Комплекс распознавания происхождения
| Субъединица 2 комплекса распознавания происхождения | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ОРК2 | ||
| Пфам | PF04084 | ||
| ИнтерПро | ИПР007220 | ||
| |||
| Субъединица 3 комплекса распознавания происхождения (ORC) N-конец | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ORC3_N | ||
| Пфам | PF07034 | ||
| ИнтерПро | ИПР010748 | ||
| |||
| Субъединица 6 комплекса распознавания происхождения (ORC6) | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ОРК6 | ||
| Пфам | PF05460 | ||
| ИнтерПро | ИПР008721 | ||
| |||
В молекулярной биологии комплекс распознавания происхождения ( ORC ) представляет собой многосубъединичный ДНК- связывающий комплекс (6 субъединиц), который связывается у всех эукариот и архей -зависимым образом АТФ с началами репликации . Субъединицы этого комплекса кодируются генами ORC1 , ORC2 , ORC3 , ORC4 , ORC5 и ORC6 . [1] [2] [3] ORC является центральным компонентом репликации эукариотической ДНК и остается связанным с хроматином в репликации точках начала на протяжении всего клеточного цикла . [4]
ORC направляет репликацию ДНК по всему геному и необходим для ее инициации. [5] [6] [7] ORC и Noc3p, связанные в начале репликации, служат основой для сборки пререпликационного комплекса (pre-RC), который включает Cdc6 , Tah11 (он же Cdt1 ) и комплекс Mcm2 - Mcm7 . [8] [9] [10] [11] Сборка Pre-RC во время G1 необходима для лицензирования репликации хромосом S перед синтезом ДНК во время -фазы . [12] [13] [14] Регулируемое клеточным циклом фосфорилирование Orc2, Orc6, Cdc6 и MCM циклин -зависимой протеинкиназой Cdc28 регулирует инициацию репликации ДНК, включая блокирование повторной инициации в G2 / фазе M . [4] [15] [16] [17]
ORC присутствует на протяжении всего клеточного цикла, связанный с началом репликации, но активен только в позднем митозе и раннем G1 .
У дрожжей ORC также играет роль в установлении молчания в типа спаривания локусах Hidden MAT Left (HML) и Hidden MAT Right (HMR). [5] [6] [7] ORC участвует в сборке транскрипционно молчащего хроматина Sir1 на HML и HMR путем привлечения сайленсера к сайленсерам HML и HMR. [7] [18] [19]
И Orc1, и Orc5 связывают АТФ, хотя только Orc1 обладает АТФазной активностью. [20] Связывание . АТФ с помощью Orc1 необходимо для связывания ORC с ДНК и важно для жизнеспособности клеток [11] АТФазная активность Orc1 участвует в формировании пре-РК. [21] [22] [23] Связывание АТФ с помощью Orc5 имеет решающее значение для стабильности ORC в целом. Orc1-5 только субъединицы Для связывания источника необходимы ; Orc6 необходим для поддержания однажды сформированных пре-RC. [24] Взаимодействия внутри ORC позволяют предположить, что Orc2-3-6 может образовывать основной комплекс. [4] В отчете 2020 года предполагается, что ORC почкующихся дрожжей димеризуется в зависимости от клеточного цикла, чтобы контролировать лицензирование. [25] [26]
Белки
[ редактировать ]В составе ORC присутствуют следующие белки:
| С. cerevisiae | С. насос | Д. меланогастер | Позвоночные животные |
|---|---|---|---|
| ОРЦ 1-6 | ОРЦ 1-6 | ОРЦ 1-6 | ОРЦ 1-6 |
| Cdc6 | Cdc18 | Cdc6 | Cdc6 |
| Cdt1/Tah11/Sid2 | Кдт1 | ДУП | Cdt1/RLF-B |
| Мкм2 | Mcm2/Cdc19/Nda1 | Мкм2 | Мкм2 |
| Мкм3 | Мкм3 | Мкм3 | Мкм3 |
| Cdc54/Mcm4 | Cdc21 | ДПА | Мкм4 |
| Кдк46/Мкм5 | Мкм5/Нда4 | Мкм5 | Мкм5 |
| Мкм6 | Мкм6/Мис5 | Мкм6 | Мкм6 |
| Cdc47/Mcm7 | Мкм7 | Мкм7 | mcm7 |
Археи представляют собой упрощенную версию ORC, Mcm, и, как следствие, объединенный pre-RC. Вместо использования шести различных белков mcm для формирования псевдосимметричного гетерогексамера все шесть субъединиц в архейном MCM одинаковы. Обычно они имеют несколько белков, гомологичных как Cdc6, так и Orc1, некоторые из которых выполняют функцию обоих. В отличие от эукариотических орков, они не всегда образуют комплекс. Фактически, когда они формируются, они имеют различные сложные структуры. Sulfolobus Islandicus также использует гомолог Cdt1 для распознавания одного из источников репликации. [28]
Автономно реплицирующиеся последовательности
[ редактировать ]Почкующиеся дрожжи
[ редактировать ]Автономно реплицирующиеся последовательности (ARS), впервые обнаруженные у почкующихся дрожжей , являются неотъемлемой частью успеха ORC. Эти последовательности длиной 100–200 п.н. облегчают репликационную активность во время S-фазы. ARS можно поместить в любое новое место хромосом почкующихся дрожжей и облегчить репликацию из этих мест. высококонсервативная последовательность из 11 пар оснований (известная как элемент А ) необходима для функции происхождения у почкующихся дрожжей. Считается, что [27] Первоначально ORC был идентифицирован по его способности связываться с элементом A ARS у почкующихся дрожжей.
Животные
[ редактировать ]Клетки животных содержат гораздо более загадочную версию ARS, консервативные последовательности которой пока не обнаружены. Здесь источники репликации собираются в пучки, называемые репликонов кластерами . Репликоны каждого кластера одинаковы по длине, но отдельные кластеры имеют репликоны разной длины. Все эти репликоны имеют схожие основные остатки, с которыми связывается ORC, которые во многом имитируют консервативный элемент A длиной 11 п.н. Все эти кластеры одновременно активируются во время S-фазы . [27]
Роль в сборке перед RC
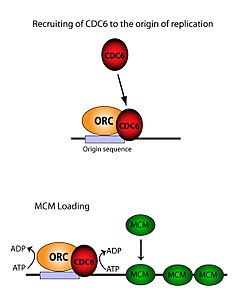
[ редактировать ]ORC необходим для загрузки комплексов MCM ( Pre-RC ) в ДНК. Этот процесс зависит от ORC, Noc3, Cdc6 и Cdt1 , включая несколько событий рекрутирования, контролируемых АТФ . Во-первых, ORC, Noc3p и Cdc6 образуют комплекс на исходной ДНК (отмеченный областями типа ARS). Новые комплексы ORC/Noc3/Cdc6 затем привлекают в этот сайт молекулы Cdt1/Mcm2-7. Как только образуется этот массивный комплекс ORC/Noc3/Cdc6/Cdt1/Mcm2-7, молекулы ORC/Noc3/Cdc6/Cdt1 работают вместе, загружая Mcm2-7 в саму ДНК путем гидролиза АТФ с помощью Cdc6. Фосфорилирующая активность Cdc6 зависит как от ORC, так и от исходной ДНК . Это приводит к снижению стабильности Cdt1 в ДНК и отпадению комплекса, что приводит к загрузке Mcm2-7 в ДНК. [29] [27] [30] [31] Решена структура ORC, MCM, а также промежуточного комплекса OCCM. [32]
![]()
Активность привязки источника
[ редактировать ]Хотя ORC состоит из шести дискретных субъединиц, только одна из них оказалась значимой — ORC1. Исследования in vivo показали, что Lys -263 и Arg -367 являются основными остатками, ответственными за точную загрузку ORC. Эти молекулы представляют собой упомянутые выше АРС. [33] ORC1 взаимодействует с АТФ и этими основными остатками, чтобы связать ORC с исходной ДНК . Было установлено, что это происходит задолго до репликации и что сам ORC уже связан с исходной ДНК к моменту любой Mcm2-7 . загрузки [31] Когда Mcm2-7 впервые загружается, он полностью окружает ДНК, и активность геликазы ингибируется. В S-фазе комплекс Mcm2-7 , взаимодействует с кофакторами геликазы Cdc45 и GINS чтобы изолировать одну цепь ДНК, раскрутить начало и начать репликацию по хромосоме . Чтобы обеспечить двунаправленную репликацию, этот процесс происходит дважды в источнике. Оба события загрузки опосредуются одним ORC посредством того же процесса, что и первый. [34]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Происхождение + распознавание + комплекс в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- ^ Датта А., Белл СП (1997). «Инициация репликации ДНК в эукариотических клетках». Ежегодный обзор клеточной биологии и биологии развития . 13 : 293–332. дои : 10.1146/annurev.cellbio.13.1.293 . ПМИД 9442876 .
- ^ Чесноков И.Н. (2007). Многофункциональность комплекса распознавания происхождения . Международный обзор цитологии. Том. 256. стр. 69–109. дои : 10.1016/S0074-7696(07)56003-1 . ISBN 9780123737007 . ПМИД 17241905 .
- ^ Jump up to: а б с Мацуда К., Макисе М., Суэясу Ю., Такехара М., Асано Т., Мидзушима Т. (декабрь 2007 г.). «Двугибридный дрожжевой анализ комплекса распознавания происхождения Saccharomyces cerevisiae: взаимодействие между субъединицами и идентификация связывающих белков». Исследование дрожжей FEMS . 7 (8): 1263–9. дои : 10.1111/j.1567-1364.2007.00298.x . ПМИД 17825065 .
- ^ Jump up to: а б Белл С.П., Стиллман Б. (май 1992 г.). «АТФ-зависимое распознавание эукариотического начала репликации ДНК мультибелковым комплексом». Природа . 357 (6374): 128–34. Бибкод : 1992Natur.357..128B . дои : 10.1038/357128a0 . ПМИД 1579162 . S2CID 4346767 .
- ^ Jump up to: а б Белл С.П., Митчелл Дж., Лебер Дж., Кобаяши Р., Стиллман Б. (ноябрь 1995 г.). «Мультидоменная структура Orc1p обнаруживает сходство с регуляторами репликации ДНК и подавления транскрипции» . Клетка . 83 (4): 563–8. дои : 10.1016/0092-8674(95)90096-9 . ПМИД 7585959 .
- ^ Jump up to: а б с Гибсон Д.Г., Белл С.П., Апарисио О.М. (июнь 2006 г.). «Анализ точки выполнения клеточного цикла функции ORC и характеристика реакции контрольной точки на инактивацию ORC в Saccharomyces cerevisiae» . Гены в клетки . 11 (6): 557–73. дои : 10.1111/j.1365-2443.2006.00967.x . ПМИД 16716188 . S2CID 22439595 .
- ^ Чжан Ю, Ю З, Фу Х, Лян С (июнь 2002 г.). «Noc3p, белок bHLH, играет важную роль в инициации репликации ДНК у почкующихся дрожжей» . Клетка . 109 (7): 849–860. дои : 10.1016/s0092-8674(02)00805-x . ПМИД 12110182 .
- ^ Рао Х., Стиллман Б. (март 1995 г.). «Комплекс распознавания происхождения взаимодействует с двусторонним сайтом связывания ДНК внутри дрожжевых репликаторов» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (6): 2224–8. Бибкод : 1995PNAS...92.2224R . дои : 10.1073/pnas.92.6.2224 . ПМК 42456 . ПМИД 7892251 .
- ^ Роули А., Кокер Дж. Х., Харвуд Дж., Диффли Дж. Ф. (июнь 1995 г.). «Сборка инициирующего комплекса в точках начала репликации почкующихся дрожжей начинается с распознавания двудольной последовательности за счет ограничения количества инициатора, ORC» . Журнал ЭМБО . 14 (11): 2631–41. дои : 10.1002/j.1460-2075.1995.tb07261.x . ПМЦ 398377 . ПМИД 7781615 .
- ^ Jump up to: а б Спек С., Чен З., Ли Х., Стиллман Б. (ноябрь 2005 г.). «АТФаза-зависимое кооперативное связывание ORC и Cdc6 с исходной ДНК» . Структурная и молекулярная биология природы . 12 (11): 965–71. дои : 10.1038/nsmb1002 . ПМЦ 2952294 . ПМИД 16228006 .
- ^ Келли Т.Дж., Браун Г.В. (2000). «Регуляция репликации хромосом». Ежегодный обзор биохимии . 69 : 829–80. doi : 10.1146/annurev.biochem.69.1.829 . ПМИД 10966477 .
- ^ Белл С.П., Дутта А. (2002). «Репликация ДНК в эукариотических клетках». Ежегодный обзор биохимии . 71 : 333–74. doi : 10.1146/annurev.biochem.71.110601.135425 . ПМИД 12045100 .
- ^ Стиллман Б. (февраль 2005 г.). «Распознавание происхождения и хромосомный цикл» . Письма ФЭБС . 579 (4): 877–84. дои : 10.1016/j.febslet.2004.12.011 . ПМИД 15680967 . S2CID 33220937 .
- ^ Вайнрайх М., Лян С., Чен Х.Х., Стиллман Б. (сентябрь 2001 г.). «Связывание циклин-зависимых киназ с ORC и Cdc6p регулирует цикл репликации хромосом» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (20): 11211–7. дои : 10.1073/pnas.201387198 . ПМК 58709 . ПМИД 11572976 .
- ^ Нгуен VQ, Co C, Ли JJ (июнь 2001 г.). «Циклин-зависимые киназы предотвращают повторную репликацию ДНК с помощью нескольких механизмов». Природа . 411 (6841): 1068–73. Бибкод : 2001Natur.411.1068N . дои : 10.1038/35082600 . ПМИД 11429609 . S2CID 4393812 .
- ^ Аршамбо В., Икуи А.Е., Драпкин Б.Дж., Кросс ФР (август 2005 г.). «Нарушение механизмов, предотвращающих повторную репликацию, запускает реакцию повреждения ДНК» . Молекулярная и клеточная биология . 25 (15): 6707–21. дои : 10.1128/MCB.25.15.6707-6721.2005 . ПМК 1190345 . ПМИД 16024805 .
- ^ Триоло Т, Стернгланц Р (май 1996 г.). «Роль взаимодействия между комплексом распознавания происхождения и SIR1 в подавлении транскрипции». Природа . 381 (6579): 251–3. Бибкод : 1996Natur.381..251T . дои : 10.1038/381251a0 . ПМИД 8622770 . S2CID 4309206 .
- ^ Фокс Калифорния, Эренхофер-Мюррей А.Э., Лу С., Райн Дж. (июнь 1997 г.). «Комплекс распознавания происхождения, SIR1, и требование S-фазы для молчания». Наука . 276 (5318): 1547–51. дои : 10.1126/science.276.5318.1547 . ПМИД 9171055 .
- ^ Клемм Р.Д., Остин Р.Дж., Белл С.П. (февраль 1997 г.). «Координационное связывание АТФ и исходной ДНК регулирует АТФазную активность комплекса распознавания происхождения» . Клетка . 88 (4): 493–502. дои : 10.1016/S0092-8674(00)81889-9 . ПМИД 9038340 .
- ^ Клемм Р.Д., Bell SP (июль 2001 г.). «АТФ, связанный с комплексом распознавания происхождения, важен для формирования preRC» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (15): 8361–7. Бибкод : 2001PNAS...98.8361K . дои : 10.1073/pnas.131006898 . ПМК 37444 . ПМИД 11459976 .
- ^ Бауэрс Дж.Л., Рэнделл Дж.К., Чен С., Белл С.П. (декабрь 2004 г.). «Гидролиз АТФ с помощью ORC катализирует повторяющуюся сборку Mcm2-7 в определенном месте начала репликации» . Молекулярная клетка . 16 (6): 967–78. doi : 10.1016/j.molcel.2004.11.038 . ПМИД 15610739 .
- ^ Рэнделл Дж.К., Бауэрс Дж.Л., Родригес Х.К., Bell SP (январь 2006 г.). «Последовательный гидролиз АТФ с помощью Cdc6 и ORC направляет загрузку геликазы Mcm2-7» . Молекулярная клетка . 21 (1): 29–39. doi : 10.1016/j.molcel.2005.11.023 . ПМИД 16387651 .
- ^ Семпл Дж.В., Да-Сильва Л.Ф., Джервис Э.Дж., Ах-Ки Дж., Аль-Аттар Х., Куммер Л. и др. (ноябрь 2006 г.). «Важная роль Orc6 в репликации ДНК посредством поддержания пререпликативных комплексов» . Журнал ЭМБО . 25 (21): 5150–8. дои : 10.1038/sj.emboj.7601391 . ПМК 1630405 . ПМИД 17053779 .
- ^ Инь Ю.К., Прашант С.Г. (июль 2021 г.). «Инициация репликации: последствия для целостности генома» . Восстановление ДНК . 103 : 103131. doi : 10.1016/j.dnarep.2021.103131 . ПМЦ 8296962 . ПМИД 33992866 .
- ^ Амин А, Ву, Р., Чунг М.Х., Скотт Дж.Ф., Ван З., Чжоу З., Лю С., Чжу Г., Вонг К.С., Ю З., Лян С. (март 2020 г.). «Основной и зависимый от клеточного цикла цикл димеризации ORC регулирует репликацию эукариотической хромосомной ДНК» . Отчеты по ячейкам . 30 (10): 3323–3338.e6. дои : 10.1016/j.celrep.2020.02.046 . ПМИД 32160540 .
- ^ Jump up to: а б с д Морган, Дэвид (2007). Клеточный цикл: принципы управления . Буквари по биологии. стр. 62–75. ISBN 978-0878935086 .
- ^ Аусянникава Д., Аллерс Т. (январь 2017 г.). «Разнообразие репликации ДНК у архей» . Гены . 8 (2): 56. doi : 10.3390/genes8020056 . ПМК 5333045 . ПМИД 28146124 .
- ^ Фернандес-Сид А., Риера А., Тогнетти С., Эррера М.С., Самель С., Эврин С. и др. (май 2013 г.). «Комплекс ORC/Cdc6/MCM2-7 образуется в результате многоступенчатой реакции и служит платформой для сборки двойного гексамера MCM» . Молекулярная клетка . 50 (4): 577–88. doi : 10.1016/j.molcel.2013.03.026 . hdl : 10044/1/19289 . ПМИД 23603117 .
- ^ Рэнделл Дж.К., Бауэрс Дж.Л., Родригес Х.К., Bell SP (январь 2006 г.). «Последовательный гидролиз АТФ с помощью Cdc6 и ORC направляет загрузку геликазы Mcm2-7» . Молекулярная клетка . 21 (1): 29–39. doi : 10.1016/j.molcel.2005.11.023 . ПМИД 16387651 .
- ^ Jump up to: а б Спек С., Чен З., Ли Х., Стиллман Б. (ноябрь 2005 г.). «АТФаза-зависимое кооперативное связывание ORC и Cdc6 с исходной ДНК» . Структурная и молекулярная биология природы . 12 (11): 965–71. дои : 10.1038/nsmb1002 . ПМЦ 2952294 . ПМИД 16228006 .
- ^ Юань З., Риера А., Бай Л., Сунь Дж., Нанди С., Спанос С. и др. (март 2017 г.). «Структурная основа репликативной загрузки геликазы Mcm2-7 с помощью ORC-Cdc6 и Cdt1» . Структурная и молекулярная биология природы . 24 (3): 316–324. дои : 10.1038/nsmb.3372 . ПМК 5503505 . ПМИД 28191893 .
- ^ Каваками Х., Охаси Э., Канамото С., Цуримото Т., Катаяма Т. (октябрь 2015 г.). «Специфическое связывание эукариотического ORC с точками начала репликации ДНК зависит от высококонсервативных основных остатков» . Научные отчеты . 5 : 14929. Бибкод : 2015NatSR...514929K . дои : 10.1038/srep14929 . ПМК 4601075 . ПМИД 26456755 .
- ^ Чистол Дж., Уолтер Дж. К. (апрель 2015 г.). «Одномолекулярная визуализация загрузки ДНК MCM2-7: увидеть — значит поверить» . Клетка . 161 (3): 429–430. дои : 10.1016/j.cell.2015.04.006 . ПМИД 25910200 .
Дальнейшее чтение
[ редактировать ]- Белл С.П. , Датта А. (июль 2002 г.). «Репликация ДНК в эукариотических клетках». Ежегодный обзор биохимии . 71 . Годовые обзоры: 333–74. doi : 10.1146/annurev.biochem.71.110601.135425 . ПМИД 12045100 .
Комплексный обзор репликации молекулярной ДНК