Динамика белка
В молекулярной биологии обычно белки считается, что имеют уникальные структуры, определяемые их аминокислотными последовательностями. Однако белки не являются строго статичными объектами, а скорее заселяют ансамбли (иногда схожих) конформаций . Переходы между этими состояниями происходят в различных масштабах длины (от десятых ангстрем до нм) и времени (от нс до с).и были связаны с функционально значимыми явлениями, такими как аллостерическая передача сигналов. [1] и ферментативный катализ . [2]
Изучение динамики белков наиболее непосредственно связано с переходами между этими состояниями, но может также затрагивать природу и равновесные популяции самих состояний.Эти две точки зрения — кинетика и термодинамика соответственно — могут быть концептуально синтезированы в парадигме « энергетического ландшафта »: [3] высоконаселенные состояния и кинетика переходов между ними могут быть описаны глубиной энергетических ям и высотой энергетических барьеров соответственно.

Локальная гибкость: остатки и атомы
Участки белковых структур часто отклоняются от равновесного состояния.Некоторые такие отклонения являются гармоническими , например стохастические колебания химических связей и валентных углов.Другие являются ангармоничными , например, боковые цепи, которые перескакивают между отдельными дискретными минимумами энергии, или ротамеры . [4]
Доказательства локальной гибкости часто получают с помощью ЯМР-спектроскопии . Гибкие и потенциально неупорядоченные области белка можно обнаружить с помощью индекса случайной катушки . Гибкость свернутых белков можно определить путем анализа спиновой релаксации отдельных атомов белка. Гибкость также можно наблюдать на картах электронной плотности очень высокого разрешения, полученных с помощью рентгеновской кристаллографии . [5] особенно когда данные дифракции собираются при комнатной температуре вместо традиционной криогенной температуры (обычно около 100 К). [6] Информацию о частотном распределении и динамике локальной гибкости белков можно получить с помощью рамановской и с эффектом Керра. оптической спектроскопии [7] а также анизотропная микроспектроскопия [8] в терагерцовой частотной области.
: внутридоменное соединение нескольких остатков гибкость Региональная
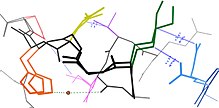
Многие остатки находятся в тесной пространственной близости в белковых структурах. Это верно для большинства остатков, которые являются смежными в первичной последовательности, но также и для многих остатков, которые расположены дистальнее по последовательности, но контактируют в окончательной складчатой структуре. Из-за этой близости энергетические ландшафты этих остатков становятся связанными на основе различных биофизических явлений, таких как водородные связи , ионные связи и взаимодействия Ван-дер-Ваальса (см. Рисунок).
Таким образом, переходы между состояниями для таких наборов остатков становятся коррелированными. [9]
Это, пожалуй, наиболее очевидно для петель, открытых на поверхности, которые часто коллективно смещаются, принимая разные конформации в разных кристаллических структурах (см. Рисунок). Однако во вторичной структуре иногда проявляется и связанная конформационная гетерогенность. [10] Например, последовательные остатки и остатки, смещенные на 4 в первичной последовательности, часто взаимодействуют в α-спиралях . Кроме того, остатки, смещенные на 2 в первичной последовательности, направляют свои боковые цепи к одной и той же стороне β-листа и расположены достаточно близко для стерического взаимодействия, как и остатки на соседних цепях одного и того же β-листа. Некоторые из этих конформационных изменений индуцируются посттрансляционными модификациями структуры белка, такими как фосфорилирование и метилирование. [10] [11]
Когда эти связанные остатки образуют пути, связывающие функционально важные части белка,они могут участвовать в аллостерической передаче сигналов.Например, когда молекула кислорода связывается с одной субъединицей тетрамера гемоглобина ,эта информация аллостерически распространяется на три другие субъединицы, тем самым увеличивая их сродство к кислороду.В этом случае связанная гибкость гемоглобина обеспечивает совместное связывание кислорода,что физиологически полезно, поскольку обеспечивает быструю подачу кислорода в легочную ткань и быструю разгрузку кислорода в тканях, лишенных кислорода (например, в мышцах).
Глобальная гибкость доменов : несколько
Наличие нескольких доменов в белках приводит к значительной гибкости и подвижности , что приводит к динамике белковых доменов . [1] Движения доменов можно сделать путем сравнения различных структур белка (как в Базе данных молекулярных движений ) или их можно напрямую наблюдать с помощью спектров. [12] [13] измерено методом нейтронной спин-эхо -спектроскопии.Их также можно предположить путем отбора проб на обширных траекториях молекулярной динамики. [14] и анализ главных компонент. [15] Движения доменов важны для:
- Транспортеры АВС [16]
- катализ [17]
- клеточная локомоция и моторные белки [18]
- образование белковых комплексов [19]
- ионные каналы [20]
- Механорецепторы и механотрансдукция [21]
- регулирующая деятельность [22]
- транспорт метаболитов через клеточные мембраны [ нужна ссылка ]
Одним из крупнейших наблюдаемых движений доменов является механизм «поворота» пируватфосфатдикиназы . Фосфоинозитидный домен переключается между двумя состояниями, чтобы перенести фосфатную группу из активного сайта нуклеотидсвязывающего домена в активный сайт фосфоенолпируват/пируватного домена. [23] Фосфатная группа перемещается на расстояние 45 Å, что приводит к перемещению домена примерно на 100 градусов вокруг одного остатка. В ферментах замыкание одного домена на другой захватывает субстрат путем индуцированного прилегания, позволяя реакции протекать контролируемым образом. Детальный анализ Герштейна привел к классификации двух основных типов движения доменов; шарнир и сдвиг. [20] Только относительно небольшая часть цепи, а именно междоменный линкер и боковые цепи, претерпевает значительные конформационные изменения при перегруппировке домена. [24]
Шарнирные движения [ править ]

Исследование Хейворда [25] обнаружили, что концы α-спиралей и β-листов в большом количестве случаев образуют шарниры. Было обнаружено, что многие петли состоят из двух элементов вторичной конструкции, действующих как петли двери, позволяя совершать открывающиеся и закрывающиеся движения. Это может возникнуть, когда две соседние цепи внутри β-листа, расположенного в одном домене, расходятся при присоединении к другому домену. Два получившихся конца затем образуют области изгиба между двумя доменами. Обнаружено, что α-спирали, которые сохраняют свою сеть водородных связей при изгибе, ведут себя как механические шарниры, накапливая «упругую энергию», которая приводит к закрытию доменов для быстрого захвата подложки. [25] Хаде и др. ал. работал над предсказанием петель [26] в любой конфигурации и в дальнейшем построил модель эластичной сети под названием hdANM. [27] который может моделировать эти движения.
От спиральной до расширенной конформации [ править ]
Взаимное превращение спиральной и вытянутой конформаций в месте доменной границы не является редкостью. В кальмодулине торсионные углы изменяются для пяти остатков в середине домена, связывающего α-спираль. Спираль расщепляется на две почти перпендикулярные спирали меньшего размера, разделенные четырьмя остатками вытянутой цепи. [28] [29]
Сдвиговые движения [ править ]
Сдвиговые движения включают небольшое скользящее движение границ раздела доменов, контролируемое боковыми цепями аминокислот внутри границы раздела. Белки, демонстрирующие сдвиговые движения, часто имеют многослойную архитектуру: укладку вторичных структур. Междоменный компоновщик играет лишь роль поддержания доменов в непосредственной близости. [ нужна ссылка ]
Движение доменов и функциональная ферментах динамика в
Анализ внутренней динамики структурно различных, но функционально сходных ферментов.подчеркнул общую взаимосвязь между расположениемактивный центр и два основных субдомена белка. Фактически, для некоторых членов суперсемейства гидролаз каталитический сайт расположен близко к границе раздела, разделяющей два основных квазижестких домена. [14] Такое позиционирование, по-видимому, способствует поддержанию точной геометрии активного сайта, в то же время допуская заметную функционально ориентированную модуляцию фланкирующих областей, возникающую в результате относительного движения двух субдоменов. [ нужна ссылка ]
для эволюции Последствия макромолекул
Имеющиеся данные свидетельствуют о том, что динамика белка важна для функционирования, например, ферментный катализ дигидрофолатредуктазы ( DHFR ),однако предполагается, что они также способствуют приобретению новых функций в ходе молекулярной эволюции . [30] Этот аргумент предполагает, что белки в ходе эволюции приобрели стабильные, в основном уникальные складчатые структуры.но неизбежная остаточная гибкость приводит к некоторой степени функциональной распущенности,которые могут быть усилены/использованы/перенаправлены последующими мутациями. [ нужна ссылка ] Исследования беспорядочных белков семейства BCL-2 показали, что динамика белков в наносекундном масштабе может играть решающую роль в поведении связывания белков и, следовательно, в беспорядочных связях. [31]
Однако растет осознание того, что неструктурированные по своей природе белки . в геномах эукариот довольно распространены [32] ставя под сомнение простейшую интерпретацию догмы Анфинсена : «последовательность определяет структуру (единственное число)».По сути, новая парадигма характеризуется добавлением двух оговорок: «последовательность и клеточная среда определяют структурный ансамбль».
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б Бу Зи, Callaway DJ (2011). «Белки движутся! Динамика белков и дальняя аллостерия в передаче сигналов в клетках». В Доневе Р. (ред.). Структура белка и болезни . Достижения в области химии белков и структурной биологии. Том. 83. Академическая пресса. стр. 163–221. дои : 10.1016/B978-0-12-381262-9.00005-7 . ISBN 9780123812629 . ПМИД 21570668 .
- ^ Фрейзер Дж.С., Кларксон М.В., Дегнан С.С., Эрион Р., Керн Д., Альбер Т. (декабрь 2009 г.). «Скрытые альтернативные структуры пролин-изомеразы, необходимые для катализа» . Природа . 462 (7273): 669–673. Бибкод : 2009Natur.462..669F . дои : 10.1038/nature08615 . ПМЦ 2805857 . ПМИД 19956261 .
- ^ Фрауэнфельдер Х., Слайгар С.Г., Волинес П.Г. (декабрь 1991 г.). «Энергетические ландшафты и движения белков». Наука . 254 (5038): 1598–1603. Бибкод : 1991Sci...254.1598F . дои : 10.1126/science.1749933 . ПМИД 1749933 .
- ^ Данбрэк, Роланд Л. (август 2002 г.). «Библиотеки ротамеров в 21 веке». Современное мнение в области структурной биологии . 12 (4): 431–440. дои : 10.1016/s0959-440x(02)00344-5 . ПМИД 12163064 .
- ^ Дэвис И.В., Арендал В.Б., Ричардсон, округ Колумбия, Ричардсон Дж.С. (февраль 2006 г.). «Движение по спине: как остов белка пожимает плечами, когда танцует боковая цепь» . Структура . 14 (2): 265–274. дои : 10.1016/j.str.2005.10.007 . ПМИД 16472746 .
- ^ Фрейзер Дж.С., ван ден Бедем Х., Самельсон А.Дж., Ланг П.Т., Холтон Дж.М., Эколс Н., Альбер Т. (сентябрь 2011 г.). «Доступ к конформационным ансамблям белков с помощью рентгеновской кристаллографии при комнатной температуре» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (39): 16247–16252. Бибкод : 2011PNAS..10816247F . дои : 10.1073/pnas.1111325108 . ПМК 3182744 . ПМИД 21918110 .
- ^ Тертон Д.А., Сенн Х.М., Харвуд Т., Лапторн А.Дж., Эллис Э.М., Винн К. (июнь 2014 г.). «Терагерцовое недостаточно затухающее колебательное движение управляет связыванием белка с лигандом в растворе» . Природные коммуникации . 5 : 3999. Бибкод : 2014NatCo...5.3999T . дои : 10.1038/ncomms4999 . ПМИД 24893252 .
- ^ Акбас, Г.; Ниссен, Калифорния; Снелл, Э.Х.; Маркельц, АГ (2014). «Белко-оптические измерения дальнодействующих колебаний белков» . Природные коммуникации . 5 : 3076. дои : 10.1038/ncomms4076 . ПМИД 24430203 .
- ^ Бу Зи, Кук Джей, Callaway DJ (сентябрь 2001 г.). «Динамические режимы и коррелирующая структурная динамика нативного и денатурированного альфа-лактальбумина». Журнал молекулярной биологии . 312 (4): 865–873. дои : 10.1006/jmbi.2001.5006 . ПМИД 11575938 .
- ^ Jump up to: Перейти обратно: а б Коста CH, Оливейра AR, Дос Сантос AM, да Коста KS, Лима AH, Алвес CN, Ламейра J (октябрь 2019 г.). «Вычислительное исследование конформационных изменений 3-гидрокси-3-метилглутарил-кофермент-редуктазы человека, индуцированных связыванием субстрата». Журнал биомолекулярной структуры и динамики . 37 (16): 4374–4383. дои : 10.1080/07391102.2018.1549508 . ПМИД 30470158 . S2CID 53717806 .
- ^ Гробан Э.С., Нараянан А., член парламента Джейкобсона (апрель 2006 г.). Шахнович Э (ред.). «Конформационные изменения в белковых петлях и спиралях, индуцированные посттрансляционным фосфорилированием» . PLOS Вычислительная биология . 2 (4): е32. Бибкод : 2006PLSCB...2...32G . дои : 10.1371/journal.pcbi.0020032 . ПМК 1440919 . ПМИД 16628247 .
- ^ Фараго Б., Ли Дж., Корнилеску Дж., Каллауэй DJ, Бу З. (ноябрь 2010 г.). «Активация наномасштабного движения доменов аллостерических белков, выявленная с помощью нейтронной спин-эхо-спектроскопии» . Биофизический журнал . 99 (10): 3473–3482. Бибкод : 2010BpJ....99.3473F . дои : 10.1016/j.bpj.2010.09.058 . ПМЦ 2980739 . ПМИД 21081097 .
- ^ Бу З, Биль Р., Монкенбуш М., Рихтер Д., Каллауэй DJ (декабрь 2005 г.). «Связанное движение белковых доменов в Taq-полимеразе, выявленное с помощью спектроскопии нейтронного спинового эха» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 102 (49): 17646–17651. Бибкод : 2005PNAS..10217646B . дои : 10.1073/pnas.0503388102 . ПМЦ 1345721 . ПМИД 16306270 .
- ^ Jump up to: Перейти обратно: а б Потестио Р., Понтиджа Ф., Микелетти С. (июнь 2009 г.). «Грубое описание внутренней динамики белков: оптимальная стратегия разложения белков на жесткие субъединицы» . Биофизический журнал . 96 (12): 4993–5002. Бибкод : 2009BpJ....96.4993P . дои : 10.1016/j.bpj.2009.03.051 . ПМК 2712024 . ПМИД 19527659 .
- ^ Барон Р., Веллор Н.А. (июль 2012 г.). «LSD1/CoREST представляет собой аллостерический наноразмерный зажим, регулируемый молекулярным распознаванием хвоста гистона H3» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (31): 12509–14. Бибкод : 2012PNAS..10912509B . дои : 10.1073/pnas.1207892109 . ПМК 3411975 . ПМИД 22802671 .
- ^ Понте-Сукре А, изд. (2009). ABC-транспортеры в микроорганизмах . Кайстер Академик. ISBN 978-1-904455-49-3 .
- ^ Камерлин С.К., Варшел А. (май 2010 г.). «На заре XXI века: является ли динамика недостающим звеном для понимания ферментативного катализа?» . Белки . 78 (6): 1339–75. дои : 10.1002/прот.22654 . ПМЦ 2841229 . ПМИД 20099310 .
- ^ Ховард Дж (2001). Механика моторных белков и цитоскелета (1-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 9780878933334 .
- ^ Callaway DJ, Мацуи Т, Вайс Т, Стингачиу ЛР, Стэнли CB, Хеллер ВТ, Бу Зи (апрель 2017 г.). «Управляемая активация наномасштабной динамики в неупорядоченном белке изменяет кинетику связывания» . Журнал молекулярной биологии . 429 (7): 987–998. дои : 10.1016/j.jmb.2017.03.003 . ПМК 5399307 . ПМИД 28285124 .
- ^ Jump up to: Перейти обратно: а б Герштейн М., Леск А.М., Чотия С. (июнь 1994 г.). «Структурные механизмы движения доменов в белках». Биохимия . 33 (22): 6739–49. дои : 10.1021/bi00188a001 . ПМИД 8204609 .
- ^ Николл И.Д., Мацуи Т., Вайс Т.М., Стэнли CB, Хеллер В.Т., Мартел А., Фараго Б., Callaway DJ, Bu Z (21 августа 2018 г.). «Структура альфа-катенина и наномасштабная динамика в растворе и в комплексе с F-актином» . Биофизический журнал . 115 (4): 642–654. Бибкод : 2018BpJ...115..642N . дои : 10.1016/j.bpj.2018.07.005 . hdl : 2436/621755 . ПМК 6104293 . ПМИД 30037495 .
- ^ Фут Д (2011). Биохимия . Воэт, Джудит Г. (4-е изд.). Хобокен, Нью-Джерси: John Wiley & Sons. ISBN 9780470570951 . OCLC 690489261 .
- ^ Херцберг О., Чен С.С., Кападиа Г., Макгуайр М., Кэрролл Л.Дж., Но С.Дж., Данауэй-Мариано Д. (апрель 1996 г.). «Механизм поворотного домена ферментативного фосфопереноса между удаленными участками реакции» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (7): 2652–7. Бибкод : 1996PNAS...93.2652H . дои : 10.1073/pnas.93.7.2652 . ПМК 39685 . ПМИД 8610096 .
- ^ Джанин Дж., Водак С.Дж. (1983). «Структурные домены в белках и их роль в динамике функции белков» . Прогресс биофизики и молекулярной биологии . 42 (1): 21–78. дои : 10.1016/0079-6107(83)90003-2 . ПМИД 6353481 .
- ^ Jump up to: Перейти обратно: а б Хейворд С. (сентябрь 1999 г.). «Структурные принципы, управляющие движением доменов в белках». Белки . 36 (4): 425–35. doi : 10.1002/(SICI)1097-0134(19990901)36:4<425::AID-PROT6>3.0.CO;2-S . ПМИД 10450084 . S2CID 29808315 .
- ^ Хаде, Пранав М.; Кумар, Амбуж; Джерниган, Роберт Л. (17 января 2020 г.). «Охарактеризация и прогнозирование белковых шарниров для понимания механизмов» . Журнал молекулярной биологии . 432 (2): 508–522. дои : 10.1016/j.jmb.2019.11.018 . ISSN 1089-8638 . ПМК 7029793 . ПМИД 31786268 .
- ^ Хаде, Пранав М.; Скарамоццино, Доменико; Кумар, Амбуж; Лачидонья, Джузеппе; Карпинтери, Альберто; Джерниган, Роберт Л. (16 ноября 2021 г.). «hdANM: новая комплексная модель динамики белковых шарниров» . Биофизический журнал . 120 (22): 4955–4965. дои : 10.1016/j.bpj.2021.10.017 . ISSN 1542-0086 . ПМЦ 8633836 . ПМИД 34687719 .
- ^ Мидор В.Е., Минс А.Р., Киочо Ф.А. (август 1992 г.). «Распознавание целевого фермента кальмодулином: 2.4 Структура комплекса кальмодулин-пептид». Наука . 257 (5074): 1251–1255. Бибкод : 1992Sci...257.1251M . дои : 10.1126/science.1519061 . ПМИД 1519061 .
- ^ Икура М., Клор Г.М., Гроненборн А.М., Чжу Г., Клее С.Б., Бакс А. (май 1992 г.). «Структура раствора комплекса кальмодулин-мишень-пептид по данным многомерного ЯМР». Наука . 256 (5057): 632–638. Бибкод : 1992Sci...256..632I . дои : 10.1126/science.1585175 . ПМИД 1585175 .
- ^ Токурики Н., Тауфик Д.С. (апрель 2009 г.). «Белковый динамизм и эволюционность». Наука . 324 (5924): 203–207. Бибкод : 2009Sci...324..203T . дои : 10.1126/science.1169375 . ПМИД 19359577 . S2CID 28576496 .
- ^ Хекмайер, Филипп Дж.; Руф, Жаннетт; Янкович, Бранка Г.; Хамм, Питер (7 марта 2023 г.). «Распущенность MCL-1 и структурная устойчивость его партнеров по связыванию». Журнал химической физики . 158 (9). arXiv : 2211.08934 . дои : 10.1063/5.0137239 .
- ^ Дайсон Х.Дж. , Райт П.П. (март 2005 г.). «Самостоятельно неструктурированные белки и их функции». Nature Reviews Молекулярно-клеточная биология . 6 (3): 197–208. дои : 10.1038/nrm1589 . ПМИД 15738986 . S2CID 18068406 .