Хеликаза
Геликазы представляют собой класс ферментов, которые считаются жизненно важными для всех организмов . организма Их основная функция — распаковка генетического материала . Хеликазы — это моторные белки , которые движутся направленно вдоль нуклеиновой кислоты фосфодиэфирного остова , разделяя две гибридизированные цепи нуклеиновой кислоты (следовательно, гелик- + -аза ), используя энергию АТФ гидролиза . Существует множество геликаз, представляющих большое разнообразие процессов, в которых необходимо катализировать разделение цепей. Примерно 1% генов эукариот кодируют хеликазы. [ 1 ]
Геном человека кодирует 95 неизбыточных геликаз: 64 РНК-хеликазы и 31 ДНК-хеликазу. [ 2 ] Многие клеточные процессы, такие как репликация ДНК , транскрипция , трансляция , рекомбинация , репарация ДНК и биогенез рибосом , включают разделение цепей нуклеиновых кислот, что делает необходимым использование геликаз. Некоторые специализированные хеликазы также участвуют в распознавании нуклеиновых кислот вируса при инфекции и выполняют иммунологическую функцию. Хеликаза — это фермент, который играет решающую роль в процессах репликации и репарации ДНК. Его основная функция — раскручивать двухцепочечную молекулу ДНК путем разрыва водородных связей между комплементарными парами оснований, позволяя нитям ДНК разделиться. При этом создается репликационная вилка, которая служит матрицей для синтеза новых цепей ДНК. Хеликаза является важным компонентом клеточных механизмов, обеспечивающим точную репликацию ДНК и сохранение генетической информации. ДНК-геликаза катализирует регрессию. RecG и фермент PriA работают вместе, перематывая дуплекс ДНК, создавая соединение Холлидея. RecG высвобождает связанные белки, а хеликаза PriA облегчает перезагрузку ДНК для возобновления репликации ДНК. RecG заменяет одноцепочечный связывающий белок (SSB), который регулирует сайты загрузки геликазной вилки во время регрессии вилки. Белок SSB взаимодействует с ДНК-хеликазами PriA и RecG, восстанавливая остановившиеся вилки репликации ДНК. Эти ферменты должны связаться с SSB-хеликазой, чтобы загрузиться на остановившиеся вилки. Термическое скольжение и связывание дуплекса ДНК, возможно, поддерживаются клиновым доменом ассоциации RecG с линкером SSB. В регрессионной реакции, поддерживаемой RecG и ATPHollidayjunctions, создаются для последующей обработки.
Функция
[ редактировать ]Хеликазы часто используются для разделения цепей ДНК двойной спирали или самоотжигающейся молекулы РНК с использованием энергии гидролиза АТФ — процесса, характеризующегося разрывом водородных связей между отожженными нуклеотидными основаниями . Они также действуют, удаляя белки, связанные с нуклеиновыми кислотами, и катализируют гомологичную рекомбинацию ДНК . [ 3 ] метаболическим процессам РНК, таким как трансляция, транскрипция, биогенез рибосом , сплайсинг РНК , транспорт РНК, редактирование РНК и деградация РНК. Геликазы способствуют [ 3 ] Хеликазы постепенно перемещаются вдоль одной цепи нуклеиновой кислоты дуплекса с направленностью и процессивностью, специфичными для каждого конкретного фермента.
Геликазы принимают различные структуры и состояния олигомеризации . В то время как DnaB -подобные хеликазы раскручивают ДНК в кольцеобразные гексамеры , было показано, что другие ферменты активны в виде мономеров или димеров . Исследования показали, что хеликазы могут действовать пассивно, ожидая, пока произойдет некатализируемое раскручивание, а затем перемещаясь между смещенными цепями. [ 4 ] или может играть активную роль в катализе разделения цепей, используя энергию, вырабатываемую при гидролизе АТФ. [ 5 ] В последнем случае геликаза действует аналогично активному мотору, раскручиваясь и перемещаясь вдоль своего субстрата как прямой результат своей АТФазной активности. [ 6 ] Хеликазы могут обрабатываться гораздо быстрее in vivo, чем in vitro, из-за присутствия дополнительных белков, которые способствуют дестабилизации вилочного соединения. [ 6 ]

Активационный барьер активности геликазы
[ редактировать ]Ферментативное действие геликазы, например раскручивание нуклеиновых кислот, достигается за счет снижения активационного барьера ( ) каждого конкретного действия. [ 7 ] [ 5 ] [ 8 ] [ 9 ] Активационный барьер является результатом различных факторов и может быть определен как


где
- = количество развернутых пар оснований (б/с),
- = свободная энергия образования пары оснований,
- = уменьшение свободной энергии за счет геликазы, и
- = уменьшение свободной энергии за счет расстегивающих сил.




Факторы, которые влияют на высоту активационного барьера, включают: специфическую последовательность нуклеиновой кислоты задействованной молекулы, количество участвующих пар оснований, напряжение, присутствующее на репликационной вилке, и силы дестабилизации. [ 7 ] [ 5 ] [ 8 ] [ 9 ]
Активные и пассивные геликазы
[ редактировать ]Размер активационного барьера, который необходимо преодолеть геликазе, способствует ее классификации как активной или пассивной геликазы. В пассивных хеликазах существует значительный активационный барьер (определяемый как , где – постоянная Больцмана и – температура системы). Из-за этого значительного активационного барьера на ход его раскручивания в значительной степени влияет последовательность раскручивающихся нуклеиновых кислот внутри молекулы и наличие сил дестабилизации, действующих на репликационную вилку. [ 7 ] [ 5 ] [ 8 ] [ 9 ] Определенные комбинации нуклеиновых кислот уменьшают скорость раскручивания (например, гуанин и цитозин ), в то время как различные дестабилизирующие силы могут увеличивать скорость раскручивания. [ 5 ] [ 8 ] [ 9 ] В пассивных системах скорость раскручивания ( ) меньше скорости транслокации ( ) (транслокация вдоль одноцепочечной нуклеиновой кислоты, оцНК), поскольку она зависит от временного распутывания пар оснований на репликационной вилке, чтобы определить скорость ее раскручивания. [ 7 ] [ 5 ] [ 8 ] [ 9 ]





В активных хеликазах , где в системе отсутствует значительный барьер, поскольку хеликаза может дестабилизировать нуклеиновые кислоты, раскручивая двойную спираль с постоянной скоростью, независимо от последовательности нуклеиновой кислоты. В активных хеликазах ближе к , из-за активной способности геликазы непосредственно дестабилизировать репликационную вилку, способствуя раскручиванию. [ 7 ] [ 5 ] [ 8 ] [ 9 ]



Активные хеликазы демонстрируют сходное поведение при воздействии на обе двухцепочечные нуклеиновые кислоты, дцНК или оцНК, в отношении скорости раскручивания и скорости транслокации, причем в обеих системах и примерно равны.
Эти две категории геликаз также можно моделировать как механизмы. В таких моделях пассивные геликазы концептуализируются как броуновские храповые механизмы, приводимые в движение тепловыми флуктуациями и последующими анизотропными градиентами в решетке ДНК. Активные геликазы, напротив, концептуализируются как шаговые двигатели, также известные как двигатели с силовым ходом, использующие для продвижения либо конформационный «дюймовый червяк», либо «ходячий» механизм «рука над рукой». [ 10 ] В зависимости от организма такое перемещение по спирали может происходить при скоростях вращения в диапазоне 5000 об/мин. [ 11 ] до 10 000 [ 12 ] об/мин
История ДНК-хеликаз
[ редактировать ]ДНК-хеликазы были обнаружены в E. coli в 1976 году. Эта хеликаза была описана как «фермент, раскручивающий ДНК», который «обнаружено, что он денатурирует дуплексы ДНК в АТФ-зависимой реакции без заметного разрушения». [ 13 ] Первая эукариотическая ДНК-геликаза была обнаружена в 1978 году в лилии. [ 14 ] С тех пор ДНК-хеликазы были открыты и выделены у других бактерий, вирусов, дрожжей, мух и высших эукариот. [ 15 ] К настоящему времени выделено не менее 14 различных геликаз из одноклеточных организмов, 6 геликаз из бактериофагов, 12 из вирусов, 15 из дрожжей, 8 из растений, 11 из тимуса теленка и около 25 геликаз из клеток человека. [ 16 ] Ниже представлена история открытия геликазы:
- 1976 – Открытие и выделение E. coli. ДНК-геликазы на основе [ 13 ]
- 1978 – Открытие первых эукариотических ДНК-хеликаз, выделенных из лилии. [ 14 ]
- 1982 - «Белок гена 41 Т4» - первая зарегистрированная ДНК-геликаза бактериофага. [ 15 ]
- 1985 – Первые ДНК-хеликазы млекопитающих выделены из тимуса теленка. [ 17 ]
- 1986 - Большой опухолевый антиген SV40 объявлен как вирусная хеликаза (первый зарегистрированный вирусный белок, который, как было установлено, служит ДНК-хеликазой). [ 18 ]
- 1986 – АТФаза III, дрожжевой белок, признан ДНК-хеликазой. [ 19 ]
- 1988 - Открытие семи консервативных аминокислотных доменов, которые были определены как мотивы геликазы.
- 1989 – Обозначение ДНК-хеликаз суперсемейства I и суперсемейства II. [ 20 ]
- 1989 – Идентификация семейства геликаз DEAD box. [ 21 ]
- 1990 – Выделение ДНК-хеликазы человека. [ 22 ]
- 1992 - Выделение первой зарегистрированной митохондриальной ДНК-геликазы (из бычьего мозга). [ 23 ]
- 1996 – Отчет об открытии первой очищенной ДНК-хеликазы хлоропластов из гороха. [ 24 ]
- 2002 г. – Выделение и характеристика первой биохимически активной ДНК-геликазы малярийного паразита – Plasmodium cynomolgi . [ 25 ]
Конструктивные особенности
[ редактировать ]Общая функция хеликаз объясняет тот факт, что они обнаруживают определенную степень аминокислотных гомологии последовательностей ; все они обладают мотивами последовательности, расположенными внутри их первичной структуры , участвующими в связывании АТФ , гидролизе АТФ и транслокации вдоль нуклеиновой кислоты субстрата . Вариабельная часть аминокислотной последовательности связана с особенностями каждой геликазы.
Присутствие этих мотивов геликазы позволяет приписать предполагаемую геликазную активность данному белку, но не обязательно подтверждает, что он является активной геликазой. Однако консервативные мотивы подтверждают эволюционную гомологию ферментов. На основе этих геликазных мотивов был выделен ряд суперсемейств геликаз.
Суперсемьи
[ редактировать ]Хеликазы классифицируются на 6 групп (суперсемейств) на основе их общих мотивов последовательности. [ 26 ] Хеликазы, не образующие кольцевую структуру, относятся к суперсемействам 1 и 2, а геликазы, образующие кольца, входят в состав суперсемейств от 3 до 6. [ 27 ] Хеликазы также классифицируются как α или β в зависимости от того, работают ли они с одноцепочечной или двухцепочечной ДНК ; α-хеликазы работают с одноцепочечной ДНК , а β-хеликазы — с двухцепочечной ДНК . Их также классифицируют по полярности транслокации. Если транслокация происходит 3'-5', то геликаза относится к типу А; если транслокация происходит 5'-3', это тип B. [ 26 ]
- Суперсемейство 1 (SF1) . Это суперсемейство можно далее подразделить на хеликазы SF1A и SF1B. [ 26 ] Геликазы этой группы могут иметь транслокационную полярность либо 3'-5' (подсемейство SF1A), либо 5'-3' (подсемейство SF1B). [ 26 ] [ 28 ] Наиболее известными хеликазами SF1A являются Rep и UvrD у грамотрицательных бактерий и PcrA-хеликаза у грамположительных бактерий. [ 26 ] Наиболее известными хеликазами группы SF1B являются хеликазы RecD и Dda. [ 26 ] Они имеют складчатое ядро, подобное RecA. [ 27 ]
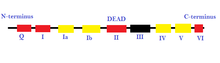
- Суперсемейство 2 (SF2) : это самая большая группа геликаз, которые участвуют в различных клеточных процессах. [ 26 ] [ 2 ] Для них характерно наличие девяти консервативных мотивов: Q, I, Ia, Ib и II–VI. [ 2 ] Эта группа в основном состоит из РНК-хеликаз DEAD-бокса. [ 27 ] Некоторые другие геликазы, включенные в SF2, представляют собой семейство RecQ-подобных и Snf2-подобных ферментов. [ 26 ] Большинство геликаз SF2 относятся к типу А, за некоторыми исключениями, такими как семейство XPD. [ 26 ] Они имеют складчатое ядро, подобное RecA. [ 27 ]
- Суперсемейство 3 (SF3) : Суперсемейство 3 состоит из геликаз AAA+, кодируемых в основном небольшими ДНК-вирусами и некоторыми крупными нуклеоцитоплазматическими ДНК-вирусами. [ 29 ] [ 30 ] Они имеют направленность транслокации 3'-5', что означает, что все они являются хеликазами типа А. [ 26 ] Наиболее известной геликазой SF3 является хеликаза Е1 вируса папилломы. [ 26 ]
- Суперсемейство 4 (SF4) : Все хеликазы семейства SF4 имеют полярность типа B (5'-3'). У них есть складка RecA. [ 26 ] Наиболее изученной SF4-хеликазой является gp4 бактериофага Т7. [ 26 ]
- Суперсемейство 5 (SF5) : белки Rho соответствуют группе SF5. У них есть складка RecA. [ 26 ]
- Суперсемейство 6 (SF6) : они содержат ядро AAA+, которое не включено в классификацию SF3. [ 26 ] Некоторыми белками в группе SF6 являются: поддерживающие мини-хромосомы MCM , RuvB, RuvA и RuvC. [ 26 ]
Все хеликазы являются членами семейства, содержащего P-петлю или мотив Уокера .
Геликазные нарушения и заболевания
[ редактировать ]Мутации геликазы ATRX
[ редактировать ]Ген ATRX кодирует АТФ-зависимую хеликазу ATRX (также известную как XH2 и XNP) из семейства подгруппы SNF2, которая, как полагают, отвечает за такие функции, как ремоделирование хроматина, регуляция генов и метилирование ДНК. [ 31 ] [ 32 ] [ 33 ] [ 34 ] Эти функции помогают предотвратить апоптоз, что приводит к регуляции размера коры, а также способствуют выживанию гиппокампа и корковых структур, влияя на память и обучение. [ 31 ] Эта хеликаза расположена на Х-хромосоме (Xq13.1-q21.1), в прицентромерном гетерохроматине и связывается с белком гетерохроматина 1 . [ 31 ] [ 33 ] Исследования показали, что ATRX играет роль в метилировании рДНК и необходим для эмбрионального развития. [ 35 ] Мутации были обнаружены во всем белке ATRX , причем более 90% из них расположены в доменах цинкового пальца и геликазы. [ 36 ] Мутации ATRX могут привести к умственной отсталости, связанной с Х-сцепленной альфа-талассемией ( синдром ATR-X ). [ 31 ]
Было обнаружено, что различные типы мутаций, обнаруженных в ATRX, связаны с ATR-X, включая наиболее часто одноосновательные миссенс-мутации, а также нонсенс-мутации, мутации сдвига рамки считывания и делеции. [ 34 ] Характеристики ATR-X включают: микроцефалию, аномалии скелета и лица, умственную отсталость, аномалии половых органов, судороги, ограниченное использование речи и ее способности, а также альфа-талассемию. [ 31 ] [ 35 ] [ 32 ] Фенотип, наблюдаемый у ATR-X, предполагает, что мутация гена ATRX вызывает снижение экспрессии генов, таких как гены альфа-глобина. [ 32 ] До сих пор неизвестно, что вызывает проявление различных характеристик ATR-X у разных пациентов. [ 35 ]
Точковые мутации геликазы XPD
[ редактировать ]XPD (пигментный фактор D Xeroderma, также известный как белок ERCC2) представляет собой 5'-3' суперсемейство II, АТФ-зависимую хеликазу, содержащую домены железо-серного кластера. [ 26 ] [ 37 ] Было показано, что наследственные точковые мутации в XPD-хеликазе связаны с расстройствами ускоренного старения, такими как синдром Коккейна (CS) и трихотиодистрофия (TTD). [ 38 ] Синдром Кокейна и трихотиодистрофия представляют собой нарушения развития, связанные с чувствительностью к ультрафиолетовому излучению и преждевременным старением, а синдром Коккейна проявляется тяжелой умственной отсталостью с момента рождения. [ 38 ] Мутация XPD-хеликазы также связана с пигментной ксеродермией (XP), заболеванием, характеризующимся чувствительностью к ультрафиолетовому излучению и приводящим к увеличению развития рака кожи в несколько 1000 раз. [ 38 ]
XPD является важным компонентом комплекса TFIIH , фактора транскрипции и репарации в клетке. [ 38 ] [ 39 ] [ 40 ] [ 41 ] [ 42 ] В составе этого комплекса он способствует эксцизионной репарации нуклеотидов путем раскручивания ДНК. [ 38 ] TFIIH помогает восстанавливать поврежденную ДНК, например, поврежденную солнцем. [ 38 ] [ 39 ] [ 40 ] [ 41 ] [ 42 ] Мутация в хеликазе XPD, которая помогает формировать этот комплекс и способствует его функции, вызывает чувствительность к солнечному свету, наблюдаемую при всех трех заболеваниях, а также повышенный риск рака, наблюдаемый при XP, и преждевременное старение, наблюдаемое при трихотиодистрофии и синдроме Коккейна. [ 38 ]
XPD-хеликазные мутации, приводящие к трихотиодистрофии, обнаруживаются по всему белку в различных местах, участвующих в белок-белковых взаимодействиях. [ 38 ] Эта мутация приводит к нестабильности белка из-за его неспособности образовывать стабилизирующие взаимодействия с другими белками в точках мутации. [ 38 ] Это, в свою очередь, дестабилизирует весь комплекс TFIIH, что приводит к дефектам механизмов транскрипции и репарации клетки. [ 38 ]
Было высказано предположение, что мутации геликазы XPD, приводящие к синдрому Коккейна, могут быть результатом мутаций внутри XPD, вызывающих ригидность белка и последующую неспособность переключиться с функций репарации на функции транскрипции из-за «блокировки» режима репарации. [ 38 ] Это может привести к тому, что геликаза разрежет сегменты ДНК, предназначенные для транскрипции. [ 38 ] Хотя текущие данные указывают на дефект геликазы XPD, приводящий к потере гибкости белка в случаях синдрома Кокейна, до сих пор неясно, как эта структура белка приводит к симптомам, описанным при синдроме Кокейна. [ 38 ]
При пигментной ксеродерме мутация XPD-хеликазы существует в месте связывания АТФ или ДНК. [ 38 ] В результате образуется структурно функциональная хеликаза, способная облегчать транскрипцию, однако она ингибирует ее функцию по раскручиванию ДНК и репарации ДНК. [ 38 ] Отсутствие способности клеток восстанавливать мутации, например, вызванные воздействием солнечных лучей, является причиной высокой заболеваемости раком у пациентов с пигментной ксеродермией.
Мутации семейства RecQ
[ редактировать ]
Хеликазы RecQ (3'-5') относятся к группе геликаз Superfamily II, которые помогают поддерживать стабильность генома и подавлять нежелательную рекомбинацию. [ 43 ] [ 44 ] Дефицит и/или мутации в хеликазах семейства RecQ демонстрируют аберрантную генетическую рекомбинацию и/или репликацию ДНК, что приводит к хромосомной нестабильности и общему снижению способности к пролиферации. [ 43 ] мутации в хеликазах семейства RecQ BLM, RECQL4 Было показано, что и WRN, которые играют роль в регуляции гомологичной рекомбинации, приводят к аутосомно-рецессивным заболеваниям: синдрому Блума (BS), синдрому Ротмунда-Томсона (RTS) и синдрому Вернера (WS). ), соответственно. [ 44 ] [ 45 ]
Синдром Блума характеризуется предрасположенностью к раку с ранним началом, средний возраст начала заболевания составляет 24 года. [ 44 ] [ 46 ] В клетках пациентов с синдромом Блума наблюдаются высокая частота реципрокного обмена между сестринскими хроматидами (СХЭ) и чрезмерное хромосомное повреждение. [ 47 ] Есть данные, позволяющие предположить, что BLM играет роль в восстановлении нарушенной репликации ДНК в репликационных вилках. [ 47 ]
Синдром Вернера — это расстройство преждевременного старения, симптомы которого включают раннее начало атеросклероза, остеопороза и других возрастных заболеваний, высокую частоту сарком и смерть, часто наступающую от инфаркта миокарда или рака на 4-6-м десятилетии жизни. [ 44 ] [ 48 ] Клетки пациентов с синдромом Вернера демонстрируют сниженную репродуктивную продолжительность жизни с хромосомными разрывами и транслокациями, а также большими делециями хромосомных компонентов, вызывающими геномную нестабильность. [ 48 ]
Синдром Ротмунда-Томсона, также известный как врожденная пойкилодермия , характеризуется преждевременным старением, аномалиями кожи и скелета, сыпью, пойкилодермией , ювенильной катарактой и предрасположенностью к раковым заболеваниям, таким как остеосаркома. [ 44 ] [ 49 ] В клетках больных синдромом Ротмунда-Томсона обнаружены хромосомные перестройки, вызывающие геномную нестабильность. RecQ — это семейство ферментов ДНК-хеликаз, которые обнаружены в различных организмах, включая бактерии, археи и эукариоты (например, люди). Эти ферменты играют важную роль в метаболизме ДНК во время репликации, рекомбинации и репарации ДНК. У человека известно пять белков-хеликаз RecQ: RecQ1, BLM, WRN, RecQ4 и RecQ5. Мутации в некоторых из этих генов связаны с генетическими нарушениями. Например, мутации в гене BLM вызывают синдром Блума, который характеризуется повышенным риском развития рака и других проблем со здоровьем. [ 50 ] Мутации в гене WRN приводят к синдрому Вернера — состоянию, характеризующемуся преждевременным старением и повышенным риском возрастных заболеваний. Хеликазы RecQ имеют решающее значение для поддержания стабильности и целостности генома. Они помогают предотвратить накопление генетических аномалий, которые могут привести к таким заболеваниям, как рак. Целостность генома зависит от семейства ДНК-хеликаз RecQ, которое включает процессы репарации ДНК, рекомбинации, репликации и транскрипции. Нестабильность генома и раннее старение — это состояния, возникающие в результате мутаций в геликазах RecQ человека. [ 51 ] Хеликаза RecQ Sgs1 отсутствует в дрожжевых клетках, что делает их полезными моделями для понимания аномалий клеток человека и функции геликазы RecQ. [ 52 ] Член семейства геликаз RecQ, RECQ1, связан с небольшим количеством редких генетических раковых заболеваний у людей. Он участвует в транскрипции, клеточном цикле и репарации ДНК. Согласно недавним исследованиям, миссенс-мутации в гене RECQ1 могут играть роль в развитии семейного рака молочной железы. ДНК-хеликазы часто притягиваются к участкам повреждения ДНК и необходимы для репликации, рекомбинации, репарации и транскрипции клеточной ДНК. Химические манипуляции с их молекулярными процессами могут изменить скорость деления раковых клеток, а также эффективность транзакций и клеточный гомеостаз. Вызванный малыми молекулами захват ДНК-хеликаз, типа метаболического белка ДНК, может иметь пагубные последствия для быстро пролиферирующих раковых клеток, что может быть эффективным при лечении рака.
Во время мейоза двухцепочечные разрывы ДНК и другие повреждения ДНК в хроматидах восстанавливаются путем гомологичной рекомбинации с использованием в качестве матрицы либо сестринской хроматиды , либо гомологичной несестринской хроматиды. Эта репарация может привести к образованию кроссинговерного (CO) или, чаще, некроссинговерного (NCO) рекомбинанта. В дрожжах Schizosaccharomyces pombe ДНК- хеликаза FmI1 семейства FANCM управляет образованием рекомбинации NCO во время мейоза. [ 53 ] также Хеликаза Rqh1 типа RecQ управляет мейотической рекомбинацией NCO. [ 54 ] Эти хеликазы, благодаря своей способности раскручивать промежуточные соединения D-петли , способствуют рекомбинации NCO посредством процесса зависимого от синтеза отжига цепи .
В растении Arabidopsis thaliana хеликаза FANCM способствует NCO и препятствует образованию рекомбинантов CO. [ 55 ] Другая хеликаза, RECQ4A/B, также независимо восстанавливает CO. Было высказано предположение, что CO ограничены из-за долгосрочных издержек рекомбинации CO, то есть разрушения благоприятных генетических комбинаций аллелей, созданных в результате прошлого естественного отбора . [ 55 ]
РНК-хеликазы
[ редактировать ]
РНК-хеликазы необходимы для большинства процессов метаболизма РНК, таких как биогенез рибосом , сплайсинг пре-мРНК и инициация трансляции . Они также играют важную роль в распознавании вирусных РНК. [ 56 ] РНК-хеликазы участвуют в обеспечении противовирусного иммунного ответа, поскольку они могут идентифицировать чужеродные РНК у позвоночных. Около 80% всех вирусов являются РНК-вирусами и содержат собственные РНК-хеликазы. [ 57 ] Дефектные РНК-хеликазы связаны с раком, инфекционными заболеваниями и нейродегенеративными расстройствами. [ 56 ] Некоторые неврологические расстройства, связанные с дефектными РНК-хеликазами, включают: боковой амиотрофический склероз , спинальную мышечную атрофию , спиноцеребеллярную атаксию 2-го типа , болезнь Альцгеймера и синдром летальной врожденной контрактуры . [ 57 ]
РНК-хеликазы и ДНК-хеликазы можно обнаружить вместе во всех суперсемействах геликаз, за исключением SF6. [ 58 ] [ 59 ] Все идентифицированные к настоящему времени эукариотические РНК-хеликазы не образуют кольца и являются частью SF1 и SF2. С другой стороны, кольцевые РНК-хеликазы обнаружены у бактерий и вирусов. [ 56 ] Однако не все РНК-хеликазы проявляют геликазную активность, определяемую ферментативной функцией, т.е. белки семейства Swi/Snf. Хотя эти белки несут типичные мотивы геликазы, гидролизуют АТФ зависимым от нуклеиновой кислоты образом и построены вокруг ядра геликазы, в целом никакой активности раскручивания не наблюдается. [ 60 ]
РНК-хеликазы, которые действительно проявляют раскручивающую активность, характеризуются по крайней мере двумя различными механизмами: каноническим раскручиванием дуплекса и локальным разделением цепи. Каноническое раскручивание дуплекса представляет собой ступенчатое направленное разделение дуплексной цепи, как описано выше, для раскручивания ДНК. Однако локальное разделение цепи происходит посредством процесса, при котором фермент геликаза загружается в любое место дуплекса. Обычно этому способствует одноцепочечный участок РНК, а загрузка фермента сопровождается связыванием АТФ. [ 61 ] Как только геликаза и АТФ связываются, происходит локальное разделение цепей, которое требует связывания АТФ, но не самого процесса гидролиза АТФ. [ 62 ] Имея меньшее количество пар оснований, дуплекс затем диссоциирует без дополнительной помощи фермента. Этот режим раскручивания используется коробчатыми хеликазами DEAD/DEAH . [ 63 ]
База данных РНК-хеликаз [ 64 ] в настоящее время доступен в Интернете и содержит полный список РНК-хеликаз с такой информацией, как последовательность, структура, биохимические и клеточные функции. [ 56 ]
Диагностические инструменты для измерения геликазы
[ редактировать ]Измерение и мониторинг активности геликазы
[ редактировать ]используются различные методы Для измерения активности геликазы in vitro . Эти методы варьируются от качественных анализов (анализы, которые обычно дают результаты, не связанные со значениями или измерениями) до количественных (анализы с численными результатами, которые можно использовать в статистическом и численном анализе). В 1982–1983 годах был разработан первый прямой биохимический метод измерения активности геликазы. [ 15 ] [ 65 ] Этот метод получил название «анализ смещения нитей».
- Анализ смещения цепи включает радиоактивную метку дуплексов ДНК. После обработки хеликазой одноцепочечную ДНК визуально определяют как отдельную от двухцепочечной ДНК с помощью неденатурирующего PAGE-электрофореза . После обнаружения одноцепочечной ДНК количественно определяют количество радиоактивной метки, находящейся на одноцепочечной ДНК, чтобы получить числовое значение степени раскручивания двухцепочечной ДНК. Анализ смещения нити приемлем для качественного анализа, его неспособность отображать результаты более чем для одного момента времени, его временные затраты и его зависимость от радиоактивных соединений для мечения оправдывают необходимость разработки диагностики, которая может контролировать активность геликазы в режиме реального времени. .
Позже были разработаны и другие методы, которые включали в себя некоторые, если не все из следующего: высокопроизводительную механику, использование нерадиоактивной маркировки нуклеотидов, более быстрое время реакции/меньшие затраты времени, мониторинг активности геликазы в реальном времени (вместо этого используется кинетическое измерение). конечной точки/анализа одной точки). Эти методологии включают в себя: «метод быстрого гашения потока, анализы на основе флуоресценции, анализы фильтрации, анализ сцинтилляционной близости с временным разрешением , анализ флуоресцентного резонансного переноса энергии , анализ, основанный на технологии флэш-пластин, гомогенные анализы тушения флуоресценции с временным разрешением и электрохемилюминесценцию. -на основе геликазных анализов». [ 16 ] С использованием специализированных математических уравнений некоторые из этих анализов можно использовать для определения того, сколько пар оснований нуклеотидов может разрушить хеликаза за гидролиз 1 молекулы АТФ. [ 66 ]
Также имеются коммерчески доступные диагностические наборы. Одним из таких наборов является диагностический анализ «Trupoint» от компании PerkinElmer , Inc. Этот анализ представляет собой анализ гашения флуоресценции с временным разрешением, в котором используется технология PerkinElmer «SignalClimb», основанная на двух метках, которые связываются в непосредственной близости друг от друга, но на противоположных концах. Нити ДНК. Одна метка представляет собой флуоресцентный хелат лантаноида, который служит меткой, которую контролируют с помощью подходящего 96/384-луночного считывателя планшетов. Другая метка — органическая молекула-гаситель. В основе этого анализа лежит «гашение» или подавление сигнала хелата лантаноидов с помощью органической молекулы-гасителя, когда они находятся в непосредственной близости – как это было бы, когда дуплекс ДНК находится в своем нативном состоянии. При хеликазной активности дуплекса метки-гасители и лантаниды разделяются по мере раскручивания ДНК. Эта потеря близости сводит на нет способность тушителей подавлять сигнал лантаноидов, вызывая заметное увеличение флуоресценции, которое отражает количество раскрученной ДНК и может использоваться в качестве количественного измерения активности геликазы. Выполнение и использование методов флуоресцентной визуализации одиночных молекул с упором на методы, которые включают оптический захват в сочетании с эпифлуоресцентной визуализацией, а также поверхностную иммобилизацию в сочетании с визуализацией флуоресценции полного внутреннего отражения. В сочетании с микроканальными проточными ячейками и микрофлюидным контролем они позволяют отображать и отслеживать отдельные флуоресцентно меченные молекулы белка и ДНК, обеспечивая измерение раскручивания и транслокации ДНК с разрешением одной молекулы. [ 67 ]
Определение полярности геликазы
[ редактировать ]Полярность геликазы, которую также называют «направленностью», определяется как направление (характеризуемое как 5’→3’ или 3’→5’) движения геликазы на одноцепочечной ДНК/РНК, вдоль которой она движется. Это определение полярности жизненно важно, например. определение того, прикрепляется ли тестируемая геликаза к ведущей цепи ДНК или к отстающей цепи ДНК. Чтобы охарактеризовать эту особенность геликазы, в качестве субстрата используется частично дуплексная ДНК, которая имеет центральную одноцепочечную область ДНК с разной длиной дуплексных участков ДНК (одна короткая область, которая проходит 5'→3', и одна более длинная область, которая проходит 3'). '→5') по обе стороны этой области. [ 68 ] Как только геликаза добавляется к этой центральной одноцепочечной области, полярность определяется путем характеристики вновь образованной одноцепочечной ДНК.
См. также
[ редактировать ]- хромодомен- ДНК-связывающий белок хеликазы: CHD1 , CHD1L , CHD2 , CHD3 , CHD4 , CHD5 , CHD6 , CHD7 , CHD8 , CHD9.
- Геликаза DEAD-бокса / DEAD/DEAH-бокса : DDX3X , DDX5 , DDX6 , DDX10 , DDX11 , DDX12 , DDX58 , DHX8 , DHX9 , DHX37 , DHX40 , DHX58.
- ASCC3 , BLM , BRIP1 , DNA2 , FBXO18 , FBXO30 , HELB , HELLS , HELQ , HELZ , HFM1 , HLTF , IFIH1 , NAV2 , PIF1 , RECQL , RTEL1 , SHPRH , SMARCA4 , SMARCAL1 , WRN , WRNIP1
- База данных РНК-хеликаз
Ссылки
[ редактировать ]- ^ Ву Ю (2012). «Размотка и перемотка: двойные грани геликазы?» . Журнал нуклеиновых кислот . 2012 : 140601. doi : 10.1155/2012/140601 . ПМЦ 3409536 . ПМИД 22888405 .
- ^ Перейти обратно: а б с Умате П., Тутея Н., Тутея Р. (январь 2011 г.). «Полногеномный комплексный анализ геликаз человека» . Коммуникативная и интегративная биология . 4 (1): 118–137. дои : 10.4161/cib.13844 . ПМК 3073292 . ПМИД 21509200 .
- ^ Перейти обратно: а б Патель СС, Донмез I (июль 2006 г.). «Механизмы геликаз» . Журнал биологической химии . 281 (27): 18265–18268. дои : 10.1074/jbc.R600008200 . ПМИД 16670085 .
- ^ Лайоннет Т., Спиринг М.М., Бенкович С.Дж., Бенсимон Д., Крокетт В. (декабрь 2007 г.). «Наблюдение за геликазой gp41 бактериофага Т4 в реальном времени выявляет механизм раскручивания» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (50): 19790–19795. Бибкод : 2007PNAS..10419790L . дои : 10.1073/pnas.0709793104 . ПМК 2148377 . ПМИД 18077411 .
- ^ Перейти обратно: а б с д и ж г Джонсон Д.С., Бай Л., Смит Б.И., Патель С.С., Ван М.Д. (июнь 2007 г.). «Исследования одиночных молекул показывают динамику раскручивания ДНК кольцевой геликазой Т7» . Клетка . 129 (7): 1299–1309. дои : 10.1016/j.cell.2007.04.038 . ПМЦ 2699903 . ПМИД 17604719 .
- ^ Перейти обратно: а б «Исследователи разгадали загадку разделения нитей ДНК» . 3 июля 2007 г. Проверено 5 июля 2007 г.
- ^ Перейти обратно: а б с д и Беттертон, доктор медицины, Юлихер Ф (31 августа 2005 г.). «Ошибка: Открытие двойных цепей нуклеиновых кислот хеликазами: активное или пассивное открытие [Phys. Rev. E 71, 011904 (2005)]» . Физический обзор E . 72 (2): 029906. Бибкод : 2005PhRvE..72b9906B . дои : 10.1103/PhysRevE.72.029906 .
- ^ Перейти обратно: а б с д и ж Маносас М., Си XG, Бенсимон Д., Крокетт В. (сентябрь 2010 г.). «Активные и пассивные механизмы геликаз» . Исследования нуклеиновых кислот . 38 (16): 5518–5526. дои : 10.1093/nar/gkq273 . ПМЦ 2938219 . ПМИД 20423906 .
- ^ Перейти обратно: а б с д и ж Харильо Х., Ибарра Б., Као-Гарсия Ф.Д. (2021). «Репликация ДНК: in vitro анализ и модели данных манипуляций с одиночными молекулами » . Журнал вычислительной и структурной биотехнологии . 19 : 3765–3778. дои : 10.1016/j.csbj.2021.06.032 . ПМЦ 8267548 . ПМИД 34285777 .
- ^ Ву, К.Г. и Спайс, М.: Обзор: Что такое геликазы? В: Шпионы, М. (Ред.): [1] . Springer Science+Business Media, Нью-Йорк, 2013 г.
- ^ «Биохимия Кевина Ахерна (BB 451/551) в Университете штата Орегон» . oregonstate.edu . Архивировано из оригинала 26 января 2021 г. Проверено 03 января 2024 г.
- ^ Библиотека 3D-анимации; Репликация: [2] (Дополнительно)
- ^ Перейти обратно: а б Абдель-Монем М., Дюрвальд Х., Хоффманн-Берлинг Х. (июнь 1976 г.). «Ферментативное раскручивание ДНК. 2. Разделение цепи АТФ-зависимым ферментом раскручивания ДНК» . Европейский журнал биохимии . 65 (2): 441–449. дои : 10.1111/j.1432-1033.1976.tb10359.x . ПМИД 133023 .
- ^ Перейти обратно: а б Хотта Ю, Стерн Х (май 1978 г.). «Белок, раскручивающий ДНК из мейотических клеток лилии». Биохимия . 17 (10): 1872–1880. дои : 10.1021/bi00603a011 . ПМИД 207302 .
- ^ Перейти обратно: а б с Венкатесан М., Сильвер Л.Л., Носсал Н.Г. (октябрь 1982 г.). «Белок гена 41 бактериофага Т4, необходимый для синтеза РНК-праймеров, также является ДНК-хеликазой» . Журнал биологической химии . 257 (20): 12426–12434. дои : 10.1016/S0021-9258(18)33731-1 . ПМИД 6288720 .
- ^ Перейти обратно: а б Тутея Н., Тутея Р. (май 2004 г.). «Прокариотические и эукариотические ДНК-хеликазы. Основные молекулярные моторные белки для клеточного механизма» . Европейский журнал биохимии . 271 (10): 1835–1848. дои : 10.1111/j.1432-1033.2004.04093.x . ПМЦ 7164108 . ПМИД 15128294 .
- ^ Хюбшер У, Сталдер Х.П. (август 1985 г.). «ДНК-геликаза млекопитающих» . Исследования нуклеиновых кислот . 13 (15): 5471–5483. дои : 10.1093/нар/13.15.5471 . ПМК 321884 . ПМИД 3162158 .
- ^ Шталь Х., Дрёге П., Книпперс Р. (август 1986 г.). «ДНК-хеликазная активность большого опухолевого антигена SV40» . Журнал ЭМБО . 5 (8): 1939–1944. дои : 10.1002/j.1460-2075.1986.tb04447.x . ПМК 1167061 . ПМИД 3019672 .
- ^ Сугино А., Рю Б.Х., Сугино Т., Наумовски Л., Фридберг ЕС (сентябрь 1986 г.). «Новая ДНК-зависимая АТФаза, которая стимулирует ДНК-полимеразу I дрожжей и обладает активностью по раскручиванию ДНК» . Журнал биологической химии . 261 (25): 11744–11750. дои : 10.1016/S0021-9258(18)67306-5 . ПМИД 3017945 .
- ^ Горбаленя А.Е., Кунин Е.В., Донченко А.П., Блинов В.М. (июнь 1989 г.). «Два родственных суперсемейства предполагаемых геликаз, участвующих в репликации, рекомбинации, репарации и экспрессии геномов ДНК и РНК» . Исследования нуклеиновых кислот . 17 (12): 4713–4730. дои : 10.1093/нар/17.12.4713 . ПМК 318027 . ПМИД 2546125 .
- ^ Линдер, П., Ласко, П.Ф., Эшбернер, М., Лерой, П., Нильсон, П.Дж., Ниши, К., Шнайр, Дж., Слонимски, П.П. (1989). Природа (Лондон) 337, 121–122.
- ^ Тутея Н., Тутея Р., Рахман К., Канг Л.И., Фаласки А. (декабрь 1990 г.). «ДНК-геликаза из клеток человека» . Исследования нуклеиновых кислот . 18 (23): 6785–6792. дои : 10.1093/нар/18.23.6785 . ПМЦ 332732 . ПМИД 1702201 .
- ^ Хеман Г.Л., Хаусвирт В.В. (сентябрь 1992 г.). «ДНК-геликаза из митохондрий млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (18): 8562–8566. Бибкод : 1992PNAS...89.8562H . дои : 10.1073/pnas.89.18.8562 . ПМК 49960 . ПМИД 1326759 .
- ^ Тутеджа Н., Фан Т.Н., Тевари К.К. (май 1996 г.). «Очистка и характеристика ДНК-хеликазы из хлоропласта гороха, которая транслоцируется в направлении от 3' к 5'» . Европейский журнал биохимии . 238 (1): 54–63. doi : 10.1111/j.1432-1033.1996.0054q.x . ПМИД 8665952 .
- ^ Тутеджа Р., Малхотра П., Сонг П., Тутеджа Н., Чаухан В.С. (2002). «Выделение и характеристика гомолога eIF-4A из Plasmodium cynomolgi». Молекулярная и биохимическая паразитология . 124 (1–2): 79–83. дои : 10.1016/S0166-6851(02)00205-0 . ПМИД 12387853 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д Синглтон М.Р., Диллингем М.С., Вигли Д.Б. (2007). «Структура и механизм геликаз и транслоказ нуклеиновых кислот». Ежегодный обзор биохимии . 76 : 23–50. doi : 10.1146/annurev.biochem.76.052305.115300 . ПМИД 17506634 .
- ^ Перейти обратно: а б с д Фэрман-Уильямс М.Э., Гюнтер У.П., Янковски Э. (июнь 2010 г.). «Геликазы SF1 и SF2: семейные дела» . Современное мнение в области структурной биологии . 20 (3): 313–324. дои : 10.1016/j.sbi.2010.03.011 . ПМК 2916977 . ПМИД 20456941 .
- ^ Стелтер М., Акаджауи С., МакСвини С., Тимминс Дж. (2013). «Структурное и механистическое понимание раскручивания ДНК Deinococcus radiodurans UvrD» . ПЛОС ОДИН . 8 (10): e77364. Бибкод : 2013PLoSO...877364S . дои : 10.1371/journal.pone.0077364 . ПМК 3797037 . ПМИД 24143224 .
- ^ Айер Л.М., Аравинд Л., Кунин Е.В. (декабрь 2001 г.). «Общее происхождение четырех различных семейств крупных эукариотических ДНК-вирусов» . Журнал вирусологии . 75 (23): 11720–11734. doi : 10.1128/JVI.75.23.11720-11734.2001 . ПМЦ 114758 . ПМИД 11689653 .
- ^ Айер Л.М., Лейпе Д.Д., Кунин Е.В., Аравинд Л. (2004). «Эволюционная история и классификация высшего порядка AAA + АТФаз». Журнал структурной биологии . 146 (1–2): 11–31. дои : 10.1016/j.jsb.2003.10.010 . ПМИД 15037234 .
- ^ Перейти обратно: а б с д и Роперс Х.Х., Хамель, Британская Колумбия (январь 2005 г.). «Х-сцепленная умственная отсталость». Обзоры природы. Генетика . 6 (1): 46–57. дои : 10.1038/nrg1501 . ПМИД 15630421 . S2CID 427210 .
- ^ Перейти обратно: а б с Гиббонс Р.Дж., Пикетс Дж., Виллард Л., Хиггс Д.Р. (март 1995 г.). «Мутации в предполагаемом глобальном регуляторе транскрипции вызывают Х-сцепленную умственную отсталость с альфа-талассемией (синдром ATR-X)» . Клетка . 80 (6): 837–845. дои : 10.1016/0092-8674(95)90287-2 . ПМИД 7697714 .
- ^ Перейти обратно: а б Интернет-база данных белков Nextprot. «ATRX-регулятор транскрипции ATRX». , Проверено 12 ноября 2012 г.
- ^ Перейти обратно: а б Пикетс DJ, Хиггс Д.Р., Бачу С., Блейк DJ, Куоррел О.В., Гиббонс Р.Дж. (декабрь 1996 г.). «ATRX кодирует нового члена семейства белков SNF2: мутации указывают на общий механизм, лежащий в основе синдрома ATR-X» . Молекулярная генетика человека . 5 (12): 1899–1907. дои : 10.1093/hmg/5.12.1899 . ПМИД 8968741 .
- ^ Перейти обратно: а б с Гиббонс Р. (май 2006 г.). «Альфа-талассемия-умственная отсталость, Х-сцепленная» . Сиротский журнал редких заболеваний . 1:15 . дои : 10.1186/1750-1172-1-15 . ПМЦ 1464382 . ПМИД 16722615 .
- ^ Стивенсон Р.Э. (1993). Адам М.П., Эверман Д.Б., Мирзаа Г.М., Пагон Р.А., Уоллес С.Е., Бин Л.Дж., Грипп К.В., Амемия А. (ред.). «Синдром умственной отсталости, сцепленный с альфа-талассемией». Джин Обзоры . Сиэтл (Вашингтон): Вашингтонский университет, Сиэтл. ПМИД 20301622 .
- ^ Рудольф Дж, Руйон К, Шварц-Линек Ю, Уайт МФ (январь 2010 г.). «Геликаза XPD раскручивает пузырьковые структуры и не останавливается из-за повреждений ДНК, удаляемых путем эксцизионной репарации нуклеотидов» . Исследования нуклеиновых кислот . 38 (3): 931–941. дои : 10.1093/нар/gkp1058 . ПМК 2817471 . ПМИД 19933257 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Фан Л., Фасс Д.О., Ченг К.Дж., Арвай А.С., Хаммел М., Робертс В.А. и др. (май 2008 г.). «Структура и активность XPD-хеликазы: понимание фенотипов рака и старения в результате мутаций XPD» . Клетка . 133 (5): 789–800. дои : 10.1016/j.cell.2008.04.030 . ПМК 3055247 . ПМИД 18510924 .
- ^ Перейти обратно: а б Лэне Ж.П., Мокет В., Эгли Ж.М. (2006). «Ферментативная активность TFIIH в транскрипции и эксцизионной репарации нуклеотидов». Восстановление ДНК, Часть А. Методы энзимологии. Том. 408. стр. 246–263. дои : 10.1016/S0076-6879(06)08015-3 . ISBN 9780121828134 . ПМИД 16793373 .
- ^ Перейти обратно: а б Тироде Ф., Буссо Д., Коин Ф., Эгли Дж.М. (январь 1999 г.). «Восстановление транскрипционного фактора TFIIH: присвоение функций трем ферментативным субъединицам: XPB, XPD и cdk7» . Молекулярная клетка . 3 (1): 87–95. дои : 10.1016/S1097-2765(00)80177-X . ПМИД 10024882 .
- ^ Перейти обратно: а б Сунг П., Байи В., Вебер С., Томпсон Л.Х., Пракаш Л., Пракаш С. (октябрь 1993 г.). «Ген пигментной ксеродерма группы D человека кодирует ДНК-хеликазу». Природа . 365 (6449): 852–855. Бибкод : 1993Natur.365..852S . дои : 10.1038/365852a0 . ПМИД 8413672 . S2CID 4334960 .
- ^ Перейти обратно: а б Шеффер Л., Рой Р., Гумберт С., Монколлин В., Вермюлен В., Хоймейкерс Дж.Х. и др. (апрель 1993 г.). «Геликаза репарации ДНК: компонент основного фактора транскрипции BTF2 (TFIIH)». Наука 260 (5104): 58–63. Бибкод : 1993Sci...260... 58S дои : 10.1126/science.8465201 . ПМИД 8465201 .
- ^ Перейти обратно: а б Ханада К., Хиксон ID (сентябрь 2007 г.). «Молекулярная генетика нарушений геликазы RecQ» . Клеточные и молекулярные науки о жизни . 64 (17): 2306–2322. дои : 10.1007/s00018-007-7121-z . ПМЦ 11136437 . ПМИД 17571213 . S2CID 29287970 .
- ^ Перейти обратно: а б с д и Опреско П.Л., Ченг В.Х., Бор В.А. (апрель 2004 г.). «Связь биохимии геликазы RecQ и болезней человека» . Журнал биологической химии . 279 (18): 18099–18102. дои : 10.1074/jbc.R300034200 . ПМИД 15023996 .
- ^ Оуян К.Дж., Ву Л.Л., Эллис Н.А. (2008). «Гомологичная рекомбинация и поддержание целостности генома: рак и старение через призму геликаз RecQ человека». Механизмы старения и развития . 129 (7–8): 425–440. дои : 10.1016/j.mad.2008.03.003 . ПМИД 18430459 . S2CID 6804631 .
- ^ Эллис Н.А., Гроден Дж., Йе Т.З., Строген Дж., Леннон Дж., Чоччи С. и др. (ноябрь 1995 г.). «Продукт гена синдрома Блума гомологичен геликазам RecQ» . Клетка . 83 (4): 655–666. дои : 10.1016/0092-8674(95)90105-1 . ПМИД 7585968 .
- ^ Перейти обратно: а б Селак Н., Бахрати Ч.З., Шевелев И., Дитши Т., ван Лун Б., Джейкоб А. и др. (сентябрь 2008 г.). «Геликаза синдрома Блума (BLM) физически и функционально взаимодействует с р12, наименьшей субъединицей ДНК-полимеразы дельта человека» . Исследования нуклеиновых кислот . 36 (16): 5166–5179. дои : 10.1093/нар/gkn498 . ПМЦ 2532730 . ПМИД 18682526 .
- ^ Перейти обратно: а б Грей М.Д., Шен Дж.К., Камат-Лёб А.С., Бланк А., Софер Б.Л., Мартин Г.М. и др. (сентябрь 1997 г.). «Белок синдрома Вернера представляет собой ДНК-геликазу». Природная генетика . 17 (1): 100–103. дои : 10.1038/ng0997-100 . ПМИД 9288107 . S2CID 20587915 .
- ^ Китао С., Симамото А., Гото М., Миллер Р.В., Смитсон В.А., Линдор Н.М., Фуруичи Ю. (май 1999 г.). «Мутации в RECQL4 вызывают часть случаев синдрома Ротмунда-Томсона». Природная генетика . 22 (1): 82–84. дои : 10.1038/8788 . ПМИД 10319867 . S2CID 195211275 .
- ^ Гупта, Соня Видуши; Шмидт, Кристина Хильдегард (18 февраля 2020 г.). «Поддержание целостности генома дрожжей с помощью ДНК-хеликаз семейства RecQ» . Гены . 11 (2): 205. doi : 10.3390/genes11020205 . ISSN 2073-4425 . ПМК 7074392 . ПМИД 32085395 .
- ^ Дебнат С., Шарма С. Хеликаза RECQ1 в стабильности генома и раке. Гены. 2020:11. doi: 10.3390/genes11060622], Дебнат С., Шарма С. Хеликаза RECQ1 в стабильности генома и раке. Гены. 2020:11. doi: 10.3390/genes11060622].
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) ; Отсутствует или пусто|title=( справка ) CS1 maint: несколько имен: список авторов ( ссылка ) CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Хендриксон, Вашингтон; Уорд, КБ (27 октября 1975 г.). «Атомные модели полипептидных остовов миогемеритрина и гемеритрина» . Связь с биохимическими и биофизическими исследованиями . 66 (4): 1349–1356. дои : 10.1016/0006-291x(75)90508-2 . ISSN 1090-2104 . ПМИД 5 .
- ^ Лоренц А., Осман Ф., Сан В., Нанди С., Штайнахер Р., Уитби MC (июнь 2012 г.). «Ортолог FANCM делящихся дрожжей управляет некроссинговерной рекомбинацией во время мейоза» . Наука . 336 (6088): 1585–1588. Бибкод : 2012Sci...336.1585L . дои : 10.1126/science.1220111 . ПМЦ 3399777 . ПМИД 22723423 .
- ^ Лоренц А., Мехатс А., Осман Ф., Уитби MC (декабрь 2014 г.). «Паралоги и медиаторы Rad51/Dmc1 противостоят ДНК-хеликазам, ограничивая образование гибридной ДНК и способствуя кроссинговерам во время мейотической рекомбинации» . Исследования нуклеиновых кислот . 42 (22): 13723–13735. дои : 10.1093/nar/gku1219 . ПМЦ 4267644 . ПМИД 25414342 .
- ^ Перейти обратно: а б Сегела-Арно М., Крисмани В., Ларшевек С., Мазель Дж., Фрогер Н., Шойнар С. и др. (апрель 2015 г.). «Множественные механизмы ограничивают мейотические кроссинговеры: TOP3α и два гомолога BLM противодействуют кроссинговерам параллельно с FANCM» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (15): 4713–4718. Бибкод : 2015PNAS..112.4713S . дои : 10.1073/pnas.1423107112 . ПМК 4403193 . ПМИД 25825745 .
- ^ Перейти обратно: а б с д Янковский А., Гюнтер УП, Янковский Э. (январь 2011 г.). «База данных РНК-хеликаз» . Исследования нуклеиновых кислот . 39 (Проблема с базой данных): D338–D341. дои : 10.1093/нар/gkq1002 . ПМК 3013637 . ПМИД 21112871 .
- ^ Янковский Э., Фэрман-Уильямс М.Э. (2010). «Введение в РНК-хеликазы: суперсемейства, семейства и основные темы». Янковский Э (ред.). РНК-геликазы (RSC Biomolecular Sciences) . Кембридж, Англия: Королевское химическое общество. п. 5. ISBN 978-1-84755-914-2 .
- ^ Ранджи А., Борис-Лори К. (2010). «РНК-хеликазы: новые роли в репликации вируса и врожденный ответ хозяина» . Биология РНК . 7 (6): 775–787. дои : 10.4161/rna.7.6.14249 . ПМК 3073335 . ПМИД 21173576 .
- ^ Янковский Э. (январь 2011 г.). «РНК-хеликазы в работе: связывание и перегруппировка» . Тенденции биохимических наук . 36 (1): 19–29. дои : 10.1016/j.tibs.2010.07.008 . ПМК 3017212 . ПМИД 20813532 .
- ^ Ян К., Дель Кампо М., Ламбовиц А.М., Янковски Э. (октябрь 2007 г.). «Белки DEAD-box раскручивают дуплексы путем локального разделения цепей» . Молекулярная клетка . 28 (2): 253–263. doi : 10.1016/j.molcel.2007.08.016 . ПМИД 17964264 .
- ^ Лю Ф., Патнэм А., Янковски Э. (декабрь 2008 г.). «Гидролиз АТФ необходим для рециркуляции белка DEAD-бокса, но не для раскручивания дуплекса» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (51): 20209–20214. Бибкод : 2008PNAS..10520209L . дои : 10.1073/pnas.0811115106 . ПМЦ 2629341 . ПМИД 19088201 .
- ^ Ярмоскайте И, Рассел Р. (2011). «Белки DEAD-бокса как РНК-хеликазы и шапероны» . Междисциплинарные обзоры Wiley. РНК . 2 (1): 135–152. дои : 10.1002/wrna.50 . ПМК 3032546 . ПМИД 21297876 .
- ^ «Индекс /» . www.rnahelicase.org . Архивировано из оригинала 18 декабря 2014 г. Проверено 7 декабря 2012 г.
- ^ Мэтсон С.В., Табор С., Ричардсон CC (ноябрь 1983 г.). «Ген 4 белка бактериофага Т7. Характеристика геликазной активности» . Журнал биологической химии . 258 (22): 14017–14024. дои : 10.1016/S0021-9258(17)44018-X . ПМИД 6315716 .
- ^ Сарлос К., Гимеси М., Ковач М. (июнь 2012 г.). «Геликаза RecQ транслоцируется по одноцепочечной ДНК с умеренной процессивностью и сильной механохимической связью» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (25): 9804–9809. Бибкод : 2012PNAS..109.9804S . дои : 10.1073/pnas.1114468109 . ПМЦ 3382518 . ПМИД 22665805 .
- ^ Паванкумар Т.Л., Экселл Дж.К., Ковальчиковски СК (1 января 2016 г.). «Прямая флуоресцентная визуализация транслокации и раскручивания отдельных геликаз ДНК». Одномолекулярная энзимология: флуоресцентные и высокопроизводительные методы . Методы энзимологии. Том. 581. стр. 1–32. дои : 10.1016/bs.mie.2016.09.010 . ISBN 9780128092675 . ПМЦ 5854184 . ПМИД 27793277 .
- ^ Боровец Ю.А. (1996). «ДНК-геликазы». В ДеПамфилисе М.Л. (ред.). Репликация ДНК в эукариотических клетках . Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. стр. 545–574. ISBN 978-0-87969-459-3 . OCLC 246537432 .
Внешние ссылки
[ редактировать ]- ДНК + геликазы в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- РНК + геликазы Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)