Моторный белок

Моторные белки — это класс молекулярных моторов , которые могут перемещаться по цитоскелету клеток. работу путем гидролиза АТФ Они преобразуют химическую энергию в механическую . жгутиков Однако вращение приводится в движение протонным насосом . [ нужна ссылка ]
Клеточные функции
[ редактировать ]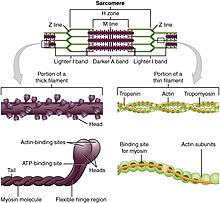
движущей силой наиболее активного транспорта белков Моторные белки являются и везикул в цитоплазме . Кинезины и цитоплазматические динеины играют важную роль во внутриклеточном транспорте, таком как аксональный транспорт , а также в формировании веретенообразного аппарата и разделении хромосом во время митоза и мейоза . Аксонемальный динеин , обнаруженный в ресничках и жгутиках , имеет решающее значение для подвижности клеток , например, в сперматозоидах , и транспорта жидкости, например, в трахее. Мышечный белок миозин «двигает» сокращение мышечных волокон у животных.
Заболевания, связанные с дефектами двигательных белков
[ редактировать ]Важность моторных белков в клетках становится очевидной, когда они не выполняют свою функцию. Например, дефицит кинезина был идентифицирован как причина болезни Шарко-Мари-Тута и некоторых заболеваний почек . Дефицит динеина может привести к хроническим инфекциям , дыхательных путей поскольку реснички не могут функционировать без динеина. Многочисленные дефициты миозина связаны с болезненными состояниями и генетическими синдромами. Поскольку миозин II необходим для сокращения мышц, дефекты мышечного миозина предсказуемо вызывают миопатии. Миозин необходим для процесса слуха из-за его роли в росте стереоцилий, поэтому дефекты в структуре белка миозина могут привести к синдрому Ушера и несиндромальной глухоте . [1]
Цитоскелетные моторные белки
[ редактировать ]Моторные белки, использующие цитоскелет для движения, делятся на две категории в зависимости от их субстрата : микрофиламенты и микротрубочки . Актиновые моторы, такие как миозин, движутся вдоль микрофиламентов за счет взаимодействия с актином , а моторы микротрубочек, такие как динеин и кинезин, движутся вдоль микротрубочек за счет взаимодействия с тубулином .
Существует два основных типа моторов микротрубочек : моторы плюсового конца и моторы минусового конца, в зависимости от направления, в котором они «идут» по кабелям микротрубочек внутри клетки.
Актиновые моторы
[ редактировать ]Миозин
[ редактировать ]Миозины — это суперсемейство актиновых моторных белков , которые преобразуют химическую энергию в форме АТФ в механическую энергию, тем самым генерируя силу и движение. Первый обнаруженный миозин, миозин II, отвечает за сокращение мышц . Миозин II представляет собой удлиненный белок, образованный из двух тяжелых цепей с моторными головками и двух легких цепей. Каждая головка миозина содержит актин и сайт связывания АТФ. Головки миозина связывают и гидролизуют АТФ, что обеспечивает энергию для движения к плюсовому концу актиновой нити. Миозин II также жизненно важен в процессе деления клеток . Например, немышечные биполярные толстые нити миозина II обеспечивают силу сокращения, необходимую для разделения клетки на две дочерние клетки во время цитокинеза. Помимо миозина II, за разнообразие движений немышечных клеток ответственны многие другие типы миозина. Например, миозин участвует во внутриклеточной организации и выпячивании богатых актином структур на поверхность клетки. Миозин V участвует в транспорте везикул и органелл. [2] [3] Миозин XI участвует в потоке цитоплазмы , при котором движение по сети микрофиламентов в клетке позволяет органеллам и цитоплазме течь в определенном направлении. [4] Известно восемнадцать различных классов миозинов. [5]
Геномное представление миозиновых моторов: [6]
- Грибы ( дрожжи ): 5
- Растения ( арабидопсис ): 17
- Насекомые ( дрозофилы ): 13
- Млекопитающие ( человек ): 40
- Хромадория ( нематода C. elegans ): 15
Моторы микротрубочек
[ редактировать ]Кинезин
[ редактировать ]Кинезины представляют собой суперсемейство родственных моторных белков, которые используют микротрубочки при антероградном движении. Они жизненно важны для формирования веретена при митотическом и мейотическом разделении хромосом во время клеточного деления, а также ответственны за перемещение митохондрий , тел Гольджи и везикул внутри эукариотических клеток . Кинезины имеют две тяжелые цепи и две легкие цепи на активный мотор. Два моторных домена глобулярной головки в тяжелых цепях могут преобразовывать химическую энергию гидролиза АТФ в механическую работу для перемещения по микротрубочкам. [7] Направление транспортировки груза может быть как к плюсовому, так и к минусовому концу, в зависимости от типа кинезина. В общем, кинезины с N-концевыми моторными доменами перемещают груз к плюсовым концам микротрубочек, расположенных на периферии клетки, тогда как кинезины с C-концевыми моторными доменами перемещают груз к минусовым концам микротрубочек, расположенных в ядре. Известны четырнадцать различных семейств кинезинов с некоторыми дополнительными кинезиноподобными белками, которые нельзя отнести к этим семействам. [8]
Геномное представление кинезиновых моторов: [6]
- Грибы ( дрожжи ): 6
- Растения ( Arabidopsis thaliana ): 61
- Насекомые ( Drosophila melanogaster ): 25
- Млекопитающие ( человек ): 45
Дайнейн
[ редактировать ]Динеины — это моторы микротрубочек, способные к ретроградному скользящему движению. Динеиновые комплексы намного крупнее и сложнее, чем кинезиновые и миозиновые моторы. Динеины состоят из двух или трех тяжелых цепей и большого и вариабельного количества связанных с ними легких цепей. Динеины управляют внутриклеточным транспортом к минус-концу микротрубочек, который находится в центре организации микротрубочек рядом с ядром. [9] Семейство динеинов имеет две основные ветви. Аксонемальные динеины облегчают биение ресничек и жгутиков за счет быстрых и эффективных скользящих движений микротрубочек. Другая ветвь — цитоплазматические динеины, которые облегчают транспорт внутриклеточных грузов. Из 15 типов аксонемного динеина только две цитоплазматические формы. известны [10]
Геномное представление динеиновых моторов: [6]
- Грибы ( дрожжи ): 1
- Растения ( Arabidopsis thaliana ): 0
- Насекомые ( Drosophila melanogaster ): 13
- Млекопитающие ( человек ): 14-15
Специализированные двигатели
[ редактировать ]В отличие от животных , грибов и несосудистых растений , в клетках цветковых растений отсутствуют динеиновые моторы. Однако они содержат большее количество различных кинезинов. Многие из этих групп кинезинов, специфичных для растений, специализируются на функциях во время растительных клеток митоза . [11] Растительные клетки отличаются от клеток животных тем, что имеют клеточную стенку . Во время митоза новая клеточная стенка строится путем образования клеточной пластинки, начинающейся в центре клетки. Этому процессу способствует фрагмопласт — набор микротрубочек, уникальный для митоза растительных клеток. Для построения клеточной пластинки и, в конечном итоге, новой клеточной стенки необходимы кинезиноподобные моторные белки. [12]
Другим моторным белком, необходимым для деления растительных клеток, является кинезин-подобный кальмодулин-связывающий белок (KCBP), который уникален для растений и состоит частично из кинезина и частично из миозина. [13]
Другие молекулярные моторы
[ редактировать ]Помимо моторных белков, описанных выше, существует еще много типов белков, способных генерировать силы и крутящий момент в клетке. Многие из этих молекулярных моторов повсеместно распространены как в прокариотических , так и в эукариотических клетках, хотя некоторые из них, например, связанные с элементами цитоскелета или хроматином , уникальны для эукариот. Двигательный белок престин , [14] экспрессируемый во наружных волосковых клетках улитки млекопитающих, вызывает механическую амплификацию в улитке. Это преобразователь напряжения в силу прямого действия, работающий с микросекундной скоростью и обладающий пьезоэлектрическими свойствами.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Хирокава Н., Такемура Р. (октябрь 2003 г.). «Биохимическая и молекулярная характеристика заболеваний, связанных с моторными белками». Тенденции биохимических наук . 28 (10): 558–65. дои : 10.1016/j.tibs.2003.08.006 . ПМИД 14559185 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (1 января 2002 г.). «Молекулярные моторы» . NCBI — Национальные институты здравоохранения .
- ^ Уоршоу, DM (февраль 2012 г.). «Наклон и вращение при движении миозина V вдоль актиновых нитей, что обнаруживается с помощью поляризации флуоресценции» . Журнал общей физиологии . 139 (2): 97–100. дои : 10.1085/jgp.201210769 . ПМЦ 3269787 . ПМИД 22291143 .
- ^ Хартман М.А., Спудич Дж.А. (апрель 2012 г.). «Краткий обзор суперсемейства миозина» . Журнал клеточной науки . 125 (Часть 7): 1627–32. дои : 10.1242/jcs.094300 . ПМЦ 3346823 . ПМИД 22566666 .
- ^ Томпсон РФ, генеральный директор Лэнгфорда (ноябрь 2002 г.). «История эволюции суперсемейства миозинов». Анатомическая запись . 268 (3): 276–89. дои : 10.1002/ar.10160 . ПМИД 12382324 . S2CID 635349 .
- ^ Перейти обратно: а б с Вейл Р.Д. (февраль 2003 г.). «Набор молекулярных моторов для внутриклеточного транспорта» . Клетка . 112 (4): 467–80. дои : 10.1016/S0092-8674(03)00111-9 . ПМИД 12600311 .
- ^ Верхей К.Дж., Каул Н., Соппина В. (01.01.2011). «Сборка кинезина и движение в клетках». Ежегодный обзор биофизики . 40 : 267–88. doi : 10.1146/annurev-biophys-042910-155310 . ПМИД 21332353 .
- ^ Мики Х., Окада Ю., Хирокава Н. (сентябрь 2005 г.). «Анализ суперсемейства кинезинов: понимание структуры и функции». Тенденции в клеточной биологии . 15 (9): 467–76. дои : 10.1016/j.tcb.2005.07.006 . ПМИД 16084724 .
- ^ Робертс А.Дж., Кон Т., Найт П.Дж., Суто К., Берджесс С.А. (ноябрь 2013 г.). «Функции и механика моторных белков динеина» . Обзоры природы. Молекулярно-клеточная биология . 14 (11): 713–26. дои : 10.1038/nrm3667 . ПМЦ 3972880 . ПМИД 24064538 .
- ^ Маллик Р., Гросс С.П. (ноябрь 2004 г.). «Молекулярные моторы: стратегии выживания» . Современная биология . 14 (22): Р971-82. дои : 10.1016/j.cub.2004.10.046 . ПМИД 15556858 . S2CID 14240073 .
- ^ Ванстрален М., Инзе Д., Гилен Д. (апрель 2006 г.). «Кинезины, специфичные для митоза у арабидопсиса» . Тенденции в науке о растениях . 11 (4): 167–75. doi : 10.1016/j.tplants.2006.02.004 . hdl : 1854/LU-364298 . ПМИД 16530461 .
- ^ Смит Л.Г. (март 2002 г.). «Цитокинез растений: едем до финиша» . Современная биология . 12 (6): Р206-8. дои : 10.1016/S0960-9822(02)00751-0 . ПМИД 11909547 .
- ^ Абдель-Гани С.Э., Дэй И.С., Симмонс, член парламента, Кугренс П., Редди А.С. (июль 2005 г.). «Происхождение и эволюция кинезинподобного кальмодулинсвязывающего белка» . Физиология растений . 138 (3): 1711–22. дои : 10.1104/стр.105.060913 . ПМК 1176440 . ПМИД 15951483 .
- ^ Даллос П., Факлер Б. (февраль 2002 г.). «Престин, новый тип моторного белка». Обзоры природы. Молекулярно-клеточная биология . 3 (2): 104–11. дои : 10.1038/nrm730 . ПМИД 11836512 . S2CID 7333228 .
Внешние ссылки
[ редактировать ]- MBInfo - Что такое моторные белки?
- Семинар Рона Вейла: «Молекулярные моторные белки»
- Биология моторных белков Институт биофизической химии , Геттинген
- Джонатан Ховард (2001), Механика моторных белков и цитоскелета. ISBN 9780878933334