Репликация эукариотической ДНК

Репликация эукариотической ДНК — это консервативный механизм, который ограничивает репликацию ДНК один раз за клеточный цикл. Репликация хромосомной ДНК эукариот играет центральную роль в дупликации клетки и необходима для поддержания эукариотического генома .
Репликация ДНК — это действие ДНК-полимераз , синтезирующих цепь ДНК, комплементарную исходной цепи матрицы. Для синтеза ДНК двухцепочечная ДНК раскручивается ДНК- хеликазами перед полимеразами, образуя репликационную вилку, содержащую две одноцепочечные матрицы. Процессы репликации позволяют копировать одну двойную спираль ДНК в две спирали ДНК, которые делятся на дочерние клетки при митозе . Основные ферментативные функции, выполняемые в репликационной вилке, хорошо консервативны от прокариот к эукариотам , но механизм репликации при репликации эукариотической ДНК представляет собой гораздо более крупный комплекс, координирующий многие белки в месте репликации, образуя реплисому . [1]
Реплисома отвечает за копирование всей геномной ДНК в каждой пролиферативной клетке. Этот процесс обеспечивает высокоточную передачу наследственной/генетической информации от родительской клетки к дочерней и, таким образом, важен для всех организмов. Большая часть клеточного цикла построена на обеспечении безошибочной репликации ДНК. [1]
В G1 фазе . клеточного цикла инициируются многие процессы регуляции репликации ДНК У эукариот большая часть синтеза ДНК происходит во время S-фазы клеточного цикла, и весь геном должен быть раскручен и продублирован для образования двух дочерних копий. Во время G 2 исправляются любые повреждения ДНК или ошибки репликации. Наконец, одна копия генома выделяется в каждую дочернюю клетку в митозе или М-фазе. [2] Каждая из этих дочерних копий содержит одну цепь родительского дуплекса ДНК и одну возникающую антипараллельную цепь.
Этот механизм сохраняется от прокариот к эукариотам и известен как полуконсервативная репликация ДНК. Процесс полуконсервативной репликации сайта репликации ДНК представляет собой вилкообразную структуру ДНК, репликационную вилку, в которой спираль ДНК открыта или раскручивается, открывая неспаренные нуклеотиды ДНК для распознавания и спаривания оснований для включения.свободных нуклеотидов в двухцепочечную ДНК. [3]
Инициация
[ редактировать ]Инициация репликации эукариотической ДНК - это первая стадия синтеза ДНК, на которой двойная спираль ДНК раскручивается, и на ведущей цепи происходит начальное событие прайминга ДНК-полимеразы α. Событие прайминга на отстающей цепи устанавливает репликационную вилку. Праймирование спирали ДНК состоит из синтеза праймера РНК, обеспечивающего синтез ДНК ДНК-полимеразой α. Праймирование происходит один раз в начале координат ведущей цепи и в начале каждого фрагмента Окадзаки на отстающей цепи.
Происхождение репликации
[ редактировать ]Репликация начинается с истоков репликации . Последовательности ДНК, содержащие эти сайты, были первоначально выделены в конце 1970-х годов на основании их способности поддерживать репликацию плазмид , отсюда и название автономно реплицирующихся последовательностей (ARS). Источники сильно различаются по своей эффективности: некоторые из них используются почти в каждом клеточном цикле, тогда как другие могут использоваться только в одной из тысячи S-фаз. [4] Общее количество дрожжевых ARS составляет не менее 1600, но может быть и более 5000, если учитывать меньшее количество активных сайтов. [5] то есть ARS может встречаться каждые 2000–8000 пар оснований.
Пререпликативный комплекс
[ редактировать ]Множественные репликативные белки собираются и отделяются от этих источников репликации, чтобы инициировать репликацию ДНК. [6] при этом образование пререпликационного комплекса (пре-РК) является ключевым промежуточным звеном в процессе инициации репликации.
Ассоциация комплекса распознавания происхождения (ORC) с началом репликации рекрутирует белок 6 цикла деления клетки (Cdc6) для формирования платформы для загрузки белков комплекса поддержания минихромосомы (Mcm 2–7) , чему способствует лицензирование хроматина и ДНК. Белок фактора репликации 1 (Cdt1). ORC, Cdc6 и Cdt1 вместе необходимы для стабильной ассоциации комплекса Mcm2-7 с репликативными источниками во время фазы G1 клеточного цикла. [7]
Эукариотические начала репликации контролируют образование нескольких белковых комплексов, которые приводят к сборке двух двунаправленных вилок репликации ДНК. Эти события инициируются образованием пререпликационного комплекса (пре-РК) в истоках репликации. Этот процесс происходит на стадии G1 клеточного цикла. Формирование пре-RC включает упорядоченную сборку многих факторов репликации, включая комплекс распознавания происхождения (ORC), белок Cdc6, белок Cdt1 и белки поддержания минихромосом (Mcm2-7). [8] [9] После образования пре-РК активация комплекса запускается двумя киназами , циклин-зависимой киназой 2 (CDK) и Dbf4-зависимой киназой (DDK), которые помогают перевести пре-РК в инициирующий комплекс до инициации ДНК. репликация. Этот переход включает в себя упорядоченную сборку дополнительных факторов репликации для раскручивания ДНК и накопления множества эукариотических ДНК-полимераз вокруг раскрученной ДНК. Центральным вопросом о том, как двунаправленные репликационные вилки устанавливаются в точках начала репликации, является механизм, с помощью которого ORC рекрутирует два противоположных комплекса Mcm2-7 в каждую точку начала репликации, чтобы сформировать пререпликационный комплекс. [10] [11] [12]
Комплекс распознавания происхождения
[ редактировать ]Первым шагом в сборке пререпликационного комплекса (пре-РК) является связывание комплекса распознавания ориджина (ORC) с ориджином репликации. В позднем митозе белок Cdc6 присоединяется к связанному ORC с последующим связыванием комплекса Cdt1-Mcm2-7. [13] ORC, Cdc6 и Cdt1 необходимы для загрузки комплекса обслуживания минихромосом из шести белков (Mcm 2–7) в ДНК. ORC представляет собой белковый комплекс Orc1p-6, состоящий из шести субъединиц, который выбирает сайты начала репликации на ДНК для инициации репликации, а связывание ORC с хроматином регулируется в ходе клеточного цикла. [8] [14] Как правило, функция и размер субъединиц ORC консервативны во многих геномах эукариот, с той разницей, что у них различаются сайты связывания ДНК.
Наиболее широко изученным комплексом распознавания происхождения является комплекс Saccharomyces cerevisiae или дрожжей, который, как известно, связывается с автономно реплицирующейся последовательностью (ARS). [15] ORC S. cerevisiae специфически взаимодействует с элементами A и B1 точки начала репликации дрожжей, охватывая область из 30 пар оснований . [16] Для связывания с этими последовательностями требуется АТФ . [8] [16]
атомная структура ORC S. cerevisiae , связанного с ДНК ARS. Определена [16] Orc1, Orc2, Orc3, Orc4 и Orc5 окружают элемент A посредством двух типов взаимодействий: неспецифических по основаниям и специфичных по основаниям, которые изгибают ДНК у элемента A. Все пять субъединиц контактируют с сахарофосфатным остовом в нескольких точках элемента А, образуя прочное сцепление без специфичности основания. Orc1 и Orc2 контактируют с малой бороздкой элемента A, тогда как домен крылатой спирали Orc4 контактирует с метильными группами инвариантных Ts в большой бороздке элемента A через вставочную спираль (IH). Отсутствие этого ИГ у многоклеточных животных. [16] объясняет отсутствие специфичности последовательности в ORC человека. [17] Удаление IH из Sc ORC приводит к тому, что он теряет свою специфичность к элементу A и беспорядочно и преимущественно (83%) связывается с областями промотора. [18] ДНК ARS также изгибается у элемента B1 посредством взаимодействия с Orc2, Orc5 и Orc6. [16] Изгиб исходной ДНК с помощью ORC, по-видимому, эволюционно консервативен, что позволяет предположить, что оно может быть необходимо для сложного механизма загрузки Mcm2-7. [16] [19]
Когда ORC связывается с ДНК в точках начала репликации, он служит основой для сборки других ключевых факторов инициации пререпликативного комплекса. [20] Эта пререпликативная сборка комплекса на стадии G1 клеточного цикла необходима до активации репликации ДНК во время S-фазы. [21] Удаление по крайней мере части комплекса (Orc1) из хромосомы в метафазе является частью регуляции ORC млекопитающих, обеспечивающей устранение образования пререпликативного комплекса до завершения метафазы. [22]
белок Cdc6
[ редактировать ]Связывание белка цикла 6 клеточного деления (Cdc6) с комплексом распознавания начала (ORC) является важным шагом в сборке пререпликационного комплекса (pre-RC) в истоках репликации. Cdc6 связывается с ORC на ДНК АТФ-зависимым образом, что вызывает изменение в паттерне связывания источника, требующее АТФазы Orc1 . [23] Cdc6 требует ORC для связи с хроматином и, в свою очередь, необходим для гептамера Cdt1-Mcm2-7. [13] связываться с хроматином. [24] Комплекс ORC-Cdc6 образует кольцевую структуру и аналогичен другим АТФ-зависимым белковым машинам. Уровни и активность Cdc6 регулируют частоту, с которой точки начала репликации используются во время клеточного цикла.
белок Cdt1
[ редактировать ]Белок лицензирования хроматина и фактора репликации ДНК 1 (Cdt1) необходим для лицензирования хроматина для репликации ДНК. [25] [26] У S. cerevisiae Cdt1 облегчает загрузку комплекса Mcm2-7 по одному на хромосому путем стабилизации левосторонней структуры открытого кольца одиночного гексамера Mcm2-7. [13] [27] [28] Было показано, что Cdt1 связывается с C-концом Cdc6, совместно способствуя ассоциации белков Mcm с хроматином. [29] Крио-ЭМ структура комплекса OCCM (ORC-Cdc6-Cdt1-MCM) показывает, что Cdt1-CTD взаимодействует с Mcm6-WHD. [30] У многоклеточных животных активность Cdt1 во время клеточного цикла жестко регулируется за счет его ассоциации с белком геминином , который одновременно ингибирует активность Cdt1 во время S-фазы, чтобы предотвратить повторную репликацию ДНК, и предотвращает ее убиквитинирование и последующий протеолиз . [31]
Белковый комплекс поддержания минихромосомы
[ редактировать ]Белки поддержания минихромосомы (Mcm) были названы в честь генетического скрининга мутантов инициации репликации ДНК у S. cerevisiae , которые влияют на стабильность плазмиды специфичным для ARS образом . [32] Mcm2, Mcm3, Mcm4, Mcm5, Mcm6 и Mcm7 образуют гексамерный комплекс, имеющий структуру открытого кольца с зазором между Mcm2 и Mcm5. [13] Сборка белков Mcm на хроматине требует скоординированной функции комплекса распознавания происхождения (ORC), Cdc6 и Cdt1. [33] Как только белки Mcm загружены на хроматин, ORC и Cdc6 могут быть удалены из хроматина, не препятствуя последующей репликации ДНК. Это наблюдение предполагает, что основная роль пререпликационного комплекса заключается в правильной загрузке белков Mcm. [34]

Белки Mcm на хроматине образуют двойной гексамер «голова к голове», где два кольца слегка наклонены, скручены и смещены от центра, создавая излом в центральном канале, где связанная ДНК захватывается на границе двух колец. [35] [36] Каждое гексамерное кольцо Mcm2-7 сначала служит каркасом для сборки реплисомы, а затем ядром каталитической геликазы CMG (Cdc45-MCM-GINS), которая является основным компонентом реплисомы. Каждый белок Mcm во многом связан со всеми остальными, но уникальные последовательности, отличающие каждый из них, [18] типы субъединиц консервативны у эукариот. Все эукариоты имеют ровно шесть аналогов белка Mcm, каждый из которых относится к одному из существующих классов (Mcm2–7), что указывает на то, что каждый белок Mcm выполняет уникальную и важную функцию. [37] [11]
Белки поддержания минихромосом необходимы для активности ДНК-хеликазы. Инактивация любого из шести белков Mcm во время S-фазы необратимо предотвращает дальнейшее развитие репликационной вилки, что позволяет предположить, что геликаза не может быть переработана и должна быть собрана в точках начала репликации. [38] Наряду с хеликазной активностью комплекса поддерживающих белков минихромосом, комплекс также обладает связанной с ним АТФазной активностью. [39] Исследования показали, что внутри белкового комплекса Mcm имеются специфические каталитические пары белков Mcm, которые функционируют вместе, координируя гидролиз АТФ. [40] Эти исследования, подтвержденные крио-ЭМ структурами комплексов Mcm2-7, [13] [35] показали, что комплекс Mcm представляет собой гексамер, субъединицы которого расположены в кольце в порядке Mcm2-Mcm6-Mcm4-Mcm7-Mcm3-Mcm5-. Оба члена каждой каталитической пары вносят вклад в конформацию, которая обеспечивает связывание и гидролиз АТФ, а смесь активных и неактивных субъединиц предположительно позволяет гексамерному комплексу Mcm завершить связывание АТФ и гидролиз в целом для создания скоординированной активности АТФазы. [41]
Ядерная локализация поддерживающих белков минихромосом регулируется в почкующихся дрожжевых клетках. [42] [43] Белки Mcm присутствуют в ядре на стадии G1 и S-фазе клеточного цикла, но экспортируются в цитоплазму на стадии G2 и М-фазе. Полный и неповрежденный комплекс Mcm из шести субъединиц необходим для проникновения в ядро клетки. [44] У S. cerevisiae экспорту ядер способствует активность циклин-зависимой киназы (CDK). Белки Mcm, связанные с хроматином, защищены от механизма экспорта CDK из-за отсутствия доступа к CDK. [45]
Комплекс инициации
[ редактировать ]На стадии G1 клеточного цикла факторы инициации репликации, комплекс распознавания происхождения (ORC), Cdc6, Cdt1 и белковый комплекс поддержания минихромосомы (Mcm) последовательно связываются с ДНК, образуя прямой димер Кольцевой комплекс MCM, известный как пререпликационный комплекс (пре-РК). Хотя пре-РК дрожжей образует закрытый комплекс ДНК, [35] [36] [46] человеческий преРК образует открытый комплекс. [47] При переходе стадии G1 к S-фазе клеточного цикла специфичные для S-фазы циклинзависимая протеинкиназа (CDK) и киназа Cdc7/Dbf4 (DDK) преобразуют инертный пре-РК в активный комплекс, способный к сборке две двунаправленные реплисомы. КриоЭМ структуры [48] [49] [50] показали, что два DDK независимо стыкуются с интерфейсом двойного гексамера MCM, расположенного между двумя кольцами. Последовательное фосфорилирование множественных субстратов на NTE Mcm4, Mcm2 и Mcm6 достигается с помощью механизма качания, посредством которого Dbf4 принимает различные состояния качания, чтобы расположить Cdc7 над своими множественными субстратами. [50] Фосфорилирование двойного гексамера MCM, в частности Mcm4-NSD, с помощью DDK имеет важное значение для жизнеспособности дрожжей. [51] Рекрутирование Cdc45 и GINS следует после активации MCM с помощью DDK и CDK.
белок Cdc45
[ редактировать ]Белок 45-го цикла клеточного деления (Cdc45) является критическим компонентом преобразования пререпликативного комплекса в инициирующий комплекс. Белок Cdc45 собирается в точках начала репликации до инициации и необходим для начала репликации у Saccharomyces cerevisiae , а также играет важную роль во время элонгации. Т.о., Cdc45 играет центральную роль как в фазе инициации, так и в фазе элонгации репликации хромосомной ДНК. [52]
Cdc45 связывается с хроматином после начала инициации на поздней стадии G1 и во время S-фазы клеточного цикла. Cdc45 физически связан с Mcm5 и демонстрирует генетические взаимодействия с пятью из шести членов семейства генов Mcm и геном ORC2 . [53] [54] Загрузка Cdc45 на хроматин имеет решающее значение для загрузки других различных белков репликации, включая ДНК-полимеразу α , ДНК-полимеразу ε, репликационный белок A (RPA) и ядерный антиген пролиферирующих клеток (PCNA) на хроматин. [55] [56] [57] [58] В безъядерной системе Xenopus было продемонстрировано, что Cdc45 необходим для раскручивания плазмидной ДНК. [58] Безъядерная система Xenopus также демонстрирует, что раскручивание ДНК и прочное связывание RPA с хроматином происходит только в присутствии Cdc45. [55]
Связывание Cdc45 с хроматином зависит от активности киназы Clb-Cdc28, а также от функциональных Cdc6 и Mcm2, что позволяет предположить, что Cdc45 связывается с пре-RC после активации S-фазных циклин-зависимых киназ (CDK). Как показывают сроки и зависимость от CDK, связывание Cdc45 с хроматином имеет решающее значение для инициации репликации ДНК. Во время S-фазы Cdc45 физически взаимодействует с белками Mcm на хроматине; однако диссоциация Cdc45 от хроматина происходит медленнее, чем у Mcm, что указывает на то, что белки высвобождаются по разным механизмам. [37]
ДЖИНЫ
[ редактировать ]Шесть поддерживающих минихромосомных белков и Cdc45 необходимы во время инициации и элонгации для движения репликационных вилок и для раскручивания ДНК. GINS необходимы для взаимодействия Mcm и Cdc45 в истоках репликации во время инициации, а затем в вилках репликации ДНК по мере развития реплисомы. [59] [60] Комплекс GINS состоит из четырех небольших белков Sld5 (Cdc105), Psf1 (Cdc101), Psf2 (Cdc102) и Psf3 (Cdc103). GINS представляет собой «go, ichi, ni, san», что означает «5, 1, 2, 3». ' по-японски. [61] Cdc45, Mcm2-7 и GINS вместе образуют геликазу CMG, [62] репликативная геликаза реплисомы. Хотя сам по себе комплекс Mcm2-7 обладает слабой геликазной активностью. [63] Cdc45 и GINS необходимы для устойчивой активности геликазы. [64] [65]
Мкм10
[ редактировать ]Mcm10 необходим для репликации хромосом и взаимодействует с хеликазой обслуживания минихромосом 2-7, которая загружена в неактивной форме в инициации репликации ДНК. [66] [67] Mcm10 также сопровождает каталитическую ДНК-полимеразу α и помогает стабилизировать полимеразу на вилках репликации. [68] [69]
Киназы DDK и CDK
[ редактировать ]В начале S-фазы пререпликативный комплекс должен быть активирован двумя специфичными для S-фазы киназами, чтобы сформировать инициирующий комплекс в начале репликации. Одна киназа — это киназа Cdc7-Dbf4, называемая Dbf4-зависимой киназой (DDK), а другая — циклин-зависимая киназа (CDK). [70] Анализы связывания Cdc45 с хроматином у дрожжей и Xenopus показали, что последующим событием действия CDK является загрузка Cdc45 на хроматин. [71] [72] Cdc6 Предполагается, что является мишенью действия CDK из-за связи между Cdc6 и CDK, а также CDK-зависимого фосфорилирования Cdc6. Считалось, что CDK-зависимое фосфорилирование Cdc6 необходимо для перехода в S-фазу. [73]
Как каталитические субъединицы DDK, Cdc7, так и белок-активатор Dbf4 консервативны у эукариот и необходимы для начала S-фазы клеточного цикла. [74] [75] И Dbf4, и Cdc7 необходимы для загрузки Cdc45 в точки начала репликации хроматина. Мишенью связывания киназы DDK является связанная с хроматином форма комплекса Mcm. [76] [77] КриоЭМ-структуры высокого разрешения показали, что субъединица Dbf4 DDK проходит через гексамерный интерфейс связанного с ДНК MCM-DH, контактируя с Mcm2 одного гексамера и Mcm4/6 противоположного гексамера. [48] [49] [50] Mcm2, Mcm4 и Mcm6 являются субстратами фосфорилирования DDK. [78] [74] но только N-концевой богатый серином/треонином домен (NSD) Mcm4 является важной мишенью DDK. [50] [51] Фосфорилирование NSD приводит к активации активности Mcm-хеликазы.
Белки Dpb11, Sld3 и Sld2
[ редактировать ]Sld3, Sld2 и Dpb11 взаимодействуют со многими белками репликации. Sld3 и Cdc45 образуют комплекс, который ассоциирован с pre-RC в ранних инициаторах репликации даже в фазе G11 и с более поздними инициаторами репликации в S-фазе взаимно Mcm-зависимым образом. [79] [80] Dpb11 и Sld2 взаимодействуют с полимеразой ɛ, и эксперименты по сшиванию показали, что Dpb11 и полимераза ɛ соосаждаются в S-фазе и связываются с точками начала репликации. [81] [82]
Sld3 и Sld2 фосфорилируются CDK, что позволяет двум репликативным белкам связываться с Dpb11. Dpb11 имел две пары C-концевых доменов BRCA1 (BRCT), которые известны как фосфопептидсвязывающие домены. [83] N-концевая пара доменов BRCT связывается с фосфорилированным Sld3, а C-концевая пара связывается с фосфорилированным Sld2. Оба этих взаимодействия необходимы для CDK-зависимой активации почкования ДНК у дрожжей. [84]
Dpb11 также взаимодействует с GINS и участвует в стадиях инициации и элонгации репликации хромосомной ДНК. [60] [85] [86] GINS являются одним из белков репликации, обнаруженных в репликационных вилках, и образуют комплекс с Cdc45 и Mcm.
Эти зависимые от фосфорилирования взаимодействия между Dpb11, Sld2 и Sld3 необходимы для CDK-зависимой активации репликации ДНК, и с помощью реагентов сшивания в некоторых экспериментах был идентифицирован хрупкий комплекс, названный комплексом предварительной загрузки (pre-LC). . Этот комплекс содержит Pol ɛ, GINS, Sld2 и Dpb11. Обнаружено, что пре-LC образуется до какой-либо ассоциации с ориджинами CDK-зависимым и DDK-зависимым образом, а активность CDK регулирует инициацию репликации ДНК посредством образования пре-LC. [87]
Удлинение
[ редактировать ]
Образование пререпликативного комплекса (пре-РК) отмечает потенциальные сайты инициации репликации ДНК. В соответствии с комплексом поддержания минихромосомы, окружающим двухцепочечную ДНК, образование пре-РК не приводит к немедленному раскручиванию исходной ДНК или привлечению ДНК-полимераз. Вместо этого пре-РК, который образуется во время G 1 клеточного цикла, активируется только для раскручивания ДНК и инициации репликации после того, как клетки переходят из фазы G 1 в фазу S клеточного цикла. [2]
Как только инициирующий комплекс сформирован и клетки переходят в S-фазу, комплекс становится реплисомой. Реплисомный комплекс эукариот отвечает за координацию репликации ДНК. Репликация ведущей и отстающей цепей осуществляется ДНК-полимеразой ε и ДНК-полимеразой δ. Многие реплисомные факторы, включая Claspin, And1, загрузчик зажима фактора репликации C и комплекс защиты вилки, ответственны за регуляцию функций полимеразы и координацию синтеза ДНК с раскручиванием цепи матрицы комплексом Cdc45-Mcm-GINS. По мере раскручивания ДНК число скручиваний уменьшается. Чтобы компенсировать это, число корчи увеличивается, создавая положительные суперспирали в ДНК. Эти суперспирали привели бы к остановке репликации ДНК, если бы их не удалить. Топоизомеразы ответственны за удаление этих суперспиралей перед репликационной вилкой.
Реплисома отвечает за копирование всей геномной ДНК в каждой пролиферативной клетке. Реакции спаривания оснований и образования цепи, образующие дочернюю спираль, катализируются ДНК-полимеразами. [88] Эти ферменты движутся по одноцепочечной ДНК и позволяют удлинить зарождающуюся цепь ДНК, «читая» цепь матрицы и позволяя включить соответствующие пуриновые основания , аденин и гуанин , а также пиримидиновые нуклеиновые основания, тимин и цитозин . Активированные свободные дезоксирибонуклеотиды существуют в клетке в виде дезоксирибонуклеотидтрифосфатов (дНТФ). Эти свободные нуклеотиды добавляются к открытой 3'-гидроксильной группе последнего включенного нуклеотида. В этой реакции пирофосфат высвобождается из свободного dNTP, генерируя энергию для реакции полимеризации и обнажая 5'-монофосфат, который затем ковалентно связывается с 3'-кислородом. Кроме того, неправильно вставленные нуклеотиды могут быть удалены и заменены правильными нуклеотидами в энергетически выгодной реакции. Это свойство жизненно важно для правильной корректуры и исправления ошибок, возникающих во время репликации ДНК.
Репликационная вилка
[ редактировать ]Репликационная вилка — это соединение между вновь разделенными цепями матрицы, известными как лидирующая и отстающая цепи, и двухцепочечной ДНК. Поскольку дуплексная ДНК антипараллельна, репликация ДНК происходит в противоположных направлениях.между двумя новыми цепями в репликационной вилке, но все ДНК-полимеразы синтезируют ДНК в направлении от 5' к 3' относительно вновь синтезированной цепи. Во время репликации ДНК требуется дальнейшая координация. Две репликативные полимеразы синтезируют ДНК в противоположных ориентациях. Полимераза ε непрерывно синтезирует ДНК на «ведущей» цепи ДНК, поскольку она указывает в том же направлении, что и раскручивание ДНК реплисомой. Напротив, полимераза δ синтезирует ДНК на «отстающей» цепи, которая является противоположной цепью матрицы ДНК, фрагментированным или прерывистым образом.
Прерывистые участки продуктов репликации ДНК на отстающей цепи известны как фрагменты Оказаки и имеют длину от 100 до 200 оснований на вилках репликации эукариот. Отстающая цепь обычно содержит более длинные участки одноцепочечной ДНК, покрытые одноцепочечными связывающими белками, которые помогают стабилизировать одноцепочечные матрицы, предотвращая образование вторичной структуры. У эукариот эти одноцепочечные связывающие белки представляют собой гетеротримерный комплекс, известный как белок репликации А (RPA). [89]
Каждому фрагменту Оказаки предшествует праймер РНК, который замещается процессией следующего фрагмента Оказаки во время синтеза. РНКаза H распознает гибриды ДНК:РНК, которые создаются с использованием праймеров РНК, и отвечает за их удаление из реплицируемой цепи, оставляя после себя соединение праймер:матрица. ДНК-полимераза α распознает эти сайты и удлиняет разрывы, оставшиеся после удаления праймера. В эукариотических клетках небольшое количество сегмента ДНК непосредственно перед РНК-праймером также смещается, создавая лоскутную структуру. Этот лоскут затем расщепляется эндонуклеазами. В репликационной вилке разрыв в ДНК после удаления лоскута закрывается ДНК-лигазой I , которая восстанавливает разрывы, оставшиеся между 3'-ОН и 5'-фосфатом вновь синтезированной цепи. [90] Из-за относительно короткой природы эукариотического фрагмента Оказаки синтез репликации ДНК, происходящий прерывисто на отстающей цепи, менее эффективен и требует больше времени, чем синтез ведущей цепи. Синтез ДНК завершается после удаления всех праймеров РНК и устранения разрывов.

Ведущая нить
[ редактировать ]Во время репликации ДНК реплисома раскручивает родительский дуплекс ДНК в двухцепочечную репликационную вилку матрицы ДНК в направлении от 5' к 3'. Ведущая цепь — это шаблонная цепь, которая реплицируется в том же направлении, что и движение репликационной вилки. Это позволяет вновь синтезированной цепи, комплементарной исходной, синтезироваться от 5' к 3' в том же направлении, что и движение репликационной вилки. [91]
После того, как праймер РНК был добавлен примазой к 3'-концу ведущей цепи, синтез ДНК будет продолжаться в направлении от 3' к 5' относительно ведущей цепи без перерыва. ДНК-полимераза ε будет постоянно добавлять нуклеотиды к цепи матрицы, поэтому для синтеза ведущей цепи требуется только один праймер и она имеет непрерывную активность ДНК-полимеразы. [92]
Отстающая прядь
[ редактировать ]Репликация ДНК на отстающей цепи прерывистая. При отстающем синтезе цепи движение ДНК-полимеразы в направлении, противоположном репликационной вилке, требует использования нескольких праймеров РНК . ДНК-полимераза синтезирует короткие фрагменты ДНК, называемые фрагментами Оказаки , которые добавляются к 3'-концу праймера. Эти фрагменты у эукариот могут иметь длину от 100 до 400 нуклеотидов. [93]
В конце синтеза фрагмента Оказаки ДНК-полимераза δ сталкивается с предыдущим фрагментом Оказаки и вытесняет его 5'-конец, содержащий РНК-праймер и небольшой сегмент ДНК. При этом образуется одноцепочечный лоскут РНК-ДНК, который необходимо расщепить, а разрыв между двумя фрагментами Оказаки должен быть запечатан ДНК-лигазой I. Этот процесс известен как созревание фрагмента Оказаки и может осуществляться двумя способами: один механизм процессов короткие закрылки, а другой — длинные закрылки. [94] ДНК-полимераза δ способна смещать до 2–3 нуклеотидов ДНК или РНК перед ее полимеризацией, создавая короткий субстрат «лоскута» для Fen1 , который может удалять нуклеотиды из лоскута по одному нуклеотиду за раз.
Повторяя циклы этого процесса, ДНК-полимераза δ и Fen1 могут координировать удаление праймеров РНК и оставлять разрыв ДНК на отстающей цепи. [95] Было высказано предположение, что этот итерационный процесс предпочтительнее клеточного, поскольку он тесно связан.регулируется и не образует больших лоскутов, которые необходимо иссечь. [96] В случае нарушения регуляции активности Fen1/ДНК-полимеразы δ клетка использует альтернативный механизм для генерации и обработки длинных лоскутов с использованием ДНК2, которая обладает как хеликазной, так и нуклеазной активностью. [97] Нуклеазная активность Dna2 необходима для удаления этих длинных лоскутов, оставляя более короткий лоскут для обработки Fen1. Электронно-микроскопические исследования показывают, что загрузка нуклеосомы на отстающую цепь происходит очень близко к месту синтеза. [98] Таким образом, созревание фрагмента Окадзаки представляет собой эффективный процесс, который происходит сразу после синтеза зарождающейся ДНК.
Репликативные ДНК-полимеразы
[ редактировать ]После того, как репликативная хеликаза раскрутила дуплекс родительской ДНК, обнажив две одноцепочечные матрицы ДНК, необходимы репликативные полимеразы для создания двух копий родительского генома. Функция ДНК-полимеразы узкоспециализирована и обеспечивает репликацию на определенных матрицах и в узких локализациях. В репликационной вилке эукариот имеются три различных репликативных полимеразных комплекса, которые способствуют репликации ДНК: полимераза α, полимераза δ и полимераза ε. Эти три полимеразы необходимы для жизнеспособности клетки. [99]
Поскольку ДНК-полимеразы требуют праймера для начала синтеза ДНК, полимераза α (Pol α) действует как репликативная примаза. Pol α связан с РНК-примазой, и этот комплекс выполняет задачу праймирования путем синтеза праймера, который содержит короткий участок РНК длиной 10 нуклеотидов, за которым следуют от 10 до 20 оснований ДНК. [3] Важно отметить, что это праймирующее действие происходит при инициации репликации в ориджине, чтобы начать синтез ведущей цепи, а также на 5'-конце каждого фрагмента Оказаки на отстающей цепи.
Однако Pol α не способен продолжать репликацию ДНК и должен быть заменен другой полимеразой для продолжения синтеза ДНК. [100] Для переключения полимеразы необходимы загрузчики зажимов, и было доказано, что нормальная репликация ДНК требует скоординированных действий всех трех ДНК-полимераз: Pol α для синтеза прайминга, Pol ε для репликации ведущей цепи и Pol δ, которая постоянно нагружена, для генерации Фрагменты Оказаки во время синтеза отстающей цепи. [101]
- Полимераза α (Pol α) : образует комплекс с малой каталитической субъединицей (PriS) и большой некаталитической субъединицей (PriL). [102] Во-первых, синтез праймера РНК позволяет синтезировать ДНК с помощью ДНК-полимеразы альфа. Происходит один раз в начале координат ведущей цепи и в начале каждого фрагмента Оказаки на отстающей цепи. Субъединицы Pri действуют как примаза, синтезируя праймер РНК. ДНК Pol α удлиняет новообразованный праймер нуклеотидами ДНК. Примерно через 20 нуклеотидов элонгация берет на себя Pol ε на ведущей цепи и Pol δ на отстающей цепи. [103]
- Полимераза δ (Pol δ) : Высокая процессивность, корректура, 3’->5’ экзонуклеазная активность. In vivo это основная полимераза, участвующая в синтезе как отстающей , так и ведущей цепи. [104]
- Полимераза ε (Pol ε) : Высокая процессивность, корректура, 3'->5' экзонуклеазная активность. Тесно связанный с pol δ, in vivo он действует главным образом при проверке ошибок pol δ. [104]
Хеликазный комплекс Cdc45–Mcm–GINS
[ редактировать ]ДНК- хеликазы и полимеразы должны оставаться в тесном контакте на репликационной вилке. Если раскручивание происходит слишком далеко до синтеза, обнажаются большие участки одноцепочечной ДНК. Это может активировать передачу сигналов о повреждении ДНК или вызвать процессы восстановления ДНК. Чтобы решить эти проблемы, эукариотическая реплисома содержит специализированные белки, которые предназначены для регулирования активности геликазы перед репликационной вилкой. Эти белки также обеспечивают места стыковки для физического взаимодействия между хеликазами и полимеразами, тем самым гарантируя, что раскручивание дуплекса сопровождается синтезом ДНК. [105]
Чтобы ДНК-полимеразы функционировали, двухцепочечную спираль ДНК необходимо раскрутить, чтобы открыть две одноцепочечные матрицы ДНК для репликации. ДНК-хеликазы отвечают за раскручивание двухцепочечной ДНК во время репликации хромосом. Геликазы в эукариотических клетках чрезвычайно сложны. [106] Каталитическое ядро геликазы состоит из шести белков обслуживания минихромосом (Mcm2-7), образующих гексамерное кольцо. Вдали от ДНК белки Mcm2-7 образуют одиночный гетерогексамер и загружаются в неактивной форме в начале репликации ДНК в виде встречных двойных гексамеров вокруг двухцепочечной ДНК. [106] [107] Белки Mcm рекрутируются в места начала репликации, а затем перераспределяются по геномной ДНК во время S-фазы, что указывает на их локализацию в репликационной вилке. [54]
Загрузка белков Mcm может происходить только во время G1 клеточного цикла, а затем загруженный комплекс активируется во время S-фазы за счет рекрутирования белка Cdc45 и комплекса GINS с образованием активной геликазы Cdc45-Mcm-GINS (CMG) в Вилки репликации ДНК. [62] [108] Активность Mcm необходима на протяжении всей S-фазы для репликации ДНК. [38] [109] Вокруг геликазы CMG собираются различные регуляторные факторы, образующие «комплекс прогрессии реплисом», который связывается с ДНК-полимеразами и образует эукариотическую реплисому, структура которой еще довольно плохо определена по сравнению с ее бактериальным аналогом. [59] [110]
Изолированная геликаза CMG и комплекс прогрессии реплисомы содержат один белковый кольцевой комплекс Mcm, что позволяет предположить, что нагруженный двойной гексамер белков Mcm в начале процесса может расщепляться на два одиночных гексамерных кольца как часть процесса инициации, при этом каждое кольцо белкового комплекса Mcm образует ядро геликазы CMG на двух репликационных вилках, установленных от каждого источника. [59] [62] Полный комплекс CMG необходим для раскручивания ДНК, а комплекс CDC45-Mcm-GINS является функциональной ДНК-хеликазой в эукариотических клетках. [64]
Белки Ctf4 и And1
[ редактировать ]Комплекс CMG взаимодействует с реплисомой посредством взаимодействия с белками Ctf4 и And1. Белки Ctf4/And1 взаимодействуют как с комплексом CMG, так и с ДНК-полимеразой α. [111] Ctf4 представляет собой вспомогательный фактор полимеразы α, который необходим для привлечения полимеразы α к местам начала репликации. [112]
Белки Mrc1 и Claspin
[ редактировать ]Белки Mrc1/Claspin связывают синтез ведущей цепи с геликазной активностью комплекса CMG. Mrc1 взаимодействует с полимеразой ε, а также с белками Mcm. [113] Важность этой прямой связи между хеликазой и полимеразой ведущей цепи подчеркивается результатами, полученными в культивируемых клетках человека, где Mrc1/Claspin необходим для эффективного развития репликационной вилки. [114] Эти результаты позволяют предположить, что эффективная репликация ДНК также требует связывания геликаз и синтеза ведущей цепи...
Ядерный антиген пролиферирующих клеток
[ редактировать ]ДНК-полимеразы требуют дополнительных факторов для поддержки репликации ДНК. ДНК-полимеразы имеют полузакрытую структуру «рука», которая позволяет полимеразе загружаться в ДНК и начинать транслокацию. Эта структура позволяет ДНК-полимеразе удерживать матрицу одноцепочечной ДНК, включать dNTP в активный сайт и высвобождать вновь образованную двухцепочечную ДНК. Однако структура ДНК-полимераз не позволяет осуществлять постоянное стабильное взаимодействие с ДНК-матрицей. [1]
Чтобы усилить взаимодействие между полимеразой и ДНК-матрицей, скользящие зажимы ДНК связываются с полимеразой, способствуя процессивности репликативной полимеразы. У эукариот скользящий зажим представляет собой кольцевую структуру гомотримера, известную как ядерный антиген пролиферирующих клеток (PCNA). Кольцо PCNA имеет полярность с поверхностями, которые взаимодействуют с ДНК-полимеразами, и надежно привязывает их к матрице ДНК. PCNA-зависимая стабилизация ДНК-полимераз оказывает значительное влияние на репликацию ДНК, поскольку PCNA способны повышать процессивность полимеразы до 1000 раз. [115] [116] PCNA является важным кофактором и отличается тем, что является одной из наиболее распространенных платформ взаимодействия в реплисоме, обеспечивающей множественные процессы в репликационной вилке, поэтому PCNA также рассматривается как регуляторный кофактор для ДНК-полимераз. [117]
Фактор репликации C
[ редактировать ]PCNA полностью окружает цепь матрицы ДНК и должен быть загружен в ДНК в репликационной вилке. В ведущей цепи загрузка PCNA происходит нечасто, поскольку репликация ДНК на ведущей цепи продолжается до тех пор, пока репликация не завершится. Однако в отстающей цепи ДНК-полимераза δ должна постоянно загружаться в начале каждого фрагмента Оказаки. Эта постоянная инициация синтеза фрагментов Оказаки требует повторной загрузки PCNA для эффективной репликации ДНК.
Загрузка PCNA осуществляется комплексом фактора репликации C (RFC). Комплекс RFC состоит из пяти АТФаз: Rfc1, Rfc2, Rfc3, Rfc4 и Rfc5. [118] RFC распознает соединения праймер-матрица и загружает PCNA в эти сайты. [119] [120] Гомотример PCNA открывается с помощью RFC путем гидролиза АТФ, а затем загружается в ДНК в правильной ориентации, чтобы облегчить его связь с полимеразой. [121] [122] Загрузчики зажимов также могут выгружать PCNA из ДНК; механизм, необходимый, когда репликация должна быть прекращена. [122]
Остановившаяся вилка репликации
[ редактировать ]Репликация ДНК в репликационной вилке может быть остановлена из-за нехватки дезоксинуклеотидтрифосфатов (dNTP) или повреждения ДНК, что приводит к репликационному стрессу . [123] Эта остановка репликации описывается как остановленная вилка репликации . Комплекс белков, защищающий вилку, стабилизирует репликационную вилку до тех пор, пока не будут устранены повреждения ДНК или другие проблемы репликации. [123] Длительная остановка репликационной вилки может привести к дальнейшему повреждению ДНК. Сигналы остановки деактивируются, если устранены проблемы, вызывающие вилку репликации. [123]
Прекращение действия
[ редактировать ]
Прекращение репликации эукариотической ДНК требует разных процессов в зависимости от того, являются ли хромосомы кольцевыми или линейными. В отличие от линейных молекул, кольцевые хромосомы способны воспроизводить всю молекулу. Однако две молекулы ДНК останутся связанными друг с другом. Эта проблема решается путем декатенации двух молекул ДНК топоизомеразой типа II . Топоизомеразы типа II также используются для разделения линейных нитей, поскольку они сложно свернуты в нуклеосому внутри клетки.
Как упоминалось ранее, линейные хромосомы сталкиваются с еще одной проблемой, которая не наблюдается при кольцевой репликации ДНК. Поскольку для инициации синтеза ДНК необходим праймер РНК, отстающая цепь оказывается в невыгодном положении при репликации всей хромосомы. В то время как ведущая цепь может использовать один праймер РНК для удлинения 5'-конца реплицирующейся цепи ДНК, множественные праймеры РНК ответственны за синтез отстающей цепи, создавая фрагменты Оказаки. Это приводит к проблеме, связанной с тем, что ДНК-полимераза способна добавлять только к 3'-концу цепи ДНК. Действие 3'-5' ДНК-полимеразы вдоль родительской цепи оставляет короткую одноцепочечную область ДНК (оцДНК) на 3'-конце родительской цепи после репарации фрагментов Оказаки. Поскольку репликация происходит в противоположных направлениях на противоположных концах родительских хромосом, каждая цепь на одном конце является отстающей. Со временем это приведет к прогрессивному укорочению обеих дочерних хромосом . Это известно как проблема конечной репликации. [1]
Проблема конечной репликации решается в эукариотических клетках теломерными участками и теломеразой . Теломеры расширяют 3'-конец родительской хромосомы за 5'-конец дочерней цепи. Эта одноцепочечная структура ДНК может действовать как точка начала репликации, которая рекрутирует теломеразу. Теломераза — это специализированная ДНК-полимераза, состоящая из множества белковых субъединиц и компонента РНК. РНК-компонент теломеразы отжигается с одноцепочечным 3'-концом матричной ДНК и содержит 1,5 копии теломерной последовательности. [93] Теломераза содержит белковую субъединицу, которая представляет собой обратную транскриптазу, называемую обратной транскриптазой теломеразы или TERT. TERT синтезирует ДНК до конца матричной теломеразной РНК, а затем отделяется. [93] Этот процесс можно повторять столько раз, сколько необходимо, с удлинением 3'-конца родительской молекулы ДНК. Это 3'-добавление обеспечивает матрицу для удлинения 5'-конца дочерней цепи за счет задержки синтеза ДНК цепи. Регуляция активности теломеразы осуществляется белками, связывающими теломеры.
Барьеры репликационной вилки
[ редактировать ]Репликация ДНК прокариот двунаправленная; В пределах репликативного начала реплисомные комплексы создаются на каждом конце начала репликации, и реплисомы удаляются друг от друга от начальной начальной точки. У прокариот двунаправленная репликация начинается в одном месте начала репликации на кольцевой хромосоме и заканчивается в сайте, противоположном начальному началу начала точки начала. [124] Эти области терминации содержат последовательности ДНК, известные как Ter сайты . Эти сайты Ter связаны с белком Tus. Комплекс Тер -Тус способен останавливать активность геликазы, прекращая репликацию. [125]
В эукариотических клетках прекращение репликации обычно происходит в результате столкновения двух репликативных вилок между двумя активными точками начала репликации. Место столкновения зависит от времени начала стрельбы. Таким образом, если репликационная вилка останавливается или разрушается на определенном сайте, репликация сайта может быть восстановлена, когда реплисома, движущаяся в противоположном направлении, завершит копирование региона. Существуют запрограммированные барьеры репликационной вилки (RFB), связанные белками RFB в различных местах по всему геному, которые способны терминировать или приостанавливать репликационные вилки, останавливая прогрессирование реплисомы. [124]
Фабрики репликации
[ редактировать ]Было обнаружено, что репликация происходит локализованно в ядре клетки. Вопреки традиционному представлению о движении репликационных вилок вдоль застойной ДНК, возникла концепция репликационных фабрик , которая означает, что репликационные вилки концентрируются в некоторых иммобилизованных «фабричных» регионах, через которые нити матричной ДНК проходят, как конвейерные ленты. [126]
Регуляция клеточного цикла
[ редактировать ]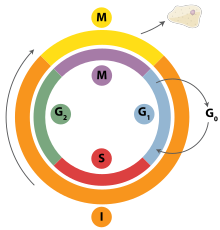
Репликация ДНК — это четко организованный процесс, который контролируется в контексте клеточного цикла . Прогресс клеточного цикла и, в свою очередь, репликация ДНК жестко регулируется образованием и активацией пререпликативных комплексов (пре-РК), что достигается за счет активации и инактивации циклин-зависимых киназ (Cdks, CDK). В частности, именно взаимодействия циклинов и циклинзависимых киназ ответственны за переход из G 1 в S-фазу.
Во время фазы G1 клеточного цикла наблюдается низкий уровень активности CDK. Этот низкий уровень активности CDK позволяет образовывать новые комплексы пре-РК, но недостаточен для инициации репликации ДНК вновь образованными пре-РК. Во время остальных фаз клеточного цикла наблюдается повышенный уровень активности CDK. Этот высокий уровень активности CDK отвечает за инициацию репликации ДНК, а также за ингибирование образования новых комплексов pre-RC. [2] После начала репликации ДНК комплекс pre-RC разрушается. В связи с тем, что уровни CDK остаются высокими во время фаз S, G 2 и M клеточного цикла, новые комплексы pre-RC не могут образовываться. Все это помогает гарантировать, что инициация не может произойти до тех пор, пока деление клеток не завершится.
Считается, что в дополнение к циклин-зависимым киназам новый раунд репликации предотвращается за счет подавления Cdt1. Это достигается за счет деградации Cdt1, а также за счет ингибирующего действия белка, известного как геминин . Геминин прочно связывается с Cdt1 и считается основным ингибитором повторной репликации. [2] Геминин впервые появляется в S-фазе и разрушается при переходе метафаза-анафаза, возможно, за счет убиквининирования комплексом, способствующим анафазе (APC). [127]
присутствуют различные клеточного цикла На протяжении всего контрольные точки клеточного цикла, которые определяют, сможет ли клетка полностью пройти процесс деления. Важно отметить, что при репликации контрольная точка G 1 , или контрольная точка ограничения, определяет, начнется ли инициация репликации или будет ли клетка помещена в стадию покоя, известную как G 0 . Клетки на стадии G0 клеточного цикла не могут начать цикл репликации, поскольку белки поддержания минихромосом не экспрессируются. Переход в S-фазу указывает на начало репликации.
Белки контрольной точки репликации
[ редактировать ]Чтобы сохранить генетическую информацию во время деления клеток, репликация ДНК должна выполняться с высокой точностью. Для решения этой задачи в эукариотических клетках в определенные моменты процесса репликации присутствуют белки, которые способны обнаруживать любые ошибки во время репликации ДНК и сохранять целостность генома. Эти белки контрольных точек способны остановить вступление клеточного цикла в митоз, чтобы дать время для восстановления ДНК. Белки контрольных точек также участвуют в некоторых путях восстановления ДНК, при этом они стабилизируют структуру репликационной вилки, предотвращая дальнейшее повреждение. Эти белки контрольных точек необходимы для предотвращения передачи мутаций или других хромосомных аберраций потомству.
Эукариотические белки контрольных точек хорошо консервативны и включают две фосфатидилинозитол-3-киназные киназы (PIKK), ATR и ATM . И ATR, и ATM имеют общую целевую последовательность фосфорилирования, мотив SQ/TQ, но их индивидуальные роли в клетках различаются. [128]
ATR участвует в остановке клеточного цикла в ответ на двухцепочечные разрывы ДНК. У ATR есть обязательный партнер контрольной точки, ATR-взаимодействующий белок (ATRIP), и вместе эти два белка реагируют на участки одноцепочечной ДНК, покрытые репликационным белком A (RPA). [129] Образование одноцепочечной ДНК происходит часто, чаще во время репликационного стресса. ATR-ATRIP способен остановить клеточный цикл, чтобы сохранить целостность генома. ATR обнаруживается на хроматине во время S-фазы, подобно RPA и класпину. [130]
Генерация одноцепочечных участков ДНК важна для инициации путей контрольных точек после повреждения репликации. Как только одноцепочечная ДНК становится достаточно длинной, одноцепочечная ДНК, покрытая RPA, может рекрутировать ATR-ATRIP. [131] Чтобы стать полностью активной, киназа ATR полагается на сенсорные белки, которые определяют, локализованы ли белки контрольной точки в действительном сайте стресса репликации ДНК. Гетеротримерный зажим RAD9 ( 9-1-1 - HUS1 - Rad1 ) и его загрузчик RFC. Рад17 способны распознавать разрывы или разрывы ДНК. RFC Рад17 загрузчик зажимов загружает 9-1-1 в поврежденную ДНК. [132] Присутствия 9-1-1 на ДНК достаточно, чтобы облегчить взаимодействие между ATR-ATRIP и группой белков, называемых медиаторами контрольных точек, таких как TOPBP1 и Mrc1/ класпин . TOPBP1 взаимодействует с фосфорилированным компонентом Rad9 9-1-1 и рекрутирует его, а также связывает ATR-ATRIP, который фосфорилирует Chk1. [133] Mrc1/Claspin также необходим дляполная активация ATR-ATRIP, которая фосфорилирует Chk1, основную эффекторную киназу нижележащей контрольной точки. [134] Класпин является компонентом реплисомы и содержит домен для стыковки с Chk1, что раскрывает специфическую функцию класпина во время репликации ДНК: стимулированиесигнализация контрольной точки на реплисоме. [135]
Передача сигналов Chk1 жизненно важна для остановки клеточного цикла и предотвращения вступления клеток в митоз с неполной репликацией ДНК или повреждением ДНК. Chk1-зависимое ингибирование Cdk важно для функции контрольной точки ATR-Chk1, а также для остановки клеточного цикла и обеспечения достаточного времени для завершения механизмов репарации ДНК, что, в свою очередь, предотвращает наследование поврежденной ДНК. Кроме того, Chk1-зависимое ингибирование Cdk играет критическую роль в ингибировании активации источника во время S-фазы. Этот механизм предотвращает дальнейший синтез ДНК и необходим для защиты генома вналичие репликационного стресса и потенциальных генотоксических состояний. [136] Таким образом, активность ATR-Chk1 дополнительно предотвращает потенциальные проблемы репликации на уровне одиночных точек начала репликации, ингибируя инициацию репликации по всему геному до тех пор, пока не отключится сигнальный каскад, поддерживающий остановку клеточного цикла.
Репликация через нуклеосомы
[ редактировать ]
Эукариотическая ДНК должна быть плотно сжата, чтобы поместиться в ограниченном пространстве ядра. Хромосомы упаковываются путем обертывания 147 нуклеотидов вокруг октамера белков- гистонов , образуя нуклеосому . Нуклеосомный октамер включает по две копии каждого гистона H2A , H2B , H3 и H4 . Из-за тесной связи белков-гистонов с ДНК эукариотические клетки имеют белки, которые предназначены для ремоделирования гистонов перед репликационной вилкой, чтобы обеспечить плавное развитие реплисомы. [137] Существуют также белки, участвующие в повторной сборке гистонов за репликационной вилкой для восстановления конформации нуклеосомы. [138]
Известно несколько гистонов-шаперонов, которые участвуют в сборке нуклеосом после репликации. Было обнаружено, что комплекс FACT взаимодействует с комплексом ДНК-полимеразы α-примаза, а субъединицы комплекса FACT генетически взаимодействуют с факторами репликации. [139] [140] Комплекс FACT представляет собой гетеродимер, который не гидролизует АТФ, но способен способствовать «разрыхлению» гистонов в нуклеосомах, но как комплекс FACT способен разряжать тесную ассоциацию гистонов для удаления ДНК, остается без ответа. [141]
Другим шапероном гистонов, который связывается с реплисомой, является Asf1 , который взаимодействует с комплексом Mcm, зависимым от димеров гистонов H3-H4. [142] Asf1 способен передавать вновь синтезированный димер H3-H4 факторам отложения позади репликационной вилки, и эта активность делает димеры гистонов H3-H4 доступными в месте отложения гистонов сразу после репликации. [143] Asf1 (и его партнер Rtt109) также участвует в ингибировании экспрессии генов из реплицируемых генов во время S-фазы. [144]
Фактор сборки гетеротримерного шаперона хроматина 1 ( CAF-1 ) представляет собой белок, образующий хроматин, который участвует в отложении гистонов на обеих вновь реплицированных цепях ДНК с образованием хроматина. [145] CAF-1 содержит мотив связывания PCNA, называемый PIP-боксом, который позволяет CAF-1 связываться с реплисомой через PCNA и способен откладывать димеры гистонов H3-H4 на вновь синтезированную ДНК. [146] [147] Шаперон Rtt106 также участвует в этом процессе и связан с димерами CAF-1 и H3-H4 во время образования хроматина. [148] Эти процессы загружают вновь синтезированные гистоны в ДНК.
После отложения гистонов H3-H4 нуклеосомы образуются за счет ассоциации гистонов H2A-H2B. Считается, что этот процесс происходит через комплекс FACT, поскольку он уже связан с реплисомой и способен связывать свободные H2A-H2B, или существует возможность другого шаперона H2A-H2B, Nap1. [149] Исследования электронной микроскопии показывают, что это происходит очень быстро, поскольку можно наблюдать, как нуклеосомы образуют всего несколько сотен пар оснований после репликационной вилки. [150] Поэтому весь процесс формирования новыхнуклеосомы происходит сразу после репликации из-за связывания шаперонов гистонов с реплисомой.
Сравнение репликации ДНК прокариот и эукариот
[ редактировать ]По сравнению с репликацией ДНК прокариот , а именно у бактерий, завершение репликации ДНК эукариот является более сложным и включает в себя несколько точек начала репликации и репликативные белки. ДНК прокариот имеет круглую форму и в момент начала репликации имеет только одно начало репликации. Напротив, эукариотическая ДНК линейна. При репликации существует до тысячи источников репликации. [151]
Эукариотическая ДНК двунаправлена. Здесь значение слова двунаправленный другой. Эукариотическая линейная ДНК имеет множество начальных точек (называемых О) и концов (называемых Т). «Т» находится справа от «О». Одна «О» и одна «Т» вместе образуют один репликон. После образования преинициационного комплекса, когда один репликон начинает элонгацию, инициация начинается во втором репликоне. Теперь, если первый репликон движется по часовой стрелке, второй репликон движется против часовой стрелки, пока не будет достигнута точка «T» первого репликона. В точке «Т» оба репликона сливаются, завершая процесс репликации. Тем временем второй репликон также движется вперед, чтобы встретиться с третьим репликоном. Это движение двух репликонов по часовой стрелке и против часовой стрелки называется двунаправленной репликацией.
Репликация эукариотической ДНК требует точной координации всех ДНК-полимераз и связанных с ними белков для репликации всего генома каждый раз, когда клетка делится. Этот процесс достигается за счет серии этапов сборки белков в точках начала репликации, при этом регуляция репликации ДНК в основном фокусируется на ассоциации хеликазы MCM с ДНК. Эти точки начала репликации определяют количество белковых комплексов, которые образуются для инициации репликации. При репликации бактериальной ДНК регуляция фокусируется на связывании белка-инициатора DnaA с ДНК, при этом инициация репликации происходит несколько раз в течение одного клеточного цикла. [93] И прокариотическая, и эукариотическая ДНК используют связывание и гидролиз АТФ для направления загрузки геликазы, и в обоих случаях геликаза загружается в неактивной форме. Однако эукариотические хеликазы представляют собой двойные гексамеры, загруженные на двухцепочечную ДНК, тогда как бактериальные хеликазы представляют собой одиночные гексамеры, загруженные на одноцепочечную ДНК. [152]
Сегрегация хромосом — еще одно различие между прокариотическими и эукариотическими клетками. Быстро делящиеся клетки, такие как бактерии, часто начинают разделять хромосомы, которые все еще находятся в процессе репликации. В эукариотических клетках разделение хромосом на дочерние клетки не начинается до тех пор, пока не завершится репликация во всех хромосомах. [93] Однако, несмотря на эти различия, основной процесс репликации одинаков как для прокариотической, так и для эукариотической ДНК.
| Репликация бактериальной ДНК | Репликация эукариотической ДНК |
|---|---|
| Происходит внутри цитоплазмы | Происходит внутри ядра |
| Только один центр репликации на молекулу ДНК. | Имеют много точек начала репликации в каждой хромосоме. |
| Начало репликации имеет длину около 100-200 и более нуклеотидов. | Каждый ориджин репликации состоит примерно из 150 нуклеотидов. |
| Репликация происходит в одной точке каждой хромосомы | Репликация происходит одновременно в нескольких точках каждой хромосомы. |
| Имеют только один источник репликации | Имеет несколько источников репликации. |
| Инициация осуществляется белками DnaA и DnaB. | Инициация осуществляется Комплексом распознавания происхождения. |
| Нужна топоизомераза | Нужна топоизомераза |
| Репликация очень быстрая | Репликация очень медленная |
Список белков репликации эукариотической ДНК
[ редактировать ]Список основных белков, участвующих в репликации ДНК эукариот:
| Белок | Функция репликации эукариотической ДНК |
|---|---|
| И1 | Нагружает ДНК-полимеразу α на хроматин вместе с комплексом CMG на отстающей цепи. Также известен как Ctf4 у почкующихся дрожжей. |
| Cdc45 | Необходим для этапов инициации и элонгации репликации ДНК. Часть геликазного комплекса Mcm2-7. Требуется после этапа pre-RC для загрузки различных белков для инициации и элонгации. |
| Комплекс Cdc45-Mcm-ГИНС (ЦМГ) | Функциональная ДНК-геликаза в эукариотических клетках |
| Cdc6 | Требуется для сборки комплекса Mcm2-7 на ORC совместно с Cdt1. |
| Киназа Cdc7-Dbf4 или Dbf4-зависимая киназа (DDK) | Протеинкиназа необходима для инициации репликации ДНК, вероятно, посредством фосфорилирования поддерживающих белков минихромосомы. |
| Кдт1 | Загружает комплекс Mcm2-7 в ДНК в ORC на этапе предварительного RC/лицензирования. У многоклеточных животных ингибируется геминином. |
| Застежка | Соедините синтез ведущей цепи с геликазной активностью комплекса CMG. Работает с Mrc1 |
| Ctf4 | Нагружает ДНК-полимеразу α на хроматин вместе с комплексом CMG на отстающей цепи. Гомолог у многоклеточных животных известен как AND-1. |
| Циклинзависимая киназа (CDK) | Циклин-зависимая протеинкиназа необходима для инициации репликации и других последующих этапов. |
| ДНК2 | 5'-флоп-эндонуклеаза и геликаза участвуют в процессинге фрагментов Оказаки. |
| ДНК-лигаза I | Присоединяется к фрагментам Окадзаки во время репликации ДНК. Активность лигазы также необходима для репарации и рекомбинации ДНК. |
| ДНК-полимераза α (Pol α) | Содержит активность примазы, необходимую для инициации синтеза ДНК как на ведущей, так и на отстающей цепи. |
| ДНК-полимераза δ (Pol δ) | Требуется для завершения синтеза фрагментов Оказаки на отстающей цепи, начатого ДНК-полимеразой α. |
| ДНК-полимераза ε (Pol ε) | Полимераза ведущей цепи. Синтезирует ДНК на репликационной вилке. Связывается на ранних стадиях через Dbp11 и необходим для загрузки ДНК-полимеразы α. |
| Дпб11 | Белок инициации репликации ДНК. Загружает ДНК-полимеразу ε в пререпликационные комплексы в точках происхождения. |
| Фен1 | 5'-флоп-эндонуклеаза, участвующая в процессинге фрагментов Оказаки. |
| корабля | Белок обнаружен у многоклеточных животных и отсутствует в дрожжах. Связывается с Cdt1 и инактивирует его, тем самым регулируя образование пререпликативного/инициаторного комплекса. Также предложено способствовать образованию пре-RC путем связывания и, таким образом, предотвращения деградации Cdt1. |
| ДЖИНЫ | Тетрамерный комплекс, состоящий из Sld5, Psf1, Psf2, Psf3. Связывается с пререпликативным комплексом примерно во время инициации и перемещается вместе с репликативными вилками на этапе элонгации. Необходим для стадии элонгации репликации ДНК и, возможно, является частью хеликазного комплекса Mcm. |
| Белки поддержания минихромосомы (Mcm) | Шесть различных белков семейства ААА+-АТФазы, образующих в растворе гексамер. Этот гексамер рекрутируется и загружается с помощью ORC, Cdc6 и Cdt1 и образует двойной гексамер, который топологически связан вокруг ДНК с образованием солеустойчивого пререпликативного комплекса. При инициации репликации Mcm2-7 удаляется от ORC с помощью вилки репликации. |
| Мкм10 | Необходим для стадий инициации и элонгации репликации ДНК. Участвует в связывании хроматина Cdc45 и ДНК-полимеразы α. Также необходим для стабильности α-каталитической субъединицы ДНК-полимеразы в почкующихся дрожжах S. cerevisiae . |
| Mrc1 | Соедините синтез ведущей цепи с геликазной активностью комплекса CMG. Гомолог многоклеточных животных известен как Claspin. |
| Комплекс распознавания происхождения (ORC) | Гетерогексамерный комплекс, состоящий из белков Orc1–Orc6. Связывается с ДНК и собирает комплекс Mcm2-7 на хроматине вместе с Cdc6 и Cdt1. |
| Ядерный антиген пролиферирующих клеток (PCNA) | Тримерный белок кольцевой структуры окружает ДНК, предотвращая диссоциацию ДНК-полимеразы. Действует как скользящий зажим для полимераз δ и ε, тем самым улучшая процессивность репликативных полимераз. |
| Фактор репликации C (RFC) | Загружает PCNA на загрунтованные матрицы и участвует в переключении между ДНК-полимеразой a и репликативными полимеразами δ и ε. |
| Барьеры репликационной вилки (RFB) | Связывается белками RFB в различных местах генома. Способны прекращать или приостанавливать репликационные вилки, останавливая прогрессирование реплисомы. |
| Репликационный белок А (RPA) | Гетеротримерный одноцепочечный связывающий белок. Стабилизирует одноцепочечную ДНК в репликационной вилке. |
| РНКаза H | Рибонуклеаза, расщепляющая РНК, гибридизованную с ДНК. Участвовал в обработке фрагментов Оказаки. |
| Слд2 | Функции инициации репликации. Ключевой субстрат CDK, фосфорилирование способствует взаимодействию с Dpb11. Требуется для запуска репликации. |
| Слд3 | Функции инициации репликации. Ключевой субстрат CDK, фосфорилирование способствует взаимодействию с Dpb11. Требуется для запуска репликации. |
| Теломераза | Рибонуклеопротеин, добавляющий повторы последовательности ДНК «TTAGGG» к 3'-концу нитей ДНК в теломерах. |
| Топоизомеразы | Регулирует перекручивание или перекручивание ДНК |
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Леман А.Р., Ногучи Э. (март 2013 г.). «Репликационная вилка: понимание механизма репликации эукариот и проблем дупликации генома» . Гены . 4 (1): 1–32. дои : 10.3390/genes4010001 . ПМЦ 3627427 . ПМИД 23599899 .
- ^ Перейти обратно: а б с д Блоу Джей-Джей, Датта А. (июнь 2005 г.). «Предотвращение повторной репликации хромосомной ДНК» . Nature Reviews Молекулярно-клеточная биология . 6 (6): 476–86. дои : 10.1038/nrm1663 . ПМЦ 2688777 . ПМИД 15928711 .
- ^ Перейти обратно: а б Фишер П.А., Ван Т.С., Корн Д. (июль 1979 г.). «Энзимологическая характеристика ДНК-полимеразы альфа. Основные каталитические свойства, процессивность и утилизация гомогенного фермента из клеток KB человека» . Журнал биологической химии . 254 (13): 6128–37. дои : 10.1016/S0021-9258(18)50528-7 . ПМИД 447699 .
- ^ Боос Д., Феррейра П. (март 2019 г.). «Правила использования происхождения для контроля времени репликации генома» . Гены . 10 (3): 199. doi : 10.3390/genes10030199 . ПМК 6470937 . ПМИД 30845782 .
- ^ Фосс Э.Дж., Личауко К., Гатбонтон-Швагер Т., Лофтс Б., Лао У., Бедалов А. (03.07.2023). «Идентификация 1600 точек начала репликации у S. cerevisiae» . электронная жизнь . 12 : RP88087. doi : 10.7554/eLife.88087 . ПМЦ 10945306 . ПМИД 38315095 .
- ^ Араки Х (2011). «Инициация репликации хромосомной ДНК в эукариотических клетках; вклад генетики дрожжей в ее объяснение» . Гены и генетические системы . 86 (3): 141–9. дои : 10.1266/ggs.86.141 . ПМИД 21952204 .
- ^ Майорано Д., Моро Дж., Мечали М. (апрель 2000 г.). «XCDT1 необходим для сборки пререпликативных комплексов у Xenopus laevis». Природа . 404 (6778): 622–5. Бибкод : 2000Natur.404..622M . дои : 10.1038/35007104 . PMID 10766247 . S2CID 4416138 .
- ^ Перейти обратно: а б с Белл С.П., Дутта А. (2002). «Репликация ДНК в эукариотических клетках». Ежегодный обзор биохимии . 71 : 333–74. doi : 10.1146/annurev.biochem.71.110601.135425 . ПМИД 12045100 .
- ^ Тай Б.К. (1999). «Белки MCM в репликации ДНК». Ежегодный обзор биохимии . 68 (1): 649–86. doi : 10.1146/annurev.biochem.68.1.649 . ПМИД 10872463 .
- ^ Тикау С., Фридман Л.Дж., Ивица Н.А., Геллес Дж., Белл С.П. (апрель 2015 г.). «Одномолекулярные исследования лицензирования происхождения выявляют механизмы, обеспечивающие двунаправленную загрузку геликазы» . Клетка . 161 (3): 513–525. дои : 10.1016/j.cell.2015.03.012 . ПМЦ 4445235 . ПМИД 25892223 .
- ^ Перейти обратно: а б Чжай Ю, Ли Н, Цзян Х, Хуан Х, Гао Н, Тай БК (июль 2017 г.). «Уникальные роли неидентичных субъединиц MCM в лицензировании репликации ДНК» . Молекулярная клетка . 67 (2): 168–179. doi : 10.1016/j.molcel.2017.06.016 . ПМИД 28732205 .
- ^ Костер Дж., Диффли Дж. Ф. (июль 2017 г.). «Двунаправленная репликация эукариотической ДНК осуществляется за счет квазисимметричной загрузки геликазы» . Наука . 357 (6348): 314–318. Бибкод : 2017Sci...357..314C . дои : 10.1126/science.aan0063 . ПМК 5608077 . ПМИД 28729513 .
- ^ Перейти обратно: а б с д и Чжай Ю, Ченг Э, Ву Х, Ли Н, Юнг ПЯ, Гао Н, Тай БК (март 2017 г.). «Разомкнутая структура комплекса Cdt1-Mcm2-7 как предшественника двойного гексамера MCM». Структурная и молекулярная биология природы . 24 (3): 300–308. дои : 10.1038/nsmb.3374 . ПМИД 28191894 . S2CID 3929807 .
- ^ Bell SP (март 2002 г.). «Комплекс распознавания координат: от простых истоков к сложным функциям» . Гены и развитие . 16 (6): 659–72. дои : 10.1101/gad.969602 . ПМИД 11914271 .
- ^ Белл С.П., Стиллман Б. (май 1992 г.). «АТФ-зависимое распознавание эукариотического начала репликации ДНК мультибелковым комплексом». Природа . 357 (6374): 128–134. Бибкод : 1992Natur.357..128B . дои : 10.1038/357128a0 . ПМИД 1579162 . S2CID 4346767 .
- ^ Перейти обратно: а б с д и ж Ли Н., Лам В.Х., Чжай Ю., Ченг Дж., Ченг Э., Чжао Ю. и др. (июль 2018 г.). «Структура комплекса распознавания происхождения, связанного с началом репликации ДНК». Природа . 559 (7713): 217–222. Бибкод : 2018Natur.559..217L . дои : 10.1038/s41586-018-0293-x . ПМИД 29973722 . S2CID 49577101 .
- ^ Миотто Б., Джи З., Струл К. (август 2016 г.). «Селективность сайтов связывания ORC и связь со временем репликации, хрупкими сайтами и делециями при раке» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (33): Е4810-9. Бибкод : 2016PNAS..113E4810M . дои : 10.1073/pnas.1609060113 . ПМЦ 4995967 . ПМИД 27436900 .
- ^ Перейти обратно: а б Ли К.С., Чунг М.Ф., Ли Дж., Чжао Ю., Лам В.Х., Хо В. и др. (январь 2021 г.). «Гуманизация комплекса распознавания происхождения дрожжей» . Природные коммуникации . 12 (1): 33. Бибкод : 2021NatCo..12...33L . дои : 10.1038/s41467-020-20277-y . ПМЦ 7782691 . ПМИД 33397927 .
- ^ Бляйхерт Ф., Лейтнер А., Эберсольд Р., Ботчан М.Р., Бергер Дж.М. (июнь 2018 г.). «Конформационный контроль и механизм связывания ДНК комплекса распознавания происхождения многоклеточных животных» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (26): E5906–E5915. Бибкод : 2018PNAS..115E5906B . дои : 10.1073/pnas.1806315115 . ПМК 6042147 . ПМИД 29899147 .
- ^ Чесноков И.Н. (2007). «Множественные функции комплекса распознавания происхождения». Международный обзор цитологии . 256 : 69–109. дои : 10.1016/S0074-7696(07)56003-1 . ISBN 9780123737007 . ПМИД 17241905 .
- ^ Мацуда К., Макисе М., Суэясу Ю., Такехара М., Асано Т., Мидзушима Т. (декабрь 2007 г.). «Двугибридный дрожжевой анализ комплекса распознавания происхождения Saccharomyces cerevisiae: взаимодействие между субъединицами и идентификация связывающих белков» . Исследование дрожжей FEMS . 7 (8): 1263–9. дои : 10.1111/j.1567-1364.2007.00298.x . ПМИД 17825065 .
- ^ Крейц С., Ритци М., Баак М., Книпперс Р. (март 2001 г.). «Белок 1 комплекса распознавания человеческого происхождения диссоциирует от хроматина во время S-фазы в клетках HeLa» . Журнал биологической химии . 276 (9): 6337–42. дои : 10.1074/jbc.M009473200 . ПМИД 11102449 .
- ^ Спек С., Чен З., Ли Х., Стиллман Б. (ноябрь 2005 г.). «АТФаза-зависимое кооперативное связывание ORC и Cdc6 с исходной ДНК» . Структурная и молекулярная биология природы . 12 (11): 965–71. дои : 10.1038/nsmb1002 . ПМЦ 2952294 . ПМИД 16228006 .
- ^ Коулман Т.Р., Карпентер П.Б., Данфи В.Г. (октябрь 1996 г.). «Белок Xenopus Cdc6 необходим для инициации одного раунда репликации ДНК в бесклеточных экстрактах» . Клетка . 87 (1): 53–63. дои : 10.1016/S0092-8674(00)81322-7 . ПМИД 8858148 . S2CID 16897247 .
- ^ Риалланд М., Сола Ф., Сантоканале К. (апрель 2002 г.). «Основная роль CDT1 человека в репликации ДНК и лицензировании хроматина». Журнал клеточной науки . 115 (Часть 7): 1435–40. дои : 10.1242/jcs.115.7.1435 . ПМИД 11896191 .
- ^ Танака С., Диффли Дж. Ф. (март 2002 г.). «Взаимозависимое ядерное накопление почкующихся дрожжей Cdt1 и Mcm2-7 во время фазы G1». Природная клеточная биология . 4 (3): 198–207. дои : 10.1038/ncb757 . ПМИД 11836525 . S2CID 45861829 .
- ^ Фригола Дж., Хе Дж., Кинкелин К., Пай В.Е., Рено Л., Дуглас М.Э., Ремус Д., Черепанов П., Коста А., Диффли Дж.Ф. (июнь 2017 г.). «Cdt1 стабилизирует открытое кольцо MCM для загрузки геликазы» . Природные коммуникации . 8 : 15720. Бибкод : 2017NatCo...815720F . дои : 10.1038/ncomms15720 . ПМК 5490006 . ПМИД 28643783 .
- ^ Тикау С., Фридман Л.Дж., Чампаса К., Корреа И.Р., Геллес Дж., Белл С.П. (март 2017 г.). «Механизм и время закрытия кольца Mcm2-7 во время лицензирования источника репликации ДНК» . Структурная и молекулярная биология природы . 24 (3): 309–315. дои : 10.1038/nsmb.3375 . ПМК 5336523 . ПМИД 28191892 .
- ^ Нишитани Х., Лигеро З., Нишимото Т., медсестра П. (апрель 2000 г.). «Белок Cdt1 необходим для лицензирования ДНК для репликации в делящихся дрожжах». Природа . 404 (6778): 625–8. Бибкод : 2000Natur.404..625N . дои : 10.1038/35007110 . ПМИД 10766248 . S2CID 205005540 .
- ^ Юань З., Риера А., Бай Л., Сунь Дж., Нанди С., Спанос С., Чен З.А., Барбон М., Раппсилбер Дж. , Стиллман Б., Спек С., Ли Х (март 2017 г.). «Структурная основа репликативной загрузки геликазы Mcm2-7 с помощью ORC-Cdc6 и Cdt1» . Структурная и молекулярная биология природы . 24 (3): 316–324. дои : 10.1038/nsmb.3372 . ПМК 5503505 . ПМИД 28191893 .
- ^ Вольшлегель Дж.А., Дуайер Б.Т., Дхар С.К., Цветич С., Уолтер Дж.К., Датта А. (декабрь 2000 г.). «Ингибирование репликации эукариотической ДНК путем связывания геминина с Cdt1». Наука . 290 (5500): 2309–12. Бибкод : 2000Sci...290.2309W . дои : 10.1126/science.290.5500.2309 . ПМИД 11125146 .
- ^ Мэн Г.Т., Синха П., Тай Б.К. (март 1984 г.). «Мутанты S. cerevisiae с дефектом содержания минихромосом» . Генетика . 106 (3): 365–85. дои : 10.1093/генетика/106.3.365 . ПМЦ 1224244 . ПМИД 6323245 .
- ^ Хуа XH, Ньюпорт Дж (январь 1998 г.). «Идентификация стадии предварительной инициации репликации ДНК, которая не зависит от комплекса распознавания происхождения и cdc6, но зависит от cdk2» . Журнал клеточной биологии . 140 (2): 271–81. дои : 10.1083/jcb.140.2.271 . ПМК 2132576 . ПМИД 9442103 .
- ^ Роулз А., Тада С., Блоу Джей-Джей (июнь 1999 г.). «Изменения в ассоциации комплекса распознавания происхождения Xenopus с хроматином при лицензировании источников репликации» . Журнал клеточной науки . 112 (12): 2011–8. дои : 10.1242/jcs.112.12.2011 . ПМЦ 3605702 . ПМИД 10341218 .
- ^ Перейти обратно: а б с д Ли Н, Чжай Ю, Чжан Ю, Ли В, Ян М, Лэй Дж, Тай БК, Гао Н (август 2015 г.). «Структура эукариотического комплекса MCM на расстоянии 3,8 Å». Природа . 524 (7564): 186–91. Бибкод : 2015Natur.524..186L . дои : 10.1038/nature14685 . ПМИД 26222030 . S2CID 4468690 .
- ^ Перейти обратно: а б Ногучи И., Юань З., Бай Л., Шнайдер С., Чжао Г., Стиллман Б., Спек С., Ли Х. (ноябрь 2017 г.). «Крио-ЭМ-структура двойного гексамера Mcm2-7 на ДНК предполагает модель экструзии ДНК с отстающей цепью» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (45): Е9529–Е9538. Бибкод : 2017PNAS..114E9529N . дои : 10.1073/pnas.1712537114 . ПМК 5692578 . ПМИД 29078375 .
- ^ Перейти обратно: а б Датта А., Белл СП (1997). «Инициация репликации ДНК в эукариотических клетках». Ежегодный обзор клеточной биологии и биологии развития . 13 : 293–332. дои : 10.1146/annurev.cellbio.13.1.293 . ПМИД 9442876 .
- ^ Перейти обратно: а б Лабиб К., Терсеро Дж.А., Диффли Дж.Ф. (июнь 2000 г.). «Непрерывная функция MCM2-7 необходима для развития вилки репликации ДНК». Наука . 288 (5471): 1643–7. Бибкод : 2000Sci...288.1643L . дои : 10.1126/science.288.5471.1643 . ПМИД 10834843 .
- ^ Швача А., Bell SP (ноябрь 2001 г.). «Взаимодействия между двумя каталитически различными подгруппами MCM необходимы для скоординированного гидролиза АТФ и репликации ДНК» . Молекулярная клетка . 8 (5): 1093–104. дои : 10.1016/S1097-2765(01)00389-6 . ПМИД 11741544 .
- ^ Бохман М.Л., Белл С.П., Швача А. (октябрь 2008 г.). «Субъединичная организация Mcm2-7 и неравная роль активных центров в гидролизе и жизнеспособности АТФ» . Молекулярная и клеточная биология . 28 (19): 5865–5873. дои : 10.1128/MCB.00161-08 . ПМК 2547011 . ПМИД 18662997 .
- ^ Хингорани М.М., Вашингтон М.Т., Мур К.С., Патель С.С. (май 1997 г.). «Механизм dTTPase ДНК-хеликазы T7 напоминает механизм изменения связывания F1-АТФазы» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (10): 5012–7. Бибкод : 1997PNAS...94.5012H . дои : 10.1073/pnas.94.10.5012 . ПМК 24622 . ПМИД 9144181 .
- ^ Ян Х., Торговец А.М., Тай Б.К. (ноябрь 1993 г.). «Регулируемая клеточным циклом ядерная локализация MCM2 и MCM3, которые необходимы для инициации синтеза ДНК в точках начала хромосомной репликации у дрожжей» . Гены и развитие . 7 (11): 2149–60. дои : 10.1101/gad.7.11.2149 . ПМИД 8224843 .
- ^ Янг М.Р., Сузуки К., Ян Х., Гибсон С., Тай Б.К. (октябрь 1997 г.). «Ядерное накопление Saccharomyces cerevisiae Mcm3 зависит от последовательности его ядерной локализации» . Гены в клетки . 2 (10): 631–43. дои : 10.1046/j.1365-2443.1997.1510349.x . ПМИД 9427284 .
- ^ Лабиб К., Диффли Дж. Ф., Кирси С. Е. (ноябрь 1999 г.). «Цикины G1-фазы и B-типа исключают фактор репликации ДНК Mcm4 из ядра». Природная клеточная биология . 1 (7): 415–22. дои : 10.1038/15649 . ПМИД 10559985 . S2CID 23407351 .
- ^ Лей М., Тай Б.К. (апрель 2001 г.). «Инициирование синтеза ДНК: от рекрутирования до активации комплекса MCM». Журнал клеточной науки . 114 (Часть 8): 1447–54. дои : 10.1242/jcs.114.8.1447 . ПМИД 11282021 .
- ^ Абид Али Ф., Дуглас М.Э., Локк Дж., Пай В.Е., Нанс А., Диффли Дж.Ф., Коста А. (декабрь 2017 г.). «Крио-ЭМ структура лицензированного источника репликации ДНК» . Природные коммуникации . 8 (1): 2241. Бибкод : 2017NatCo...8.2241A . дои : 10.1038/s41467-017-02389-0 . ПМК 5740162 . ПМИД 29269875 .
- ^ Ли Дж., Донг Дж., Ван В., Ю Д., Фань Х., Хуэй Ю.К. и др. (январь 2023 г.). «Пререпликационный комплекс человека представляет собой открытый комплекс» (PDF) . Клетка . 186 (1): 98–111.e21. дои : 10.1016/j.cell.2022.12.008 . ПМИД 36608662 . S2CID 255442139 .
- ^ Перейти обратно: а б Грейве Дж.Ф., Миллер Т.К., Локк Дж., Мартино Ф., Хауэлл С., Шрайбер А. и др. (январь 2022 г.). «Структурный механизм селективного фосфорилирования двойных гексамеров MCM, нагруженных ДНК, с помощью Dbf4-зависимой киназы» . Структурная и молекулярная биология природы . 29 (1): 10–20. дои : 10.1038/s41594-021-00698-z . ПМЦ 8770131 . ПМИД 34963704 .
- ^ Перейти обратно: а б Салех А., Ногучи Ю., Арамайо Р., Иванова М.Е., Стивенс К.М., Монтойя А. и др. (май 2022 г.). «Структурная основа зависимого от киназы Cdc7-Dbf4 нацеливания и фосфорилирования двойного гексамера MCM2-7» . Природные коммуникации . 13 (1): 2915. Бибкод : 2022NatCo..13.2915S . дои : 10.1038/s41467-022-30576-1 . ПМЦ 9133112 . ПМИД 35614055 .
- ^ Перейти обратно: а б с д Ченг Дж, Ли Н, Хо Ю, Данг С, Тай БК, Гао Н, Чжай Ю (март 2022 г.). «Структурное понимание активации двойного гексамера MCM киназой Dbf4-Cdc7» . Природные коммуникации . 13 (1): 1396. Бибкод : 2022NatCo..13.1396C . дои : 10.1038/s41467-022-29070-5 . ПМЦ 8927117 . ПМИД 35296675 .
- ^ Перейти обратно: а б Шу Ю.Дж., Стиллман Б. (январь 2010 г.). «Киназа Dbf4-Cdc7 способствует S-фазе, ослабляя ингибирующую активность Mcm4» . Природа . 463 (7277): 113–117. Бибкод : 2010Natur.463..113S . дои : 10.1038/nature08647 . ПМК 2805463 . ПМИД 20054399 .
- ^ Терсеро Дж. А., Лабиб К., Диффли Дж. Ф. (май 2000 г.). «Синтез ДНК на отдельных вилках репликации требует наличия существенного фактора инициации Cdc45p» . Журнал ЭМБО . 19 (9): 2082–93. дои : 10.1093/emboj/19.9.2082 . ПМК 305696 . ПМИД 10790374 .
- ^ Хеннесси К.М., Ли А., Чен Э., Ботштейн Д. (июнь 1991 г.). «Группа взаимодействующих генов репликации ДНК дрожжей» . Гены и развитие . 5 (6): 958–69. дои : 10.1101/gad.5.6.958 . ПМИД 2044962 .
- ^ Перейти обратно: а б Апарисио О.М., Вайнштейн Д.М., Белл С.П. (октябрь 1997 г.). «Компоненты и динамика комплексов репликации ДНК у S. cerevisiae: перераспределение белков MCM и Cdc45p во время S-фазы» . Клетка . 91 (1): 59–69. дои : 10.1016/S0092-8674(01)80009-X . ПМИД 9335335 . S2CID 10353164 .
- ^ Перейти обратно: а б Мимура С., Масуда Т., Мацуи Т., Такисава Х. (июнь 2000 г.). «Центральная роль cdc45 в создании комплекса инициации репликации ДНК в экстрактах яиц Xenopus» . Гены в клетки . 5 (6): 439–52. дои : 10.1046/j.1365-2443.2000.00340.x . PMID 10886370 .
- ^ Апарисио О.М., Стаут А.М., Белл С.П. (август 1999 г.). «Дифференциальная сборка Cdc45p и ДНК-полимераз в ранних и поздних точках начала репликации ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (16): 9130–5. Бибкод : 1999PNAS...96.9130A . дои : 10.1073/pnas.96.16.9130 . ЧВК 17744 . ПМИД 10430907 .
- ^ Цзоу Л., Стиллман Б. (май 2000 г.). «Сборка комплекса, содержащего Cdc45p, репликационный белок A и Mcm2p в точках начала репликации, контролируемых циклин-зависимыми киназами S-фазы и киназой Cdc7p-Dbf4p» . Молекулярная и клеточная биология . 20 (9): 3086–96. дои : 10.1128/mcb.20.9.3086-3096.2000 . ПМК 85601 . ПМИД 10757793 .
- ^ Перейти обратно: а б Уолтер Дж., Ньюпорт Дж. (апрель 2000 г.). «Инициация репликации эукариотической ДНК: раскручивание начала и последовательная ассоциация хроматина Cdc45, RPA и ДНК-полимеразы альфа» . Молекулярная клетка . 5 (4): 617–27. дои : 10.1016/S1097-2765(00)80241-5 . ПМИД 10882098 .
- ^ Перейти обратно: а б с Гамбус А., Джонс Р.К., Санчес-Диас А., Канемаки М., ван Дёрсен Ф., Эдмондсон Р.Д., Лабиб К. (апрель 2006 г.). «GINS поддерживает ассоциацию Cdc45 с MCM в комплексах реплисомной прогрессии на вилках репликации эукариотической ДНК». Природная клеточная биология . 8 (4): 358–66. дои : 10.1038/ncb1382 . ПМИД 16531994 . S2CID 21543095 .
- ^ Перейти обратно: а б Канемаки М., Санчес-Диас А., Гамбус А., Лабиб К. (июнь 2003 г.). «Функциональная протеомная идентификация белков репликации ДНК путем индуцированного протеолиза in vivo». Природа . 423 (6941): 720–4. Бибкод : 2003Natur.423..720K . дои : 10.1038/nature01692 . ПМИД 12768207 . S2CID 4345091 .
- ^ Макарова К.С., Вольф Ю.И., Мехедов С.Л., Миркин Б.Г., Кунин Е.В. (2005). «Предковые паралоги и псевдопаралоги и их роль в возникновении эукариотической клетки» . Исследования нуклеиновых кислот . 33 (14): 4626–38. дои : 10.1093/nar/gki775 . ПМЦ 1187821 . ПМИД 16106042 .
- ^ Перейти обратно: а б с Мойер С.Е., Льюис П.В., Ботчан М.Р. (июль 2006 г.). «Выделение комплекса Cdc45/Mcm2-7/GINS (CMG), кандидата на роль геликазы вилки репликации эукариотической ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (27): 10236–10241. Бибкод : 2006PNAS..10310236M . дои : 10.1073/pnas.0602400103 . ПМЦ 1482467 . ПМИД 16798881 .
- ^ Бохман М.Л., Швача А. (июль 2008 г.). «Комплекс Mcm2-7 обладает геликазной активностью in vitro» . Молекулярная клетка . 31 (2): 287–93. doi : 10.1016/j.molcel.2008.05.020 . ПМИД 18657510 .
- ^ Перейти обратно: а б Коста А., Ильвес И., Тамберг Н., Петоевич Т., Ногалес Е., Ботчан М.Р., Бергер Дж.М. (апрель 2011 г.). «Структурная основа активации хеликазы MCM2-7 с помощью GINS и Cdc45» . Структурная и молекулярная биология природы . 18 (4): 471–7. дои : 10.1038/nsmb.2004 . ПМК 4184033 . ПМИД 21378962 .
- ^ Юань З., Бай Л., Сунь Дж., Джорджеску Р., Лю Дж., О'Доннелл М.Э., Ли Х. (март 2016 г.). «Структура эукариотической репликативной геликазы CMG предполагает движение качалки для транслокации» . Структурная и молекулярная биология природы . 23 (3): 217–24. дои : 10.1038/nsmb.3170 . ПМЦ 4812828 . ПМИД 26854665 .
- ^ Хомсли Л., Лей М., Кавасаки Ю., Сойер С., Кристенсен Т., Тай Б.К. (апрель 2000 г.). «Mcm10 и комплекс MCM2-7 взаимодействуют, инициируя синтез ДНК и высвобождая факторы репликации из источников» . Гены и развитие . 14 (8): 913–26. дои : 10.1101/gad.14.8.913 . ПМК 316538 . ПМИД 10783164 .
- ^ Кристенсен Т.В., Тай Б.К. (июнь 2003 г.). «Дрозофила MCM10 взаимодействует с членами пререпликационного комплекса и необходима для правильной конденсации хромосом» . Молекулярная биология клетки . 14 (6): 2206–15. дои : 10.1091/mbc.e02-11-0706 . ЧВК 194871 . ПМИД 12808023 .
- ^ Ли С., Лячко И., Бутен Р., Кельман З., Тай Б.К. (январь 2010 г.). «Альтернативные механизмы координации альфа-полимеразы и геликазы MCM» . Молекулярная и клеточная биология . 30 (2): 423–35. дои : 10.1128/MCB.01240-09 . ПМЦ 2798462 . ПМИД 19917723 .
- ^ ван Дёрсен Ф., Сенгупта С., Де Пикколи Г., Санчес-Диас А., Лабиб К. (май 2012 г.). «Mcm10 связывается с загруженной ДНК-хеликазой в начале репликации и определяет новый этап ее активации» . Журнал ЭМБО . 31 (9): 2195–206. дои : 10.1038/emboj.2012.69 . ПМЦ 3343467 . ПМИД 22433841 .
- ^ Масаи Х., Сато Н., Такеда Т., Араи К. (декабрь 1999 г.). «Киназный комплекс CDC7 как молекулярный переключатель репликации ДНК». Границы бионауки . 4 (1–3): Д834-40. дои : 10.2741/масаи . ПМИД 10577390 .
- ^ Мимура С., Такисава Х. (октябрь 1998 г.). «Xenopus Cdc45-зависимая загрузка ДНК-полимеразы альфа на хроматин под контролем S-фазы Cdk» . Журнал ЭМБО . 17 (19): 5699–707. дои : 10.1093/emboj/17.19.5699 . ПМЦ 1170898 . ПМИД 9755170 .
- ^ Цзоу Л., Стиллман Б. (апрель 1998 г.). «Образование преинициативного комплекса путем S-фазного циклина CDK-зависимой загрузки Cdc45p на хроматин». Наука . 280 (5363): 593–6. Бибкод : 1998Sci...280..593Z . дои : 10.1126/science.280.5363.593 . ПМИД 9554851 .
- ^ Цзян В., Уэллс, Нью-Джерси, Хантер Т. (май 1999 г.). «Многоступенчатая регуляция репликации ДНК путем фосфорилирования Cdk HsCdc6» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (11): 6193–8. Бибкод : 1999PNAS...96.6193J . дои : 10.1073/pnas.96.11.6193 . ПМК 26858 . ПМИД 10339564 .
- ^ Перейти обратно: а б Цзян В., Макдональд Д., Хоуп ТиДжей, Хантер Т. (октябрь 1999 г.). «Комплекс протеинкиназы Cdc7-Dbf4 млекопитающих необходим для инициации репликации ДНК» . Журнал ЭМБО . 18 (20): 5703–5713. дои : 10.1093/emboj/18.20.5703 . ПМЦ 1171637 . ПМИД 10523313 .
- ^ Кумагай Х., Сато Н., Ямада М., Махони Д., Сегецци В., Лис Э. и др. (июль 1999 г.). «Новый белок, регулирующий рост и клеточный цикл, ASK, активирует человеческую киназу, родственную Cdc7, и необходим для перехода G1/S в клетках млекопитающих» . Молекулярная и клеточная биология . 19 (7): 5083–5095. дои : 10.1128/MCB.19.7.5083 . ПМЦ 84351 . ПМИД 10373557 .
- ^ Фрэнсис Л.И., Рэнделл Дж.К., Такара Т.Дж., Учима Л., Белл С.П. (март 2009 г.). «Включение в пререпликативный комплекс активирует хеликазу Mcm2-7 для фосфорилирования Cdc7-Dbf4» . Гены и развитие . 23 (5): 643–654. дои : 10.1101/gad.1759609 . ПМЦ 2658526 . ПМИД 19270162 .
- ^ Сунь Дж., Фернандес-Сид А., Риера А., Тогнетти С., Юань З., Стиллман Б. и др. (октябрь 2014 г.). «Структурные и механистические сведения о сборке и функционировании двойного гексамера Mcm2-7» . Гены и развитие . 28 (20): 2291–2303. дои : 10.1101/gad.242313.114 . ПМК 4201289 . ПМИД 25319829 .
- ^ Лей М., Кавасаки Ю., Янг М.Р., Кихара М., Сугино А., Тай Б.К. (декабрь 1997 г.). «Mcm2 является мишенью регуляции Cdc7-Dbf4 во время инициации синтеза ДНК» . Гены и развитие . 11 (24): 3365–3374. дои : 10.1101/gad.11.24.3365 . ПМК 316824 . ПМИД 9407029 .
- ^ Камимура Ю., Так Ю.С., Сугино А., Араки Х. (апрель 2001 г.). «Sld3, который взаимодействует с Cdc45 (Sld4), участвует в репликации хромосомной ДНК у Saccharomyces cerevisiae» . Журнал ЭМБО . 20 (8): 2097–107. дои : 10.1093/emboj/20.8.2097 . ПМК 125422 . ПМИД 11296242 .
- ^ Канемаки М., Лабиб К. (апрель 2006 г.). «Различные роли Sld3 и GINS во время создания и развития вилок репликации эукариотической ДНК» . Журнал ЭМБО . 25 (8): 1753–63. дои : 10.1038/sj.emboj.7601063 . ПМК 1440835 . ПМИД 16601689 .
- ^ Масумото Х., Сугино А., Араки Х. (апрель 2000 г.). «Dpb11 контролирует связь между ДНК-полимеразами альфа и эпсилон и автономно реплицирующейся областью последовательности почкующихся дрожжей» . Молекулярная и клеточная биология . 20 (8): 2809–17. дои : 10.1128/mcb.20.8.2809-2817.2000 . ПМК 85497 . ПМИД 10733584 .
- ^ Араки Х., Лим С.Х., Фонгдара А., Сугино А. (декабрь 1995 г.). «Dpb11, который взаимодействует с ДНК-полимеразой II (эпсилон) в Saccharomyces cerevisiae, играет двойную роль в развитии S-фазы и в контрольной точке клеточного цикла» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (25): 11791–5. дои : 10.1073/pnas.92.25.11791 . ПМК 40488 . ПМИД 8524850 .
- ^ Гловер Дж. Н., Уильямс Р. С., Ли М. С. (ноябрь 2004 г.). «Взаимодействие между повторами BRCT и фосфопротеинами: запутано пополам». Тенденции биохимических наук . 29 (11): 579–85. дои : 10.1016/j.tibs.2004.09.010 . ПМИД 15501676 .
- ^ Масумото Х., Мурамацу С., Камимура Ю., Араки Х. (февраль 2002 г.). «S-Cdk-зависимое фосфорилирование Sld2, необходимое для репликации хромосомной ДНК у почкующихся дрожжей». Природа . 415 (6872): 651–5. Бибкод : 2002Natur.415..651M . дои : 10.1038/nature713 . ПМИД 11807498 . S2CID 4300863 .
- ^ Такаяма Ю., Камимура Ю., Окава М., Мурамацу С., Сугино А., Араки Х. (май 2003 г.). «GINS, новый мультибелковый комплекс, необходимый для репликации хромосомной ДНК у почкующихся дрожжей» . Гены и развитие . 17 (9): 1153–65. дои : 10.1101/gad.1065903 . ПМК 196052 . ПМИД 12730134 .
- ^ Лабиб К., Гамбус А. (июнь 2007 г.). «Ключевая роль комплекса GINS в вилках репликации ДНК». Тенденции в клеточной биологии . 17 (6): 271–8. дои : 10.1016/j.tcb.2007.04.002 . ПМИД 17467990 .
- ^ Мурамацу С., Хираи К., Так Ю.С., Камимура Ю., Араки Х. (март 2010 г.). «CDK-зависимое образование комплекса между белками репликации Dpb11, Sld2, Pol (эпсилон) и GINS у почкующихся дрожжей» . Гены и развитие . 24 (6): 602–12. дои : 10.1101/gad.1883410 . ПМЦ 2841337 . ПМИД 20231317 .
- ^ Леман И.Р., Бессман М.Дж., Симмс Э.С., Корнберг А. (июль 1958 г.). «Ферментативный синтез дезоксирибонуклеиновой кислоты. I. Получение субстратов и частичная очистка фермента из кишечной палочки» . Журнал биологической химии . 233 (1): 163–70. дои : 10.1016/S0021-9258(19)68048-8 . ПМИД 13563462 .
- ^ Алани Э., Трешер Р., Гриффит Дж.Д., Колоднер Р.Д. (сентябрь 1992 г.). «Характеристика свойств стимуляции ДНК-связывания и обмена цепей y-RPA, дрожжевого белка, связывающего одноцепочечную ДНК». Журнал молекулярной биологии . 227 (1): 54–71. дои : 10.1016/0022-2836(92)90681-9 . ПМИД 1522601 .
- ^ Гулиан М., Ричардс С.Х., Херд СиДжей, Бигсби Б.М. (октябрь 1990 г.). «Прерывистый синтез ДНК очищенными белками млекопитающих» . Журнал биологической химии . 265 (30): 18461–71. дои : 10.1016/S0021-9258(17)44775-2 . ПМИД 2170412 .
- ^ Маккалок С.Д., Кункель Т.А. (январь 2008 г.). «Правильность синтеза ДНК с помощью эукариотических полимераз репликативного и транслейкционного синтеза» . Клеточные исследования . 18 (1): 148–61. дои : 10.1038/cr.2008.4 . ПМЦ 3639319 . ПМИД 18166979 .
- ^ Перселл З.Ф., Исоз И., Лундстрем Э.Б., Йоханссон Э., Кункель Т.А. (июль 2007 г.). «ДНК-полимераза дрожжей эпсилон участвует в репликации ДНК ведущей цепи» . Наука . 317 (5834): 127–30. Бибкод : 2007Sci...317..127P . дои : 10.1126/science.1144067 . ПМК 2233713 . ПМИД 17615360 .
- ^ Перейти обратно: а б с д и Уотсон Дж., Бейкер Т., Белл С., Ганн А., Левин М., Лосик Р., Молекулярная биология гена, 6-е издание , Pearson Education Inc, 2008. ISBN 080539592X
- ^ Вага С., Бауэр Г., Стиллман Б. (апрель 1994 г.). «Восстановление полной репликации ДНК SV40 с помощью очищенных факторов репликации» . Журнал биологической химии . 269 (14): 10923–34. дои : 10.1016/S0021-9258(17)34146-7 . ПМИД 8144677 .
- ^ Гарг П., Стит К.М., Сабури Н., Йоханссон Э., Бургерс П.М. (ноябрь 2004 г.). «Бездействие ДНК-полимеразы дельта сохраняет лигируемый разрыв во время репликации отстающей цепи ДНК» . Гены и развитие . 18 (22): 2764–73. дои : 10.1101/gad.1252304 . ПМК 528896 . ПМИД 15520275 .
- ^ Burgers PM (февраль 2009 г.). «Динамика полимеразы на репликационной вилке эукариотической ДНК» . Журнал биологической химии . 284 (7): 4041–5. дои : 10.1074/jbc.R800062200 . ПМК 2640984 . ПМИД 18835809 .
- ^ Джин Ю.Х., Айягари Р., Резник М.А., Горденин Д.А., Бургерс П.М. (январь 2003 г.). «Созревание фрагмента Оказаки в дрожжах. II. Сотрудничество между полимеразной и 3'-5'-экзонуклеазной активностью Pol delta при создании лигируемого разрыва» . Журнал биологической химии . 278 (3): 1626–33. дои : 10.1074/jbc.M209803200 . ПМИД 12424237 .
- ^ Сого Дж. М., Лопес М., Фойани М. (июль 2002 г.). «Реверс вилки и накопление оцДНК при остановке репликационных вилок из-за дефектов контрольных точек». Наука . 297 (5581): 599–602. Бибкод : 2002Sci...297..599S . дои : 10.1126/science.1074023 . ПМИД 12142537 . S2CID 33502697 .
- ^ Моррисон А., Араки Х., Кларк А.Б., Хаматаке Р.К., Сугино А. (сентябрь 1990 г.). «Третья незаменимая ДНК-полимераза S. cerevisiae». Клетка . 62 (6): 1143–51. дои : 10.1016/0092-8674(90)90391-Q . ПМИД 2169349 . S2CID 29672985 .
- ^ Вага С., Стиллман Б. (май 1994 г.). «Анатомия репликационной вилки ДНК, выявленная путем восстановления репликации ДНК SV40 in vitro». Природа . 369 (6477): 207–12. Бибкод : 1994Natur.369..207W . дои : 10.1038/369207a0 . ПМИД 7910375 . S2CID 4351628 .
- ^ Цуримото Т., Стиллман Б. (январь 1991 г.). «Факторы репликации, необходимые для репликации ДНК SV40 in vitro. II. Переключение альфа- и дельта-полимеразы ДНК во время инициации синтеза ведущей и отстающей цепи» . Журнал биологической химии . 266 (3): 1961–8. дои : 10.1016/S0021-9258(18)52386-3 . ПМИД 1671046 .
- ^ Барри Э.Р., Белл С.Д. (декабрь 2006 г.). «Репликация ДНК у архей» . Обзоры микробиологии и молекулярной биологии . 70 (4): 876–87. дои : 10.1128/MMBR.00029-06 . ПМК 1698513 . ПМИД 17158702 .
- ^ Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2003). Биохимия . Гейдельберг/Берлин: Springer.
- ^ Перейти обратно: а б Джонсон Р.Э., Классен Р., Пракаш Л., Пракаш С. (июль 2015 г.). «Основная роль ДНК-полимеразы δ в репликации как ведущих, так и отстающих цепей ДНК» . Молекулярная клетка . 59 (2): 163–175. doi : 10.1016/j.molcel.2015.05.038 . ПМЦ 4517859 . ПМИД 26145172 .
- ^ Бюн Т.С., Пачек М., Йи MC, Уолтер Дж.К., Цимприч К.А. (май 2005 г.). «Функциональное развязывание активности MCM-хеликазы и ДНК-полимеразы активирует ATR-зависимую контрольную точку» . Гены и развитие . 19 (9): 1040–52. дои : 10.1101/gad.1301205 . ПМЦ 1091739 . ПМИД 15833913 .
- ^ Перейти обратно: а б Ремус Д., Бейрон Ф., Толан Дж., Гриффит Дж.Д., Моррис Э.П., Диффли Дж.Ф. (ноябрь 2009 г.). «Согласованная загрузка двойных гексамеров Mcm2-7 вокруг ДНК во время лицензирования происхождения репликации ДНК» . Клетка . 139 (4): 719–30. дои : 10.1016/j.cell.2009.10.015 . ПМК 2804858 . ПМИД 19896182 .
- ^ Эврин С., Кларк П., Зех Дж., Лурц Р., Сан Дж., Уле С., Ли Х., Стиллман Б., Спек С. (декабрь 2009 г.). «Двойной гексамерный комплекс MCM2-7 загружается в исходную ДНК во время лицензирования репликации эукариотической ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (48): 20240–5. Бибкод : 2009PNAS..10620240E . дои : 10.1073/pnas.0911500106 . ПМЦ 2787165 . ПМИД 19910535 .
- ^ Лабиб К. (июнь 2010 г.). «Как Cdc7 и циклин-зависимые киназы запускают инициацию репликации хромосом в эукариотических клетках?» . Гены и развитие . 24 (12): 1208–19. дои : 10.1101/gad.1933010 . ПМЦ 2885657 . ПМИД 20551170 .
- ^ Пачек М., Уолтер Дж. К. (сентябрь 2004 г.). «Необходимость MCM7 и Cdc45 при раскручивании хромосом во время репликации эукариотической ДНК» . Журнал ЭМБО . 23 (18): 3667–76. дои : 10.1038/sj.emboj.7600369 . ПМК 517609 . ПМИД 15329670 .
- ^ Яо, Нью-Йорк, О'Доннелл М. (июнь 2010 г.). «Снимок: Реплисома» . Клетка . 141 (6): 1088–1088.e1. дои : 10.1016/j.cell.2010.05.042 . ПМК 4007198 . ПМИД 20550941 .
- ^ Бермудес В.П., Фарина А., Таппин И., Гурвиц Дж. (март 2010 г.). «Влияние фактора установления сплоченности человека Ctf4/AND-1 на репликацию ДНК» . Журнал биологической химии . 285 (13): 9493–505. дои : 10.1074/jbc.M109.093609 . ПМК 2843200 . ПМИД 20089864 .
- ^ Чжу В., Укомаду С., Джа С., Сенга Т., Дхар С.К., Вольшлегель Дж.А., Натт Л.К., Корнблут С., Дутта А. (сентябрь 2007 г.). «Mcm10 и And-1/CTF4 рекрутируют ДНК-полимеразу альфа в хроматин для инициации репликации ДНК» . Гены и развитие . 21 (18): 2288–99. дои : 10.1101/gad.1585607 . ЧВК 1973143 . ПМИД 17761813 .
- ^ Лу Х., Комата М., Като Ю., Гуан З., Рейс К.С., Бадд М., Сирахигэ К., Кэмпбелл Дж.Л. (октябрь 2008 г.). «Mrc1 и ДНК-полимераза эпсилон функционируют вместе, связывая репликацию ДНК и контрольную точку S-фазы» . Молекулярная клетка . 32 (1): 106–17. doi : 10.1016/j.molcel.2008.08.020 . ПМЦ 2699584 . ПМИД 18851837 .
- ^ Петерманн Э., Хелледей Т., Калдекотт К.В. (июнь 2008 г.). «Класпин способствует нормальной скорости репликации в клетках человека» . Молекулярная биология клетки . 19 (6): 2373–8. дои : 10.1091/mbc.E07-10-1035 . ПМК 2397295 . ПМИД 18353973 .
- ^ Браво Р., Фрэнк Р., Бланделл П.А., Макдональд-Браво Х. (1987). «Циклин/PCNA — вспомогательный белок ДНК-полимеразы-дельта». Природа . 326 (6112): 515–7. Бибкод : 1987Natur.326..515B . дои : 10.1038/326515a0 . ПМИД 2882423 . S2CID 4344147 .
- ^ Прелич Г., Тан С.К., Костура М., Мэтьюз М.Б., Со А.Г., Дауни К.М., Стиллман Б. (1987). «Функциональная идентичность ядерного антигена пролиферирующей клетки и вспомогательного белка ДНК-полимеразы-дельта». Природа . 326 (6112): 517–20. Бибкод : 1987Natur.326..517P . дои : 10.1038/326517a0 . ПМИД 2882424 . S2CID 4345171 .
- ^ Молдаванин Г.Л., Пфандер Б., Йенч С. (май 2007 г.). «PCNA, маэстро репликационной вилки» . Клетка . 129 (4): 665–79. дои : 10.1016/j.cell.2007.05.003 . ПМИД 17512402 . S2CID 3547069 .
- ^ Кай Дж., Яо Н., Гиббс Э., Финкельштейн Дж., Филлипс Б., О'Доннелл М., Гурвиц Дж. (сентябрь 1998 г.). «Гидролиз АТФ, катализируемый фактором репликации C человека, требует участия нескольких субъединиц» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (20): 11607–12. Бибкод : 1998PNAS...9511607C . дои : 10.1073/pnas.95.20.11607 . ПМК 21688 . ПМИД 9751713 .
- ^ Цуримото Т., Стиллман Б. (январь 1991 г.). «Факторы репликации, необходимые для репликации ДНК SV40 in vitro. I. Структурно-специфическое распознавание соединения праймер-матрица эукариотическими ДНК-полимеразами и их вспомогательными белками» . Журнал биологической химии . 266 (3): 1950–60. дои : 10.1016/S0021-9258(18)52385-1 . ПМИД 1671045 .
- ^ Подуст Л.М., Подуст В.Н., Сого Ю.М., Хюбшер У (июнь 1995 г.). «Вспомогательные белки ДНК-полимеразы млекопитающих: анализ загрузки ядерного антигена пролиферирующих клеток, катализируемой фактором репликации C, на кольцевую двухцепочечную ДНК» . Молекулярная и клеточная биология . 15 (6): 3072–81. дои : 10.1128/MCB.15.6.3072 . ПМК 230538 . ПМИД 7760803 .
- ^ Чжан Г., Гиббс Э., Кельман З., О'Доннелл М., Гурвиц Дж. (март 1999 г.). «Исследования взаимодействия между фактором репликации человека C и ядерным антигеном пролиферирующих клеток человека» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (5): 1869–74. Бибкод : 1999PNAS...96.1869Z . дои : 10.1073/pnas.96.5.1869 . ПМЦ 26703 . ПМИД 10051561 .
- ^ Перейти обратно: а б Яо Нью-Йорк, Джонсон А., Боуман Г.Д., Куриян Дж., О'Доннелл М. (июнь 2006 г.). «Механизм открытия зажима ядерного антигена пролиферирующих клеток фактором репликации C» . Журнал биологической химии . 281 (25): 17528–39. дои : 10.1074/jbc.M601273200 . ПМИД 16608854 .
- ^ Перейти обратно: а б с Сабатинос, SA (2010). «Восстановление остановившейся вилки репликации» . Воспитательное природное образование . 3 (9): 31.
- ^ Перейти обратно: а б Ротштейн Р., Мишель Б., Ганглофф С. (январь 2000 г.). «Приостановка репликационной вилки и рекомбинация или «дай мне передышку» » . Гены и развитие . 14 (1): 1–10. дои : 10.1101/gad.14.1.1 . ПМИД 10640269 . S2CID 40751623 .
- ^ Бастия Д., Ззаман С., Крингс Г., Саксена М., Пэн Х., Гринберг М.М. (сентябрь 2008 г.). «Механизм терминации репликации, выявленный посредством Tus-опосредованного полярного ареста скользящей геликазы» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (35): 12831–6. Бибкод : 2008PNAS..10512831B . дои : 10.1073/pnas.0805898105 . ПМК 2529109 . ПМИД 18708526 .
- ^ Фабрики репликации, Павел Хозак, Питер Р. Кук, Тенденции в клеточной биологии, том 4, выпуск 2; Сайенсдирект , DOI
- ^ МакГарри Т.Дж., Киршнер М.В. (июнь 1998 г.). «Геминин, ингибитор репликации ДНК, разрушается во время митоза» . Клетка . 93 (6): 1043–53. doi : 10.1016/S0092-8674(00)81209-X . ПМИД 9635433 . S2CID 235485 .
- ^ де Кляйн А., Муйтьенс М., ван Ос Р., Верховен Ю., Смит Б., Карр А.М., Леманн А.Р., Хоймейкерс Дж.Х. (апрель 2000 г.). «Направленное нарушение гена контрольной точки клеточного цикла ATR приводит к ранней эмбриональной смертности у мышей» . Современная биология . 10 (8): 479–82. дои : 10.1016/S0960-9822(00)00447-4 . ПМИД 10801416 . S2CID 15401622 .
- ^ Кортес Д., Гунтуку С., Цинь Дж., Элледж С.Дж. (ноябрь 2001 г.). «ATR и ATRIP: партнеры в сигнализации контрольно-пропускных пунктов». Наука . 294 (5547): 1713–6. Бибкод : 2001Sci...294.1713C . дои : 10.1126/science.1065521 . ПМИД 11721054 . S2CID 32810119 .
- ^ Дарт Д.А., Адамс К.Е., Акерман И., Лакин Н.Д. (апрель 2004 г.). «Привлечение киназы контрольной точки клеточного цикла ATR к хроматину во время S-фазы» . Журнал биологической химии . 279 (16): 16433–40. дои : 10.1074/jbc.M314212200 . ПМИД 14871897 .
- ^ Болл Х.Л., Майерс Дж.С., Кортес Д. (май 2005 г.). «Связывание ATRIP с одноцепочечной ДНК репликационного белка A способствует локализации ATR-ATRIP, но не является обязательным для фосфорилирования Chk1» . Молекулярная биология клетки . 16 (5): 2372–81. дои : 10.1091/mbc.E04-11-1006 . ПМЦ 1087242 . ПМИД 15743907 .
- ^ Бермудес В.П., Линдси-Больц Л.А., Чезаре А.Дж., Манива Ю., Гриффит Дж.Д., Гурвиц Дж., Санкар А. (февраль 2003 г.). «Загрузка комплекса контрольных точек человека 9-1-1 на ДНК с помощью комплекса загрузчика контрольных точек hRad17-фактора репликации C in vitro» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (4): 1633–8. Бибкод : 2003PNAS..100.1633B . дои : 10.1073/pnas.0437927100 . ПМК 149884 . ПМИД 12578958 .
- ^ Зоу Л., Кортес Д., Элледж С.Дж. (январь 2002 г.). «Регуляция выбора субстрата ATR путем Rad17-зависимой загрузки комплексов Rad9 на хроматин» . Гены и развитие . 16 (2): 198–208. дои : 10.1101/gad.950302 . ПМК 155323 . ПМИД 11799063 .
- ^ Кумагай А., Данфи В.Г. (октябрь 2000 г.). «Класпин, новый белок, необходимый для активации Chk1 во время ответа контрольной точки репликации ДНК в экстрактах яиц Xenopus» . Молекулярная клетка . 6 (4): 839–49. дои : 10.1016/S1097-2765(05)00092-4 . ПМИД 11090622 .
- ^ Кумагай А., Данфи В.Г. (февраль 2003 г.). «Повторяющиеся фосфопептидные мотивы в Claspin опосредуют регулируемое связывание Chk1» (PDF) . Природная клеточная биология . 5 (2): 161–5. дои : 10.1038/ncb921 . ПМИД 12545175 . S2CID 29331203 .
- ^ Мяо Х., Зайлер Дж.А., Бурханс В.К. (февраль 2003 г.). «Регуляция начала репликации клеток и вируса SV40 с помощью Chk1-зависимых внутренних и контрольных точек, индуцированных УФ-излучением» . Журнал биологической химии . 278 (6): 4295–304. дои : 10.1074/jbc.M204264200 . ПМИД 12424256 .
- ^ Бар-Зив Р., Войчек Ю., Баркай Н. (сентябрь 2016 г.). «Динамика хроматина во время репликации ДНК» . Геномные исследования . 26 (9): 1245–56. дои : 10.1101/гр.201244.115 . ПМК 5052047 . ПМИД 27225843 .
- ^ Люгер К., Мэдер А.В., Ричмонд Р.К., Сарджент Д.Ф., Ричмонд Т.Дж. (сентябрь 1997 г.). «Кристаллическая структура ядра нуклеосомы при разрешении 2,8 А». Природа . 389 (6648): 251–60. Бибкод : 1997Natur.389..251L . дои : 10.1038/38444 . ПМИД 9305837 . S2CID 4328827 .
- ^ Виттмейер Дж., Джосс Л., Формоза Т. (июль 1999 г.). «Spt16 и Pob3 Saccharomyces cerevisiae образуют важный, обильный гетеродимер, который является ядерным, связанным с хроматином и очищается совместно с ДНК-полимеразой альфа». Биохимия . 38 (28): 8961–71. дои : 10.1021/bi982851d . ПМИД 10413469 .
- ^ Виттмейер Дж., Формоза Т. (июль 1997 г.). «Каталитическая альфа-субъединица ДНК-полимеразы Saccharomyces cerevisiae взаимодействует с Cdc68/Spt16 и с Pob3, белком, подобным HMG1-подобному белку» . Молекулярная и клеточная биология . 17 (7): 4178–90. дои : 10.1128/MCB.17.7.4178 . ПМК 232271 . ПМИД 9199353 .
- ^ Синь Х, Такахата С, Бланксма М, Маккалоу Л, Стиллман DJ, Формоза Т (август 2009 г.). «yFACT индуцирует глобальную доступность нуклеосомной ДНК без смещения H2A-H2B» . Молекулярная клетка . 35 (3): 365–76. doi : 10.1016/j.molcel.2009.06.024 . ПМК 2748400 . ПМИД 19683499 .
- ^ Грот А., Корпет А., Кук А.Дж., Рош Д., Бартек Дж., Лукас Дж., Альмузни Дж. (декабрь 2007 г.). «Регуляция развития репликационной вилки посредством спроса и предложения гистонов». Наука . 318 (5858): 1928–31. Бибкод : 2007Sci...318.1928G . дои : 10.1126/science.1148992 . hdl : 1885/53159 . ПМИД 18096807 . S2CID 22350778 .
- ^ Даганзо С.М., Эрцбергер Дж.П., Лам В.М., Скордалакес Э., Чжан Р., Франко А.А., Брилл С.Дж., Адамс П.Д., Бергер Дж.М., Кауфман П.Д. (декабрь 2003 г.). «Структура и функция консервативного ядра белка отложения гистонов Asf1» . Современная биология . 13 (24): 2148–58. дои : 10.1016/j.cub.2003.11.027 . ПМИД 14680630 . S2CID 15164132 .
- ^ Войчек Ю., Бар-Зив Р., Баркай Н. (март 2016 г.). «Гомеостаз экспрессии во время репликации ДНК». Наука . 351 (6277): 1087–90. Бибкод : 2016Sci...351.1087V . дои : 10.1126/science.aad1162 . ПМИД 26941319 . S2CID 32751800 .
- ^ Стиллман Б. (май 1986 г.). «Сборка хроматина во время репликации ДНК SV40 in vitro». Клетка . 45 (4): 555–65. дои : 10.1016/0092-8674(86)90287-4 . ПМИД 3011272 . S2CID 21212160 .
- ^ Шибахара К., Стиллман Б. (февраль 1999 г.). «Репликационно-зависимая маркировка ДНК с помощью PCNA облегчает наследование хроматина, связанное с CAF-1» . Клетка . 96 (4): 575–85. дои : 10.1016/S0092-8674(00)80661-3 . ПМИД 10052459 . S2CID 13912324 .
- ^ Ролеф Бен-Шахар Т., Кастильо А.Г., Осборн М.Дж., Борден К.Л., Корнблатт Дж., Верро А. (декабрь 2009 г.). «Два принципиально различных пептида взаимодействия PCNA способствуют функции фактора сборки хроматина 1» . Молекулярная и клеточная биология . 29 (24): 6353–65. дои : 10.1128/MCB.01051-09 . ПМЦ 2786881 . ПМИД 19822659 .
- ^ Хуан С., Чжоу Х., Кацманн Д., Хохштрассер М., Атанасова Е., Чжан З. (сентябрь 2005 г.). «Rtt106p представляет собой гистоновый шаперон, участвующий в подавлении активности, опосредованном гетерохроматином» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (38): 13410–5. Бибкод : 2005PNAS..10213410H . дои : 10.1073/pnas.0506176102 . ПМЦ 1224646 . ПМИД 16157874 .
- ^ Ито Т., Тайлер Дж. К., Балджер М., Кобаяши Р., Кадонага Дж. Т. (октябрь 1996 г.). «АТФ-облегченная сборка хроматина с нуклеоплазминоподобным белком Drosophila melanogaster» . Журнал биологической химии . 271 (40): 25041–8. дои : 10.1074/jbc.271.40.25041 . ПМИД 8798787 .
- ^ Гассер Р., Коллер Т., Сого Дж. М. (май 1996 г.). «Стабильность нуклеосом на репликационной вилке». Журнал молекулярной биологии . 258 (2): 224–39. дои : 10.1006/jmbi.1996.0245 . ПМИД 8627621 .
- ^ Мезельсон М., Шталь Ф.В. (июль 1958 г.). «Репликация ДНК в Escherichia coli» . Труды Национальной академии наук Соединенных Штатов Америки . 44 (7): 671–82. Бибкод : 1958ПНАС...44..671М . дои : 10.1073/pnas.44.7.671 . ПМК 528642 . ПМИД 16590258 .
- ^ Белл С.П., Кагуни Дж.М. (июнь 2013 г.). «Загрузка геликазы в хромосомном начале репликации» . Перспективы Колд-Спринг-Харбор в биологии . 5 (6): а010124. doi : 10.1101/cshperspect.a010124 . ПМК 3660832 . ПМИД 23613349 .