотя в большинстве клеток есть и другие ферменты, которые могут расщеплять РНК либо с 3'- , либо с 5'-конца РНК, экзосомный комплекс необходим для выживания клеток. Когда экспрессия белков экзосом искусственно снижается или прекращается, например, с помощью РНК-интерференции , рост прекращается и клетки в конечном итоге погибают. Оба основных белка экзосомального комплекса, а также два основных ассоциированных белка являются незаменимыми белками. [34] Бактерии не имеют экзосомного комплекса; однако аналогичные функции выполняет более простой комплекс, включающий белок ПНПазу , называемый деградосомой . [35]

Экзосома является ключевым комплексом в контроле качества клеточной РНК. В отличие от прокариот, эукариоты обладают высокоактивными системами наблюдения за РНК, которые распознают непроцессированные и неправильно процессированные комплексы РНК-белок (такие как рибосомы ) до их выхода из ядра. Предполагается, что эта система предотвращает вмешательство аберрантных комплексов в важные клеточные процессы, такие как синтез белка . [36]
Помимо процессинга, оборота и надзора за РНК, экзосома важна для деградации так называемых загадочных нестабильных транскриптов (CUT), которые производятся из тысяч локусов в геноме дрожжей. [37] [38] Важность этих нестабильных РНК и их деградация до сих пор неясны, но подобные виды РНК также были обнаружены в клетках человека. [39]
Болезнь
[ редактировать ]Аутоиммунитет
[ редактировать ]Экзосомальный комплекс является мишенью аутоантител у пациентов с различными аутоиммунными заболеваниями . Эти аутоантитела в основном обнаруживаются у людей с синдромом перекрытия PM/Scl , аутоиммунным заболеванием, при котором у пациентов наблюдаются симптомы как склеродермии , так и полимиозита или дерматомиозита . [40] Аутоантитела можно обнаружить в сыворотке пациентов с помощью различных анализов. В прошлом наиболее часто используемыми методами были двойная иммунодиффузия теленка с использованием экстрактов тимуса , иммунофлуоресценция на клетках HEp-2 или иммунопреципитация из экстрактов клеток человека. В анализах иммунопреципитации с сыворотками из антиэкзосомо-положительных сывороток осаждается особый набор белков. Уже за несколько лет до того, как экзосомный комплекс был идентифицирован, этот паттерн был назван комплексом PM/Scl . [41] Иммунофлуоресценция сыворотки этих пациентов обычно показывает типичное окрашивание ядрышка клеток , что вызвало предположение о том, что антиген, распознаваемый аутоантителами, может играть важную роль в синтезе рибосом . [42] Совсем недавно стали доступны рекомбинантные белки экзосом, и они были использованы для разработки линейных иммуноанализов (LIA) и иммуноферментных анализов (ELISA) для обнаружения этих антител. [7]
При этих заболеваниях антитела в основном направлены против двух белков комплекса, называемых PM/Scl-100 (РНКаза D-подобный белок) и PM/Scl-75 (один из РНКазы PH-подобных белков из кольца), а также антитела. распознающие эти белки обнаруживаются примерно у 30% пациентов с синдромом перекрытия PM/Scl. [43] Хотя эти два белка являются основной мишенью аутоантител, у этих пациентов могут быть нацелены и другие субъединицы экзосом и связанные с ними белки (например, C1D). [44] [45] В настоящее время наиболее чувствительным способом обнаружения этих антител является использование пептида , полученного из белка PM/Scl-100, в качестве антигена в ELISA вместо полноценных белков. С помощью этого метода аутоантитела обнаруживаются до 55% пациентов с синдромом перекрытия PM/Scl, но их также можно обнаружить у пациентов с только склеродермией, полимиозитом или дерматомиозитом. [46]
Поскольку аутотела обнаруживаются преимущественно у пациентов с характеристиками нескольких различных аутоиммунных заболеваний, клинические симптомы этих пациентов могут широко варьировать. Симптомы, которые наблюдаются чаще всего, являются типичными симптомами отдельных аутоиммунных заболеваний и включают феномен Рейно , артрит , миозит и склеродермию . [47] Лечение этих пациентов симптоматическое и аналогично лечению отдельного аутоиммунного заболевания, часто с применением иммунодепрессантов или иммуномодулирующих препаратов. [48]
Лечение рака
[ редактировать ]что экзосома ингибируется антиметаболитом фторурацилом , препаратом, используемым при химиотерапии рака Было показано , . Это один из наиболее успешных препаратов для лечения солидных опухолей . В дрожжевых клетках, обработанных фторурацилом, были обнаружены дефекты процессинга рибосомальной РНК, идентичные тем, которые наблюдались, когда активность экзосомы блокировалась молекулярно-биологическими стратегиями. Отсутствие правильного процессинга рибосомальной РНК смертельно для клеток, что объясняет антиметаболический эффект препарата. [49]
Неврологические расстройства
[ редактировать ]Мутации в экзосомном компоненте 3 вызывают инфантильную болезнь спинальных мотонейронов , атрофию мозжечка, прогрессирующую микроцефалию и глубокую глобальную задержку развития, что соответствует понтоцеребеллярной гипоплазии типа 1B ( PCH1B; MIM 614678 ). [50]
Список субъединиц
[ редактировать ]| Легенда | Общее имя | Домены | Человек | Дрожжи ( S. cerevisiae ) | Архея | МВт (кД) | Человеческий ген | Дрожжевой ген |
|---|---|---|---|---|---|---|---|---|
| 1 | Csl4 | С1 РБД | hCsl4 | Csl4p/Ski4p | Csl4 | 21–32 | EXOSC1 | YNL232W |
| 2 | Ррп4 | С1/КХ РБД | HRRP4 | Ррп4п | Ррп4 | 28–39 | EXOSC2 | YHR069C |
| 3 | 40 рупий | С1/КХ РБД | чРрп40 | Цена за 40 пенсов | (Ррп4) А | 27–32 | EXOSC3 | YOL142W |
| 4 | Ррп41 | РНКаза PH | HRRP41 | Цена 41p/Ski6p | Ррп41 С | 26–28 | EXOSC4 | YGR195W |
| 5 | Ррп46 | РНКаза PH | hrrp46 | Ррп46п | (Ррп41) А, С | 25–28 | EXOSC5 | YGR095C |
| 6 | Мтр3 | РНКаза PH | hMtr3 | Мтр3п | (Ррп41) А, С | 24–37 | EXOSC6 | YGR158C |
| 7 | Ррп42 | РНКаза PH | HRRP42 | Ррп42п | Ррп42 | 29–32 | EXOSC7 | YDL111C |
| 8 | Ррп43 | РНКаза PH | ОИП2 | Ррп43п | (Ррп42) А | 30–44 | EXOSC8 | YCR035C |
| 9 | Цена за 45 рублей | РНКаза PH | ПМ/Скл-75 | Цена за 45 пенсов | (Ррп42) А | 34–49 | EXOSC9 | 280 Вт ярдов |
| 10 | Ррп6 | РНКаза D | ПМ/Скл-100 С | Ррп6п С | н/д | 84–100 | EXOSC10 | YOR001W |
| 11 | Ррп44 | РНКаза R | Дис3 Б, С | Ррп44п/Dis3p С | н/д | 105–113 | ДИС3 | YOL021C |
- А У архей несколько белков экзосом присутствуют в нескольких копиях, образуя полное ядро экзосомного комплекса.
- Б У человека в этом положении могут быть связаны два разных белка. В цитоплазме клеток Dis3L1 связан с экзосомой, тогда как в ядре Dis3 может связываться с коровым комплексом.
- С Способствует рибонуклеолитической активности комплекса.
См. также
[ редактировать ]- Протеасома . — основной механизм клеточного расщепления белков
- Сплайсосома — комплекс, участвующий в сплайсинге РНК , который также содержит кольцевую структуру, связывающую РНК.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Митчелл, П; Петфальский, Э; Шевченко А; Манн, М; Толлерви, Д. (1997). «Экзосома: консервативный эукариотический комплекс процессинга РНК, содержащий несколько 3'→5'-экзорибонуклеаз» . Клетка . 91 (4): 457–466. дои : 10.1016/S0092-8674(00)80432-8 . ПМИД 9390555 . S2CID 16035676 .
- ^ Оллманг, К; Петфальский, Э; Подтелейников А; Манн, М; Толлерви, Д; Митчелл, П. (1999). «Экзосома дрожжей и PM-Scl человека представляют собой родственные комплексы 3' → 5' экзонуклеаз» . Гены и развитие . 13 (16): 2148–58. дои : 10.1101/gad.13.16.2148 . ПМК 316947 . ПМИД 10465791 .
- ^ Брюэр, Р.; Оллманг, К; Райджмейкерс, Р; Ван Арссен, Ю; Эгбертс, Западная Вирджиния; Петфальский, Э; Ван Венрой, WJ; Толлерви, Д; Пруейн, Дж.Дж. (2001). «Три новых компонента экзосомы человека» . Журнал биологической химии . 276 (9): 6177–84. дои : 10.1074/jbc.M007603200 . hdl : 2066/186951 . ПМИД 11110791 .
- ^ Чен, CY; Герци, Р; Онг, ЮВ; Чан, Эл; Райджмейкерс, Р; Пруейн, Г.Дж.; Стеклин, Г; Мороний, К; и др. (2001). «AU-связывающие белки привлекают экзосому для разрушения ARE-содержащих мРНК» . Клетка . 107 (4): 451–64. дои : 10.1016/S0092-8674(01)00578-5 . ПМИД 11719186 . S2CID 14817671 .
- ^ Кунин Е.В. ; Вольф, Ю.И.; Аравинд, Л. (2001). «Прогнозирование экзосомы архей и ее связей с протеасомой, а также механизмами трансляции и транскрипции с помощью сравнительно-геномного подхода» . Геномные исследования . 11 (2): 240–52. дои : 10.1101/гр.162001 . ПМК 311015 . ПМИД 11157787 .
- ^ Евгеньева-Хакенберг, Э; Уолтер, П; Хохляйтнер, Э; Лотспайх, Ф; Клюг, Г. (2003). «Экзосомоподобный комплекс у Sulfolobus solfataricus» . Отчеты ЭМБО . 4 (9): 889–93. дои : 10.1038/sj.embor.embor929 . ПМЦ 1326366 . ПМИД 12947419 .
- ^ Перейти обратно: а б с д и ж Шильдерс, Г; Ван Дейк, Э; Райджмейкерс, Р; Пруейн, Дж.Дж. (2006). Клеточная и молекулярная биология экзосомы: как создавать или разрушать РНК . Международный обзор цитологии. Том. 251. С. 159–208. дои : 10.1016/S0074-7696(06)51005-8 . ISBN 9780123646552 . ПМИД 16939780 .
- ^ Лоренцен, Э; Уолтер, П; Фрибур, С; Евгеньева-Хакенберг, Э; Клюг, Г; Конти, Э (2005). «Ядро экзосомы архей представляет собой гексамерную кольцевую структуру с тремя каталитическими субъединицами». Структурная и молекулярная биология природы . 12 (7): 575–81. дои : 10.1038/nsmb952 . ПМИД 15951817 . S2CID 2003922 .
- ^ Перейти обратно: а б Шен, В; Киледжян, М (2006). «Взгляд на убийство: структура экзосомы РНК» . Клетка . 127 (6): 1093–5. дои : 10.1016/j.cell.2006.11.035 . ЧВК 1986773 . ПМИД 17174886 .
- ^ Райджмейкерс, Р; Эгбертс, Западная Вирджиния; Ван Венрой, WJ; Пруейн, Дж.Дж. (2002). «Белко-белковые взаимодействия между компонентами экзосом человека поддерживают сборку субъединиц РНКазы PH-типа в шестичленное ПНПазоподобное кольцо». Журнал молекулярной биологии . 323 (4): 653–63. дои : 10.1016/S0022-2836(02)00947-6 . hdl : 2066/186665 . ПМИД 12419256 .
- ^ Уолтер, П; Кляйн, Ф; Лоренцен, Э; Ильхманн, А; Клюг, Г; Евгеньева-Хакенберг, Э (2006). «Характеристика нативных и восстановленных экзосомных комплексов гипертермофильной археи Sulfolobus solfataricus». Молекулярная микробиология . 62 (4): 1076–89. дои : 10.1111/j.1365-2958.2006.05393.x . ПМИД 17078816 . S2CID 27114625 .
- ^ Исии, Р; Нуреки, О; Ёкояма, С. (2003). «Кристаллическая структура фермента процессинга тРНК РНКазы PH из Aquifex aeolicus» . Журнал биологической химии . 278 (34): 32397–404. дои : 10.1074/jbc.M300639200 . ПМИД 12746447 .
- ^ Харлоу, Л.С.; Кадзиола, А; Дженсен, К.Ф.; Ларсен, С. (2004). «Кристаллическая структура фосфоролитической экзорибонуклеазы РНКазы PH из Bacillus subtilis и значение ее четвертичной структуры и связывания тРНК» . Белковая наука . 13 (3): 668–77. дои : 10.1110/ps.03477004 . ПМК 2286726 . ПМИД 14767080 .
- ^ Симмонс, МФ; Джонс, GH; Луизи, БФ (2000). «Дублированная складка является структурной основой каталитической активности, процессивности и регуляции полинуклеотидфосфорилазы» . Структура . 8 (11): 1215–26. дои : 10.1016/S0969-2126(00)00521-9 . ПМИД 11080643 .
- ^ Линь-Чао, С; Чиу, Северная Каролина; Шустер, Г. (2007). «ПНПаза, экзосома и РНК-хеликазы как строительные компоненты эволюционно-консервативных машин деградации РНК» . Журнал биомедицинской науки . 14 (4): 523–32. дои : 10.1007/s11373-007-9178-y . ПМИД 17514363 .
- ^ Лебретон, А; Томецки, Р; Дзембовский, А; Серафин, Б. (2008). «Эндонуклеолитическое расщепление РНК эукариотической экзосомой» (PDF) . Природа . 456 (7224): 993–6. Бибкод : 2008Natur.456..993L . дои : 10.1038/nature07480 . ПМИД 19060886 . S2CID 1808371 .
- ^ Шнайдер, К; Люнг, Э; Браун, Дж; Толлерви, Д. (2009). «N-концевой PIN-домен субъединицы экзосомы Rrp44 обладает эндонуклеазной активностью и привязывает Rrp44 к экзосоме ядра дрожжей» . Исследования нуклеиновых кислот . 37 (4): 1127–40. дои : 10.1093/нар/gkn1020 . ПМК 2651783 . ПМИД 19129231 .
- ^ Шнайдер, К; Андерсон, Джей Ти; Толлерви, Д. (2007). «Субъединица экзосомы Rrp44 играет непосредственную роль в распознавании субстрата РНК» . Молекулярная клетка . 27 (2): 324–31. doi : 10.1016/j.molcel.2007.06.006 . ПМК 7610968 . ПМИД 17643380 .
- ^ Перейти обратно: а б Стаалс, Р.Х.; Бронкхорст, AW; Шильдерс, Г; Сломович, С; Шустер, Г; Черт возьми, Эй Джей; Райджмейкерс, Р; Пруейн, Дж.Дж. (2010). «Dis3-подобный 1: новая экзорибонуклеаза, связанная с экзосомой человека» . Журнал ЭМБО . 29 (14): 2358–67. дои : 10.1038/emboj.2010.122 . ПМК 2910272 . ПМИД 20531389 .
- ^ Перейти обратно: а б Томецки, Р; Кристиансен, MS; Люкке-Андерсен, С; Хлебовский, А; Ларсен, К.М.; Щесны, Р.Дж.; Дражковска, К; Пастула, А; и др. (2010). «Коровая экзосома человека взаимодействует с дифференциально локализованными процессивными РНКазами: hDIS3 и hDIS3L» . Журнал ЭМБО . 29 (14): 2342–57. дои : 10.1038/emboj.2010.121 . ПМЦ 2910271 . ПМИД 20531386 .
- ^ Миан, И.С. (1997). «Сравнительный анализ последовательностей рибонуклеаз HII, III, II PH и D» . Исследования нуклеиновых кислот . 25 (16): 3187–3195. дои : 10.1093/нар/25.16.3187 . ПМЦ 146874 . ПМИД 9241229 .
- ^ Райджмейкерс, Р; Шильдерс, Г; Пруейн, Дж.Дж. (2004). «Экзосома, молекулярная машина для контролируемой деградации РНК как в ядре, так и в цитоплазме». Европейский журнал клеточной биологии . 83 (5): 175–83. дои : 10.1078/0171-9335-00385 . ПМИД 15346807 .
- ^ Ван, Л; Льюис, MS; Джонсон, AW (2005). «Доменные взаимодействия внутри комплекса Ski2/3/8 и между комплексом Ski и Ski7p» . РНК . 11 (8): 1291–302. дои : 10.1261/rna.2060405 . ПМК 1370812 . ПМИД 16043509 .
- ^ ЛаКава, Дж; Хаусли, Дж; Савану, К; Петфальский, Э; Томпсон, Э; Жакье, А; Толлерви, Д. (2005). «Деградации РНК экзосомой способствует ядерный комплекс полиаденилирования» . Клетка . 121 (5): 713–24. дои : 10.1016/j.cell.2005.04.029 . ПМИД 15935758 . S2CID 14898055 .
- ^ Лю, Кью; Грейманн, Дж. К.; Лима, компакт-диск (2007). «Ошибка: восстановление, активность и структура экзосомы эукариотической РНК» . Клетка . 131 (1): 188–189. дои : 10.1016/j.cell.2007.09.019 .
- ^ Перейти обратно: а б Дзембовский, А; Лоренцен, Э; Конти, Э; Серафин, Б. (2007). «Одна субъединица, Dis3, по сути, отвечает за основную активность дрожжевой экзосомы». Структурная и молекулярная биология природы . 14 (1): 15–22. дои : 10.1038/nsmb1184 . ПМИД 17173052 . S2CID 24691764 .
- ^ Лю, Кью; Грейманн, Дж. К.; Лима, компакт-диск (2006). «Восстановление, активность и структура экзосомы эукариотической РНК» . Клетка . 127 (6): 1223–37. дои : 10.1016/j.cell.2006.10.037 . ПМИД 17174896 . S2CID 62785677 .
- ^ Лоренцен, Э; Конти, Э (2005). «Структурные основы распознавания 3'-конца РНК и экзорибонуклеолитического расщепления ядром экзосомы РНКазы PH» . Молекулярная клетка . 20 (3): 473–81. doi : 10.1016/j.molcel.2005.10.020 . ПМИД 16285928 .
- ^ ЛеЖен, Ф; Ли, Х; Макват, Л.Е. (2003). «Нонсенс-опосредованный распад мРНК в клетках млекопитающих включает декапирование, деаденилирование и экзонуклеолитическую активность» . Молекулярная клетка . 12 (3): 675–87. дои : 10.1016/S1097-2765(03)00349-6 . ПМИД 14527413 .
- ^ Уилсон, Массачусетс; Мо, С; Ван Хоф, А. (2007). «Геномный скрининг дрожжей раскрывает новые аспекты непрерывного метаболизма мРНК» . Генетика . 177 (2): 773–84. doi : 10.1534/genetics.107.073205 . ПМК 2034642 . ПМИД 17660569 .
- ^ Лин, В.Дж.; Даффи, А; Чен, CY (2007). «Локализация мРНК, содержащей элементы, богатые AU, в цитоплазматических гранулах, содержащих субъединицы экзосом» . Журнал биологической химии . 282 (27): 19958–68. дои : 10.1074/jbc.M702281200 . ПМИД 17470429 .
- ^ Перейти обратно: а б Оллманг, К; Куфель, Дж; Шанфро, Дж; Митчелл, П; Петфальский, Э; Толлерви, Д. (1999). «Функции экзосомы в синтезе рРНК, мяРНК и мяРНК» . Журнал ЭМБО . 18 (19): 5399–410. дои : 10.1093/emboj/18.19.5399 . ПМЦ 1171609 . ПМИД 10508172 .
- ^ Шильдерс, Г; Райджмейкерс, Р; Раатс, Дж. М.; Пруейн, Дж.Дж. (2005). «MPP6 представляет собой ассоциированный с экзосомами РНК-связывающий белок, участвующий в созревании 5,8S рРНК» . Исследования нуклеиновых кислот . 33 (21): 6795–804. дои : 10.1093/nar/gki982 . ПМЦ 1310903 . ПМИД 16396833 .
- ^ ван Дейк, Эль; Шильдерс, Г; Пруейн, Дж.Дж. (2007). «Для роста клеток человека необходима функциональная цитоплазматическая экзосома, которая участвует в различных путях распада мРНК» . РНК . 13 (7): 1027–35. дои : 10.1261/rna.575107 . ЧВК 1894934 . ПМИД 17545563 .
- ^ Карпусис А.Дж., Эй.Дж. (2002). «Деградосома РНК Escherichia coli: структура, функции и взаимоотношения в других рибонуклеолитических мультиферментных комплексах». Биохим. Соц. Транс . 30 (2): 150–5. дои : 10.1042/BST0300150 . ПМИД 12035760 .
- ^ Хаусли Дж., ЛаКава Дж., Толлерви Д. (июль 2006 г.). «Контроль качества РНК экзосомой». Нат. Преподобный мол. Клеточная Биол . 7 (7): 529–39. дои : 10.1038/nrm1964 . ПМИД 16829983 . S2CID 22499032 .
- ^ Уайерс Ф, Ф; Ружмейль, М; Бадис, Г; и др. (июнь 2005 г.). «Загадочные транскрипты pol II разрушаются путем ядерного контроля качества с участием новой поли(А)-полимеразы» . Клетка . 121 (5): 725–37. дои : 10.1016/j.cell.2005.04.030 . ПМИД 15935759 . S2CID 1390706 .
- ^ Нил Х., Малабат С., д'Обентон-Карафа Ю., Сюй З., Стейнмец Л.М., Жакье А. (февраль 2009 г.). «Широко распространенные двунаправленные промоторы являются основным источником загадочных транскриптов у дрожжей». Природа . 457 (7232): 1038–42. Бибкод : 2009Natur.457.1038N . дои : 10.1038/nature07747 . ПМИД 19169244 . S2CID 4329373 .
- ^ Прекер П, П; Нильсен, Дж; Каммлер, С; и др. (декабрь 2008 г.). «Истощение экзосом РНК обнаруживает транскрипцию выше активных человеческих промоторов» . Наука . 322 (5909): 1851–4. Бибкод : 2008Sci...322.1851P . дои : 10.1126/science.1164096 . ПМИД 19056938 .
- ^ Дж. Э. Поуп, Дж. Э. (2002). «Синдромы перекрытия склеродермии». Современное мнение в ревматологии . 14 (6): 704–10. дои : 10.1097/00002281-200211000-00013 . ПМИД 12410095 .
- ^ Гелпи, К; Альгеро, А; Анхелес Мартинес, М; Видаль, С; Хуарес, К; Родригес-Санчес, JL (1991). «Идентификация белковых компонентов, реагирующих с аутоантителами против PM/Scl» . Клиническая и экспериментальная иммунология . 81 (1): 59–64. дои : 10.1111/j.1365-2249.1990.tb05291.x . ПМК 1535032 . ПМИД 2199097 .
- ^ Таргофф, И.Н.; Райхлин, М. (1985). «Ядрышковая локализация антигена PM-Scl». Артрит и ревматизм . 28 (2): 226–30. дои : 10.1002/арт.1780280221 . ПМИД 3918546 .
- ^ Райджмейкерс, Р; Ренц, М; Виманн, К; Эгбертс, Западная Вирджиния; Силиг, HP; Ван Венрой, WJ; Пруейн, Дж.Дж. (2004). «PM-Scl-75 является основным аутоантигеном у пациентов с синдромом перекрытия полимиозита/склеродермии». Артрит и ревматизм . 50 (2): 565–9. дои : 10.1002/арт.20056 . ПМИД 14872500 .
- ^ Брауэр, Р; Ври Эгбертс, WT; Хенгстман, Дж.Дж.; Райджмейкерс, Р; Ван Энгелен, БГ; Силиг, HP; Ренц, М; Мирау, Р; и др. (2002). «Аутоантитела, направленные к новым компонентам комплекса PM/Scl, экзосомы человека» . Исследования и терапия артрита . 4 (2): 134–8. дои : 10.1186/ar389 . ПМЦ 83843 . ПМИД 11879549 .
- ^ Шильдерс, Г; Эгбертс, Западная Вирджиния; Райджмейкерс, Р; Пруейн, Дж.Дж. (2007). «C1D является основной мишенью аутоантител у пациентов с синдромом перекрытия полимиозита и склеродермии». Артрит и ревматизм . 56 (7): 2449–54. дои : 10.1002/арт.22710 . ПМИД 17599775 .
- ^ Малер, М; Райджмейкерс, Р; Дэнрих, К; Блютнер, М; Фрицлер, MJ (2005). «Клиническая оценка аутоантител к новому пептидному антигену PM/Scl» . Исследования и терапия артрита . 7 (3): R704–13. дои : 10.1186/ar1729 . ПМК 1174964 . ПМИД 15899056 .
- ^ Малер, М; Райджмейкерс, Р. (2007). «Новые аспекты аутоантител к комплексу PM/Scl: клинические, генетические и диагностические данные». Обзоры аутоиммунитета . 6 (7): 432–7. дои : 10.1016/j.autrev.2007.01.013 . ПМИД 17643929 .
- ^ Яблонска, С; Блащик, М (1998). «Склеромиозит: синдром перекрытия склеродермии и полимиозита». Клиническая ревматология . 17 (6): 465–7. дои : 10.1007/BF01451281 . ПМИД 9890673 . S2CID 39237322 .
- ^ Лам, ПЯ; Броня, компакт-диск; Степанянц, С.Б.; Кавет, Г; Вольф, МК; Батлер, Дж.С.; Хиншоу, Джей Си; Гарнье, П; и др. (2004). «Обнаружение механизмов действия терапевтических соединений с использованием полногеномного скрининга дрожжевых гетерозигот» . Клетка . 116 (1): 121–37. дои : 10.1016/S0092-8674(03)01035-3 . ПМИД 14718172 . S2CID 11922219 .
- ^ Ван, Дж.; Юршоу, М.; Мамса, Х.; Рудник-Шёнеборн, С.; Менезес, член парламента; Хонг, JE; Леонг, Д.В.; Сендерек, Дж.; Салман, MS; Читаят, Д.; Симан, П.; Фон Моерс, А.; Грауль-Нойманн, Л.; Корнберг, AJ; Кастро-Гаго, М.; Собридо, MAJS; Санефудзи, М.; Ши, ПБ; Саламон, Н.; Ким, RC; Винтерс, Х.В.; Чен, З.; Зеррес, К.; Райан, ММ; Нельсон, Сан-Франциско; Джен, Джей Си (2012). «Мутации в гене компонента экзосомы РНК EXOSC3 вызывают понтоцеребеллярную гипоплазию и дегенерацию мотонейронов спинного мозга» . Природная генетика . 44 (6): 704–708. дои : 10.1038/ng.2254 . ПМК 3366034 . ПМИД 22544365 .
Дальнейшее чтение
[ редактировать ]- Шильдерс, Г; Пруейн, Дж.Дж. (2008). «Глава 11 Биохимические исследования экзосомы млекопитающих с интактными клетками». Оборот РНК у эукариот: нуклеазы, пути и анализ распада мРНК . Методы энзимологии. Том. 448. стр. 211–226. дои : 10.1016/S0076-6879(08)02611-6 . ISBN 9780123743787 . ПМИД 19111178 .
- Хаусли, Дж; Толлерви, Д. (2008). «Механизм наблюдения за ядерными РНК: связь между нкРНК и структурой генома почкующихся дрожжей?». Биохим Биофиз Акта . 1779 (4): 239–246. дои : 10.1016/j.bbagrm.2007.12.008 . ПМИД 18211833 .
- Ванакова, С; Стефл, Р. (2007). «Контроль качества экзосом и РНК в ядре» . Отчеты ЭМБО . 8 (7): 651–657. дои : 10.1038/sj.embor.7401005 . ПМК 1905902 . ПМИД 17603538 .
- Бюттнер, К; Вениг, К; Хопфнер, КП (2006). «Экзосома: макромолекулярная клетка для контролируемой деградации РНК». Молекулярная микробиология . 61 (6): 1372–1379. CiteSeerX 10.1.1.232.6756 . дои : 10.1111/j.1365-2958.2006.05331.x . ПМИД 16968219 . S2CID 6872855 .
- Лоренцен, Э; Конти, Э (2006). «Экзосома и протеасома: наноотделения для деградации» . Клетка . 125 (4): 651–654. дои : 10.1016/j.cell.2006.05.002 . ПМИД 16713559 .
- Пруейн, Дж.Дж. (2005). «Пончики, имеющие дело с РНК». Структурная и молекулярная биология природы . 12 (7): 562–564. дои : 10.1038/nsmb0705-562 . ПМИД 15999107 . S2CID 43218090 .
Внешние ссылки
[ редактировать ]- Структура экзосомы человека в Банке данных белков RCSB
- Структура экзосомы архей в Банке белковых данных RCSB
- Структура архейной экзосомы, связанной с РНК, в Банке белковых данных RCSB
- Структура белка экзосомы дрожжей Rrp6 в Банке данных белков RCSB
- 3D макромолекулярные структуры экзосом в Банке данных EM (EMDB)