ФДЭ1
| Фосфодиэстераза I | |||
|---|---|---|---|
| Идентификаторы | |||
| Номер ЕС. | 3.1.4.1 | ||
| Номер CAS. | 9025-82-5 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| |||
Фосфодиэстераза 1 , ФДЭ1 , EC 3.1.4.1, систематическое название олигонуклеотид 5' - нуклеотидогидролаза ) — фосфодиэстераза, фермент также известный как кальций- и кальмодулин -зависимая фосфодиэстераза. Это одно из 11 семейств фосфодиэстеразы (ФДЭ1-ФДЭ11). Фосфодиэстераза 1 имеет три подтипа : PDE1A, PDE1B и PDE1C, которые далее делятся на различные изоформы . Различные изоформы обладают разным сродством к цАМФ и цГМФ . [1] [2]
Открытие
[ редактировать ]Существование Са 2+ -стимулированная фосфодиэстераза 1 была впервые продемонстрирована Cheung (1970), Kakiuchi и Yamazaki (1970) в результате их исследования бычьего мозга и мозга крысы соответственно. [1] [3] С тех пор было обнаружено, что он широко распространен в различных млекопитающих тканях , а также у других эукариот . В настоящее время это один из наиболее интенсивно изучаемых членов суперсемейства PDE . ферментов [3] который сегодня представляет 11 семейств генов , [1] [4] а также наиболее охарактеризованный. [3]
Дальнейшие исследования в этой области, наряду с увеличением доступности моноклональных антител, показали, что существуют различные изоферменты фосфодиэстеразы 1 , которые были идентифицированы и очищены. Сейчас известно, что фосфодиэстераза 1 существует в виде тканеспецифичных изоферментов. [3]
Структура
[ редактировать ]Семейство изоферментов фосфодиэстеразы 1 принадлежит к ферментам класса I. [2] [5] который включает все фосфодиэстеразы позвоночных и некоторые дрожжей . ферменты [5] Все ферменты класса I имеют каталитическое ядро, состоящее как минимум из 250 аминокислот , тогда как ферменты класса II лишены такой общей особенности. [5]
Обычно ФДЭ позвоночных представляют собой димеры линейных белков массой 50–150 кДа . [5] Они состоят из трех функциональных областей ; консервативное каталитическое ядро, регуляторный N-конец и С-конец [3-5]. Белки являются химерными , и каждый домен связан со своей конкретной функцией. [2]
Регуляторный N -конец существенно различается у разных типов фосфодиэстеразы. [4] [5] Они окружены каталитическим ядром и включают области, которые автоматически ингибируют каталитические домены. Они также нацелены на последовательности, которые контролируют субклеточную локализацию. В фосфодиэстеразы 1 эта область содержит домен, связывающий кальмодулин. [4]
Каталитические домены фосфодиэстеразы 1 (и других типов фосфодиэстеразы) имеют три спиральных субдомена: N-концевую циклиновую область, линкерную область и С-концевой спиральный пучок. глубокий гидрофобный На границе этих субдоменов образуется карман. Он состоит из четырех дочерних сайтов . Это: сайт связывания металла (М-сайт), основной карман (Q-карман), гидрофобный карман (Н-карман) и область крышки (L-область). М-сайт расположен на дне гидрофобного кармана с несколькими атомами металла . Атомы металлов связываются с остатками, которые полностью консервативны у всех членов семейства фосфодиэстераз. Идентичность атомов металла не известна с абсолютной уверенностью. Однако некоторые данные указывают на то, что по крайней мере один из металлов — цинк , а другой, скорее всего, магний . цинка Координационная сфера состоит из трех гистидинов , одного аспартата и двух молекул воды . В координационную сферу магния входит тот же аспартат, а также пять молекул воды, одна из которых является общей с молекулой цинка. Предполагаемая роль ионов металлов включает стабилизацию структуры, а также активацию гидроксид, обеспечивающий катализ . [4]
Домены разделены «шарнирными» областями, где их можно экспериментально разделить путем ограниченного протеолиза. [2]
Семейство изоферментов фосфодиэстеразы 1 (наряду с семейством фосфодиэстеразы 4) является наиболее разнообразным и включает многочисленные сплайсинговые варианты изоформ ФДЭ1. Он имеет три подтипа: PDE1A, PDE1B и PDE1C, которые далее делятся на различные изоформы. [1] [2]
Локализация
[ редактировать ]Локализация изоформ ФДЭ1 в различных тканях/клетках и их расположение внутри клеток следующая:
| изоформа | Тканевая/клеточная локализация | Внутриклеточная локализация |
|---|---|---|
| ПДЭ1А ( ПДЭ1А ) | Гладкая мускулатура, сердце, легкие, мозг, сперма. [2] | Преимущественно цитозольный [2] |
| PDE1A1 | Сердце, легкие [2] | Преимущественно цитозольный [2] |
| PDE1A2 | Мозг [2] | Преимущественно цитозольный [2] |
| ПДЭ1Б1 ( ПДЭ1Б ) | Нейроны, лимфоциты, гладкие мышцы [2] мозг, сердце, скелетные мышцы [1] | Цитозольный [2] |
| ПДЭ1Б2 | Макрофаги, лимфоциты [2] | Цитозольный [2] |
| ПДЭ1К ( ПДЭ1К ) | Мозг, пролиферирующие гладкие мышцы человека, сперматиды [2] | Цитозольный [2] |
| PDE1C1 | Мозг, сердце, яички [6] | - |
| PDE1C2 | Обонятельный эпителий [2] | Цитозольный [2] |
| PDE1C4/5 | мРНК присутствует в семенниках [6] | - |
Таблица 1. Различное расположение ФДЭ1 в тканях и внутри клеток.
Сообщается, что большинство изоформ PDE1 являются цитозольными . Однако есть случаи локализации PDE1 в субклеточных регионах, но мало что известно о молекулярных механизмах, ответственных за такую локализацию. Считается вероятным, что уникальные N-концевые или C-концевые области различных изоформ позволяют различным белкам нацеливаться на определенные субклеточные домены. [2]
Функциональная роль
[ редактировать ]Фосфодиэстераза1 катализирует следующую химическую реакцию : [7]
- Гидролитически удаляет 5' - нуклеотиды последовательно с 3' - гидроксиконцевых 3' - гидроксиконцевых олигонуклеотидов .
Он гидролизует как рибонуклеотиды , так и дезоксирибонуклеотиды , но обладает низкой активностью по отношению к полинуклеотидам .
Внутриклеточные вторичные мессенджеры, такие как цГМФ и цАМФ, претерпевают быстрые изменения концентрации в ответ на широкий спектр клеточно-специфичных стимулов. Концентрация этих вторичных мессенджеров во многом определяется относительной синтетической активностью циклазы и деградирующей активностью циклических нуклеотидов ФДЭ. [3] PDE1 разрушает как цГМФ, так и цАМФ. [8]
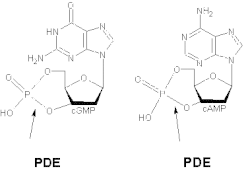
Различные изоформы обладают разным сродством к цАМФ и цГМФ. PDE1A и PDE1B преимущественно гидролизуют цГМФ, тогда как PDE1C разрушает как цАМФ, так и цГМФ с высоким сродством. Например, в гладких мышцах дыхательных путей человека и других видов на общий PDE1 приходится более 50% гидролитической активности циклических нуклеотидов. [9] Было продемонстрировано, что делеция и сверхэкспрессия PDE1 оказывает сильное влияние на индуцируемую агонистами передачу сигналов цАМФ, но мало влияет на базальный уровень цАМФ. [10] При корковых и таламических входах в полосатое тело активность PDE1 регулирует высвобождение нейротрансмиттера через цГМФ. [11]
Фармакология
[ редактировать ]Благодаря регуляции in vitro с помощью Ca 2+ Считается, что ФДЭ1 функционируют как механизм интеграции клеточных сигнальных путей, опосредованных цГМФ и цАМФ, с путями, которые регулируют внутриклеточные уровни кальция. [2] Точная функция изоферментов PDE1 в различных патофизиологических процессах не ясна, поскольку большинство исследований проводилось in vitro. Поэтому крайне важно направить дальнейшие исследования на исследования in vivo. [3]
PDE1 участвует в ряде физиологических и патологических процессов:
- PDE1A, скорее всего, служит для регулирования концентрации гладких мышц сосудов , и было обнаружено, что его уровень повышается в аорте крыс в ответ на хроническое лечение нитроглицерином . Также возможно, что он играет роль в функции сперматозоидов . [8]
- PDE1B У мышей с нокаутом повышена двигательная активность, а в некоторых парадигмах снижена память и способности к обучению. PDE1B также участвует в дофаминергической передаче сигналов и индуцируется в нескольких типах активированных иммунных клеток. [8] PDE1B мРНК индуцируется в PHA или анти-CD3/CD28-активированных Т-лимфоцитах человека и участвует в регуляции IL-13, вовлеченной в аллергические заболевания. [1]
- Было показано, что PDE1C является основным регулятором пролиферации гладких мышц, по крайней мере, в гладких мышцах человека. Непролиферирующие гладкомышечные клетки (ГМК) демонстрируют лишь низкий уровень экспрессии PDE1C , но он высоко экспрессируется в пролиферирующих ГМК. Таким образом, можно предположить, что ингибирование PDE1C может иметь положительные эффекты благодаря предполагаемому ингибированию пролиферации ГМК, событию, которое вносит важный вклад в патофизиологию атеросклероза . [8] Другая вероятная роль PDE1C заключается в обонянии [4], регуляции функции сперматозоидов и передаче сигналов нейронов . [8]
Регулирование
[ редактировать ]Отличительной особенностью PDE1 как семейства является их регуляция кальцием (Ca 2+ ) и кальмодулин (CaM). [12] Было показано, что кальмодулин активирует циклические нуклеотиды ФДЭ кальций-зависимым образом и кооперативное связывание четырех Са 2+ кальмодулин необходим для полной активации ФДЭ1 [2]. Связывание одного Ca 2+ Комплекс /CaM на мономер с сайтами связывания вблизи N-конца стимулирует гидролиз циклических нуклеотидов. В интактных клетках PDE1 почти исключительно активируется Ca. 2+ поступление в клетку из внеклеточного пространства. Регуляция ФДЭ1 с помощью Ca 2+ и CaM были изучены in vitro, и эти исследования показали, что восемь остатков метионина в гидрофобных щелях Ca 2+ -CaM необходимы для связывания и активации PDE1. Мутации в N-концевой доле CaM влияют на его способность активировать PDE1, поэтому считается, что C-концевая доля CaM служит для нацеливания CaM на PDE1, в то время как N-концевая доля активирует фермент. Наличие ароматического остатка, обычно триптофана , в CaM-связывающей области Ca. 2+ -CaM-регулируемые белки также могут потребоваться для связывания с PDE1. [12]
Между разными изоферментами PDE1 существует значительная разница в сродстве к Ca. 2+ /КаМ. В целом ферменты PDE1 обладают высоким сродством к комплексу, но на сродство может влиять фосфорилирование. Фосфорилирование PDE1A1 и PDE1A2 протеинкиназой A и PDE1B1 CaM-киназой II снижает их чувствительность к активации кальмодулина. [1] Это фосфорилирование может быть обращено вспять фосфатазой кальциневрином. [2] Фосфорилирование изоферментов сопровождается снижением сродства изоферментов к СаМ, а также увеличением содержания Са 2+ концентрации, необходимые для активации CaM изоферментов. [3]
Ингибиторы и их функции
[ редактировать ]ФДЭ использовались в качестве терапевтических целей из-за основного фармакологического принципа, согласно которому регуляция деградации любого лиганда или вторичного мессенджера часто может вызывать более быстрое и большее процентное изменение концентрации, чем сопоставимые скорости синтеза . Другая причина заключается в том, что ФДЭ не должны конкурировать с очень высокими уровнями эндогенного субстрата, чтобы быть эффективными, поскольку уровни цАМФ и цГМФ в большинстве клеток обычно находятся в микромолярном диапазоне. [2]
Наличие кристаллических структур каталитических доменов ФДЭ с высоким разрешением делает возможной разработку высокоэффективных и специфических ингибиторов . [6]
Многие соединения, о которых сообщалось как ингибиторы ФДЭ1, не взаимодействуют напрямую с каталитическим сайтом ФДЭ1, но взаимодействуют во время активации либо на уровне сайтов связывания кальмодулина, таких как соединение KS505a , либо непосредственно на Ca. 2+ /кальмодулин, такой как беприл , флунаризин и амиодарон . [1]
Те ингибиторы, которые взаимодействуют с каталитическим центром, занимают часть активного центра, главным образом вокруг Q-кармана и иногда вблизи М-кармана. [13] Основной точкой взаимодействия является консервативный гидрофобный карман, который участвует в ориентации пуринового кольца субстрата для взаимодействия с остатком глутамина, который имеет решающее значение для каталитического механизма ФДЭ. [6]
Взаимодействия ингибиторов можно разделить на три основных типа: взаимодействия с ионами металлов, опосредованные водой, взаимодействия Н-связей с белковыми остатками, участвующими в узнавании нуклеотидов, и, что наиболее важно, взаимодействие с гидрофобными остатками, выстилающими полость активного центра. Все известные ингибиторы, похоже, используют эти три типа взаимодействий, и, следовательно, эти взаимодействия должны служить основой для разработки новых типов ингибиторов. [13]
Первоначально ингибиторы ФДЭ1 считались эффективными сосудистыми релаксантами. Однако благодаря доступности очищенных клонированных ферментов теперь известно, что такие ингибиторы на самом деле одинаково активны против ФДЭ5. [4] Эти ингибиторы включают, например, запринаст , 8-метоксиметил IPMX и SCH 51866 . [1]
Все терапевтически эффективные ингибиторы ФДЭ должны быть включены в клетку, поскольку все ФДЭ локализованы в цитоплазме и/или на внутриклеточных мембранах. [8]
На сегодняшний день не существует реального и эффективного специфического ингибитора ФДЭ1, который можно было бы использовать для оценки функциональной роли ФДЭ1 в тканях. [1]
Общие ингибиторы
[ редактировать ]

Нимодипин представляет собой дигидропиридин , который специфически блокирует кальций L-типа. 2+ -канала и впервые был описан как ингибитор PDE1. Этот эффект не связан с его свойствами антагониста кальция, поскольку он ингибирует в микромолярном диапазоне базальную и стимулированную кальмодулином очищенную ФДЭ1. Поскольку нимодипин в более низких концентрациях блокирует кальциевые каналы L-типа, его можно использовать только для оценки участия ФДЭ1 в гомогенатах тканей и клеток. [1]
Винпоцетин был описан как специфический ингибитор базальной и кальмодулин-активируемой ФДЭ1. Этот эффект приводит к увеличению содержания цАМФ по сравнению с цГМФ. [1] [14] В основном он используется в качестве фармакологического инструмента для выявления PDE1. Винпоцетин по-разному ингибирует различные подтипы ФДЭ1 (IC 50 от 8 до 50 мкм), а также способен ингибировать ФДЭ7В. Его нельзя использовать в качестве специального инструмента для исследования функциональной роли ФДЭ1 из-за его прямого активаторного воздействия на каналы BK(Ca). [1] Винпоцетин проникает через гематоэнцефалический барьер и поглощается тканями головного мозга. Была выдвинута гипотеза, что винпоцетин может влиять на потенциалзависимые кальциевые каналы. [14]
IC224 ингибирует ФДЭ1 (IC 50 = 0,08 мкМ) с селективным коэффициентом 127 (соотношение значения IC 50 для следующего наиболее чувствительного ФДЭ и значения IC 50 для ФДЭ1). Его разработала корпорация ICOS. Если IC224 аналогичным образом ингибирует базальный и кальмодулин-активируемый подтипы ФДЭ1, это соединение может быть очень полезным для характеристики активности ФДЭ1 и четкого изучения различных ролей ФДЭ1 в патофизиологии. [1]
Ингибиторы при заболеваниях
[ редактировать ]Почти все фосфодиэстеразы экспрессируются в ЦНС, что делает это семейство генов привлекательным источником новых мишеней для лечения психиатрических и нейродегенеративных расстройств. [6]
PDE1A2 играет потенциальную роль в нейродегенеративных заболеваниях, в том числе: [4]
- болезнь Паркинсона
- аксональных нейрофиламентов Деградация
- Деградация мотонейронов
- Нейрональная ишемия
- болезнь Альцгеймера
- Эпилепсия
PDE1C может играть роль в регуляции инсулина высвобождения [5] и может воздействовать на пролиферирующие гладкомышечные клетки при атеросклеротических поражениях или во время рестеноза . [4] [15]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н Люнье С. (март 2006 г.). «Суперсемейство циклических нуклеотидных фосфодиэстераз (ФДЭ): новая мишень для разработки специфических терапевтических средств». Фармакол. Там . 109 (3): 366–98. doi : 10.1016/j.pharmthera.2005.07.003 . ПМИД 16102838 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В Бендер А.Т., Беаво Дж.А. (сентябрь 2006 г.). «Циклические нуклеотидные фосфодиэстеразы: молекулярная регуляция для клинического использования». Фармакол. Преподобный . 58 (3): 488–520. дои : 10.1124/пр.58.3.5 . ПМИД 16968949 . S2CID 7397281 .
- ^ Перейти обратно: а б с д и ж г Каккар Р., Раджу Р.В., Шарма Р.К. (июль 1999 г.). «Кальмодулинзависимая циклическая нуклеотидфосфодиэстераза (ФДЭ1)» . Клетка. Мол. Наука о жизни . 55 (8–9): 1164–86. дои : 10.1007/s000180050364 . ПМЦ 11146777 . ПМИД 10442095 . S2CID 23724340 . [ постоянная мертвая ссылка ]
- ^ Перейти обратно: а б с д и ж г Чон Й.Х., Хо Й.С., Ким СМ и др. (июнь 2005 г.). «Фосфодиэстераза: обзор белковых структур, потенциальное терапевтическое применение и недавний прогресс в разработке лекарств» . Клетка. Мол. Наука о жизни . 62 (11): 1198–220. дои : 10.1007/s00018-005-4533-5 . ПМЦ 11139162 . ПМИД 15798894 . S2CID 9806864 .
- ^ Перейти обратно: а б с д и ж Доуса Т.П. (январь 1999 г.). «Изозимы циклической-3 ' ,5' - нуклеотидной фосфодиэстеразы в клеточной биологии и патофизиологии почек» . Почки Int . 55 (1): 29–62. дои : 10.1046/j.1523-1755.1999.00233.x . ПМИД 9893113 .
- ^ Перейти обратно: а б с д и Меннити Ф.С., Фарачи В.С., Шмидт К.Дж. (август 2006 г.). «Фосфодиэстеразы в ЦНС: цели для разработки лекарств». Nat Rev Drug Discov . 5 (8): 660–70. дои : 10.1038/nrd2058 . ПМИД 16883304 . S2CID 419810 .
- ^ Хорана Г.Х. (1961). «Фосфодиэстераза». В Бойер П.Д., Ларди Х., Мирбек К. (ред.). Ферменты . Том. 5 (2-е изд.). Нью-Йорк: Академическая пресса. стр. 79–94.
- ^ Перейти обратно: а б с д и ж Бишофф Э. (июнь 2004 г.). «Потенциал, селективность и последствия неселективности ингибирования ФДЭ» . Межд. Дж. Импот. Рез . 16 (Приложение 1): С11–4. дои : 10.1038/sj.ijir.3901208 . ПМИД 15224129 .
- ^ Гимбич М.А. (июнь 2005 г.). «Жизнь после ФДЭ4: преодоление побочных эффектов с помощью ингибиторов фосфодиэстеразы двойной специфичности». Карр Опин Фармакол . 5 (3): 238–44. дои : 10.1016/j.coph.2005.04.001 . ПМИД 15907909 .
- ^ Тевелейн Дж. М., де Винде Дж. Х. (сентябрь 1999 г.). «Новые механизмы чувствительности и мишени для пути цАМФ-протеинкиназы А в дрожжах Saccharomyces cerevisiae » . Мол. Микробиол . 33 (5): 904–18. дои : 10.1046/j.1365-2958.1999.01538.x . ПМИД 10476026 .
- ^ Фиблингер, Тим; Перес-Альварес, Альберто; Ламот-Молина, Пол Дж; Ну и дела, Кристин Э; Эртнер, Томас Дж. (3 августа 2022 г.). «Пресинаптический цГМФ устанавливает синаптическую силу в полосатом теле и важен для двигательного обучения» . Отчеты ЭМБО . 23 (8): e54361. дои : 10.15252/эмбр.202154361 . ISSN 1469-221X . ПМЦ 9346481 . ПМИД 35735260 .
- ^ Перейти обратно: а б Горая Т.А., Купер Д.М. (июль 2005 г.). "Ка 2+ -кальмодулинзависимая фосфодиэстераза (PDE1): текущие перспективы». Cell. Signal . 17 (7): 789–97. doi : 10.1016/j.cellsig.2004.12.017 . PMID 15763421 .
- ^ Перейти обратно: а б Кард Г.Л., Англия BP, Suzuki Y и др. (декабрь 2004 г.). «Структурные основы активности препаратов, ингибирующих фосфодиэстеразу» . Структура . 12 (12): 2233–47. дои : 10.1016/j.str.2004.10.004 . ПМИД 15576036 .
- ^ Перейти обратно: а б «Винпоцетин. Монография». Альтернативный Мед Преп . 7 (3): 240–3. Июнь 2002 г. PMID 12126465 .
- ^ Мацумото Т., Кобаяши Т., Камата К. (август 2003 г.). «Фосфодиэстеразы в сосудистой системе» . J Резус гладких мышц . 39 (4): 67–86. дои : 10.1540/jsmr.39.67 . ПМИД 14692693 .
Внешние ссылки
[ редактировать ]- Фосфодиэстераза + I в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)