Кинезин
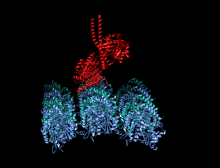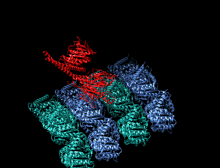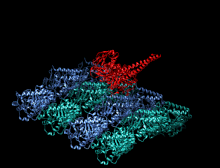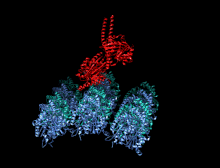

Кинезин принадлежащий - это белок, классу моторных белков, обнаруженных в эукариотических клетках. Кинеры перемещаются вдоль микротрубочек (МТ) филаментов и питаются гидролизом аденозинтрифосфата ( АТФ) (таким образом , кинезины являются АТФазами , типом фермента). Активное движение кинезинов подтверждает несколько клеточных функций, включая митоз , мейоз и транспорт клеточного груза, например, в аксональном транспорте и внутрикафлагеллярный транспорт . Большинство кинезинов идут к плюсу микротрубочки, которая в большинстве клеток влечет за собой транспортировка груза, таких как белковые и мембранные компоненты из центра клетки к периферии. [ 1 ] Эта форма транспорта известна как антероградный транспорт . Напротив, динеины - это моторные белки, которые движутся в сторону минус -конце микротрубочки при ретроградном транспорте .
Открытие
[ редактировать ]Первые кинезины, которые были обнаружены [ 2 ] В 1985 году, основанный на их подвижности в цитоплазме, экструдированной из гигантского аксона кальмара . [ 3 ]
Оботителем этой суперсемейства, Kinesin-1, был выделен как гетеротетрамерный двигатель переноса из аксонов, состоящий из четырех частей, состоящий из четырех частей: две идентичные моторные субъединицы (называемые молекулами кинезиновой тяжелой цепи (KHC)) и двух других молекул, каждая из которых известна как кинезиновый свет. Цепь (KLC). Они были обнаружены посредством очистки аффинной микротрубочек из экстрактов нейрональных клеток. [ 4 ] Впоследствии другой, гетеротримерный мотор на основе MT, направленный на основе MT с названием Kinesin-2, состоящий из двух различных моторных субъединиц, связанных с KHC и вспомогательной субъединицы «KAP», была очищена от экстрактов Echinoderm Embract [ 5 ] и наиболее известен своей ролью в транспортировке белковых комплексов ( внутрифлагеллярные транспортные частицы) вдоль аксонеем во время цилиогенеза . [ 6 ] Молекулярные генетические и геномные подходы привели к признанию того, что кинезины образуют разнообразное суперсемейство двигателей, которые ответственны за множественные внутриклеточные события подвижности в эукариотических клетках. [ 7 ] [ 8 ] [ 9 ] [ 10 ] Например, геномы млекопитающих кодируют более 40 кинезиновых белков, [ 11 ] организовано как минимум в 14 семей под названием Kinesin-1 через кинезин-14. [ 12 ]
Структура
[ редактировать ]Общая структура
[ редактировать ]Члены кинезиновой суперсемейства различаются по форме, но прототипичный мотор кинезина-1 состоит из двух молекул тяжелой цепи кинезина (KHC), которые образуют димер белка (пара молекул), которые связывают две светлые цепи (KLC), которые являются уникальными для разных карго.
Тяжелая цепь кинезина-1 содержит глобулярную головку (моторный домен) на амино-терминальном конце, соединенном с помощью короткого гибкого линкера шеи с стеблем-длинный, центральный альфа-спиральный домен катушки -который заканчивается в карбокси-терминале хвостовой домен, который связывается с световыми цепями. Стаи двух KHC переплетают образующуюся спиральную катушку , которая направляет димеризацию двух KHC. В большинстве случаев транспортируемый груз связывается с кинезиновыми световыми цепями, в последовательности мотива TPR KLC, но в некоторых случаях груз связывается с С-концевыми доменами тяжелых цепей. [ 13 ]
Кинезиновый моторный домен
[ редактировать ]| Кинезиновый моторный домен | |||
|---|---|---|---|
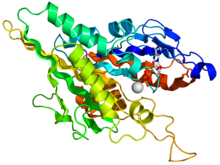 Кристаллографическая структура кинезинового моторного домена человека, изображенная как мультфильм с цветом радуги ( n-конец = синий, c-конец = красный), в комплекте с помощью ADP (диаграмма палки, углерод = белый, кислород = красный, азот = синий, фосфор = оранжевый) и магний ион (серая сфера). [ 14 ] | |||
| Идентификаторы | |||
| Символ | Кинезиновый моторный домен | ||
| Pfam | PF00225 | ||
| InterPro | IPR001752 | ||
| УМНЫЙ | SM00129 | ||
| PROSITE | PS50067 | ||
| Краткое содержание | 1bg2 / scope / supfam | ||
| CDD | CD00106 | ||
| |||
Голова является подписью кинезина, а его аминокислотная последовательность хорошо сохранилась среди различных кинезинов. Каждая голова имеет два отдельных сайта связывания : один для микротрубочки, а другая для АТФ. Связывание АТФ и гидролиз, а также высвобождение ADP изменяют конформацию доменов, связывающих микротрубочки и ориентацию линкера шеи относительно головы; Это приводит к движению кинезина. Несколько структурных элементов в голове, в том числе центральный домен бета-листа и домены Switch I и II, были замечены как опосредованные взаимодействия между двумя сайтами связывания и доменом шеи. Кинеины структурно связаны с G -белками , которые гидролизуют GTP вместо АТФ. Несколько структурных элементов распространяются между двумя семьями, в частности, домен Switch I и Switch II.

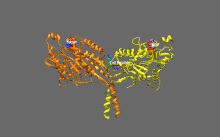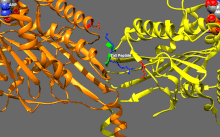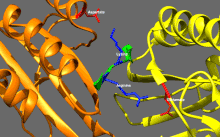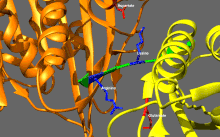
Основное регулирование кинезина
[ редактировать ]Кинеины имеют тенденцию обладать низкой базальной ферментативной активностью, которая становится значительной при активировании микротрубочек. [ 16 ] Кроме того, многие члены суперсемейства кинезина могут быть самостоятельно ингибируются путем связывания хвостового домена с моторным доменом. [ 17 ] Такое самостоятельное ингибирование может быть облегчено посредством дополнительной регуляции, такой как связывание с грузом, грузовыми адаптерами или другими микротрубочками, связанными с белками . [ 18 ] [ 19 ] [ 20 ]
Грузовой транспорт
[ редактировать ]В клетке мелкие молекулы, такие как газы и глюкоза , диффундируют , где они необходимы. Большие молекулы, синтезируемые в клеточном организме, внутриклеточные компоненты, такие как везикулы и органеллы, такие как митохондрии, слишком велики (и цитозол слишком многолюдный), чтобы иметь возможность диффундировать в свои места. Моторные белки выполняют роль транспортировки большого груза вокруг клетки в их необходимые направления. Кинеины представляют собой моторные белки, которые переносят такой груз путем однонаправленно ходя вдоль микротрубочек , отслеживает гидролизацию одной молекулы аденозинтрифосфата (АТФ) на каждом этапе. [ 21 ] Считалось, что гидролиз АТФ приводил в действие каждый шаг, высвобождаемая энергия, подталкивающая головы вперед к следующему сайту связывания. [ 22 ] Тем не менее, было предложено, чтобы голова диффундировала вперед, а сила связывания с микротрубочками - это то, что тянет груз. [ 23 ] Кроме того, вирусы, например, ВИЧ, используйте кинезины, чтобы позволить вирусным частицам перекрывать после сборки. [ 24 ]
Существуют значительные доказательства того, что грузы in vivo транспортируются несколькими двигателями. [ 25 ] [ 26 ] [ 27 ] [ 28 ]
Направление движения
[ редактировать ]Моторные белки движутся в определенном направлении вдоль микротрубочки. Микротрубочки полярны; Это означает, что головы связываются только с микротрубочками в одной ориентации, в то время как связывание АТФ дает каждому шагу свое направление через процесс, известный как линкер -линкер на шейке. [ 29 ]
Ранее было известно, что кинезин перемещает груз к плюс (+) конец микротрубочки, также известной как антероградный транспорт/ортоградный транспорт. [ 30 ] Тем не менее, недавно было обнаружено, что в почкуенных дрожжевых клетках Kinesin CIN8 (член семейства кинезин-5) также может двигаться к концу минус или ретроградный транспорт. Это означает, что эти уникальные дрожжевые гомотетрамеры имеют новую способность двигаться двунаправленно. [ 31 ] [ 32 ] [ 33 ] До сих пор было показано, что кинезин только движется к мину, когда в группе, с двигателями, скользящими в антипараллельном направлении, в попытке отделить микротрубочки. [ 34 ] Эта двойная направленность наблюдалась в идентичных условиях, когда свободные молекулы CIN8 движутся в сторону минус, но сшивая CIN8 движется к плюс концы каждой сшитой микротрубочки. В одном конкретном исследовании протестировалось скорость, с которой двигались двигатели CIN8, их результаты дали диапазон около 25-55 нм/с в направлении полюсов веретена. [ 35 ] На индивидуальной основе было обнаружено, что различными ионными условиями CIN8 Motors могут стать столь же быстрыми, как 380 нм/с. [ 35 ] Предполагается, что двунаправленность дрожжевых кинезиновых двигателей, таких как CIN8 и CUT7, является результатом связи с другими двигателями CIN8 и помогает выполнять роль динеина в почкующихся дрожжах, в отличие от гомолога человека этих двигателей, плюс Режиссер EG5. [ 36 ] Это открытие в белках семейства кинезин-14 (таких как Drosophila melanogaster NCD, почкующие дрожжи Kar3 и Arabidopsis thaliana Atk5) позволяет кинезину ходить в противоположном направлении, к микротрубочкам минус. [ 37 ] Это не типично для кинезина, скорее, исключение из нормального направления движения.

Другой тип моторного белка, известный как динеины , движется к минусу конца микротрубочки. Таким образом, они переносят груз с периферии ячейки к центру. Примером этого будет транспорт, происходящий из терминальных бутонов нейронального аксона к клеточному телу (SOMA). Это известно как ретроградный транспорт .
Механизм движения
[ редактировать ]В 2023 году сообщалось о прямой визуализации кинезина «ходьбы» вдоль микротрубочек в режиме реального времени. [ 38 ] В «ручной ручной» механизме кинезин выступает мимо друг друга, чередуя позицию ведущего. Таким образом, на каждом шаге ведущая голова становится задней головой, в то время как задняя голова становится ведущей головой.
- Этот цикл начинается с зацепленной головки, высвобождающей неорганический фосфат (PI), полученный из гидролиза АТФ.
- Тяжелая головка отсоединяется от микротрубочки и вращается в его направо, смещенное несвязанное состояние.
- Ведущая голова связывает ATP, что заставляет линкеров шеи приклеивать к нему, что перемещает заднюю голову вокруг ведущей головы в положение дальше вдоль микротрубочки в направлении движения. Тяжелая голова остается несвязанной.
- АТФ в ведущей головке гидролизован.
- Тяжелая головка выпускает свой ADP, а привязанность к микротрубочкам становится ведущей головой. [ 39 ] [ 40 ] [ 41 ] [ 42 ] [ 43 ] [ 44 ]
Теоретическое моделирование
[ редактировать ]Был предложен ряд теоретических моделей молекулярного моторного белка кинезина. [ 45 ] [ 46 ] [ 47 ] Многие проблемы возникают в теоретических исследованиях, учитывая оставшуюся неопределенности в отношении ролей белковых структур, точная энергия из АТФ превращается в механическую работу, и роли, которые играют тепловые колебания. Это довольно активная область исследований. Особенно существует необходимость для подходов, которые лучше связаны с молекулярной архитектурой белка и данными, полученными в результате экспериментальных исследований.
Динамика с одной молекулы уже хорошо описана [ 48 ] Но кажется, что эти нано -масштабные машины обычно работают в крупных командах.
Динамика одномолекулы основана на различных химических состояниях двигателя и наблюдениях о его механических этапах. [ 49 ] Для небольших концентраций аденозинфосфата поведение двигателя регулируется конкуренцией двух химиомеханических моторных циклов, которые определяют силу киоска двигателя. Третий цикл становится важным для больших концентраций ADP. [ 49 ] Модели с одним циклом также обсуждались. Seiferth et al. продемонстрировали, как величины, такие как скорость или производство энтропии изменений двигателя, когда смежные состояния объединяются в многоциклической модели, пока в конечном итоге количество циклов не будет уменьшено. [ 50 ]
Недавние экспериментальные исследования показали, что кинезины, двигаясь по микротрубочкам, взаимодействуют друг с другом, [ 51 ] [ 52 ] Взаимодействие является коротким и слабым привлекательным (1,6 ± 0,5 К б т). Одна модель, которая была разработана, учитывает эти взаимодействия частиц, [ 48 ] где динамические скорости меняются соответственно с энергией взаимодействия. Если энергия является положительной, скорость создания связей (Q) будет выше, в то время как скорость разбитых связей (R) будет ниже. Можно понять, что скорости входа и выхода в микротрубочки также будут изменены энергией (см. Рисунок 1 в ссылке 30). Если второй сайт занят, скорость входа будет α*Q, и если последний, но один сайт занят, скорость выхода будет β*r. Этот теоретический подход согласуется с результатами моделирования Монте -Карло для этой модели, особенно для ограничивающего случая очень большой негативной энергии. Нормальный полностью асимметричный простой процесс исключения для (или TASEP) результатов может быть восстановлен из этой модели, что делает энергию равной нулю.

Митоз
[ редактировать ]В последние годы было обнаружено, что молекулярные двигатели на основе микротрубочек (включая ряд кинезинов) играют роль в митозе (деление клеток). Кинеины важны для правильной длины веретена и участвуют в скольжении микротрубочек, отделенных внутри шпинделя во время прометафазы и метафазы, а также деполимеризационные микротрубочки минус заканчиваются в центросомах во время анафазы. [ 53 ] В частности, белки семейства кинезин-5 действуют в шпинделе, чтобы раздвигать микротрубочки, в то время как семейство Кинезина 13 действует для деполимеризации микротрубочек.
Кинезин суперсемейство
[ редактировать ]Члены суперсемейства человека включают следующие белки, которые в стандартизированной номенклатуре, разработанной сообществом исследователей кинезина, организованы в 14 семей с названием кинезин-1 через кинезин-14: [ 12 ]
- 1A- KIF1A , 1B- KIF1B , 1C- KIF1C = Kinesin-3
- 2A- KIF2A , 2C- KIF2C = Kinesin-13
- 3B - KIF3B или 3C - KIF3C , 3A - KIF3A = Kinesin -2
- 4A- KIF4A , 4B- KIF4B = Kinesin-4
- 5A- KIF5A , 5B- KIF5B , 5C- KIF5C = KINESIN-1
- 6- KIF6 = кинезин-9
- 7- KIF7 = кинезин-4
- 9- KIF9 = кинезин-9
- 11- KIF11 = Kinesin-5
- 12- KIF12 = кинезин-12
- 13A- KIF13A , 13B- KIF13B = Kinesin-3
- 14- KIF14 = кинезин-3
- 15- KIF15 = кинезин-12
- 16b- KIF16B = Kinesin-3
- 17- KIF17 = кинезин-2
- 18A- KIF18A , 18B- KIF18B = Kinesin-8
- 19- KIF19 = кинезин-8
- 20A- KIF20A , 20B- KIF20B = Kinesin-6
- 21a- KIF21A , 21b- KIF21b = Kinesin-4
- 22- KIF22 = кинезин-10
- 23- KIF23 = кинезин-6
- 24- KIF24 = кинезин-13
- 25- KIF25 = кинезин-14
- 26A- KIF26A , 26B- KIF26B = Kinesin-11
- 27- KIF27 = кинезин-4
- C1- KIFC1 , C2- KIFC2 , C3- KIFC3 = Kinesin-14
Световые цепи кинезина-1:
Кинезин-2-ассоциированный белок:
- Kifap3 (также известный как KAP-1, KAP3)
Смотрите также
[ редактировать ]- Аксональный транспорт
- Динамин
- Внутрикафлагелентный транспорт вдоль ресничек
- Кинезин 8
- Кинезин 13
- КРП
- Молекулярный мотор
- Транспорт с помощью нескольких моторных белков
Ссылки
[ редактировать ]- ^ Berg J, Tymoczko JL, Stryer L (2002). «Кинезин и динеин перемещаются по микротрубочкам» . Биохимия. 5 -е издание .
- ^ Vale Rd (февраль 2003 г.). «Молекулярный моторный набор инструментов для внутриклеточного транспорта» . Клетка . 112 (4): 467–80. doi : 10.1016/s0092-8674 (03) 00111-9 . PMID 12600311 . S2CID 15100327 .
- ^ Endow SA, Kull FJ, Liu H (октябрь 2010 г.). «Кинеры с первого взгляда» . Журнал сотовой науки . 123 (Pt 20): 3420–4. doi : 10.1242/jcs.064113 . PMC 2951464 . PMID 20930137 .
- ^ Vale Rd, Reese TS, Sheetz MP (август 1985 г.). «Идентификация нового, генерирующего силы белка, кинезина, участвующего в подвижности на основе микротрубочек» . Клетка . 42 (1): 39–50. doi : 10.1016/s0092-8674 (85) 80099-4 . PMC 2851632 . PMID 3926325 .
- ^ Cole DG, Chinn SW, Wedaman KP, Hall K, Vuong T, Scholey JM (ноябрь 1993). «Новый гетеротримерный кинезиновый белок, очищенный от яиц морских ежей». Природа . 366 (6452): 268–70. Bibcode : 1993natur.366..268c . doi : 10.1038/366268A0 . PMID 8232586 . S2CID 4367715 .
- ^ Розенбаум JL, Witman GB (ноябрь 2002 г.). «Внутренний транспорт». Природные обзоры. Молекулярная клеточная биология . 3 (11): 813–25. doi : 10.1038/nrm952 . PMID 12415299 . S2CID 12130216 .
- ^ Ян Д.Т., Леймон Р.А., Гольдштейн Л.С. (март 1989 г.). «Трехдоменная структура тяжелой цепи кинезина, выявленная последовательности ДНК и анализа связывания микротрубочек». Клетка . 56 (5): 879–89. doi : 10.1016/0092-8674 (89) 90692-2 . PMID 2522352 . S2CID 44318695 .
- ^ Aizawa H, Sekine Y, Takemura R, Zhang Z, Nangaku M, Hirokawa N (декабрь 1992 г.). «Семейство кинезин в мышиной центральной нервной системе» . Журнал клеточной биологии . 119 (5): 1287–96. doi : 10.1083/jcb.119.5.1287 . PMC 2289715 . PMID 1447303 .
- ^ ENOS AP, Morris NR (март 1990 г.). «Мутация гена, который кодирует кинезиноподобное белок, блокирует ядерное деление у A. nidulans». Клетка . 60 (6): 1019–27. doi : 10.1016/0092-8674 (90) 90350-N . PMID 2138511 . S2CID 27420513 .
- ^ Meluh PB, Rose MD (март 1990 г.). «KAR3, связанный с кинезином ген, необходимый для ядерного слияния дрожжей». Клетка . 60 (6): 1029–41. doi : 10.1016/0092-8674 (90) 90351-E . PMID 2138512 . S2CID 19660190 .
- ^ Hirokawa N, Noda Y, Tanaka Y, Niwa S (октябрь 2009 г.). «Моторные белки суперсемейства кинезина и внутриклеточный транспорт». Природные обзоры. Молекулярная клеточная биология . 10 (10): 682–96. doi : 10.1038/nrm2774 . PMID 19773780 . S2CID 18129292 .
- ^ Jump up to: а беременный Лоуренс К.Дж., Доу Р.К., Кристи К.Р., Кливленд Д.В., Доусон С.К., Эндоу С.А., Гольдштейн Л.С., Гудсон Х.В., Хирокава Н., Ховард Дж., Мальмберг Р.Л., Макинтош младший, Мики Х, Митчисон Т.Дж., Окада Й, Редди А.С. , Schliwa M, Scholey JM, Vale Rd, Walczak CE, Wordeman L (октябрь 2004 г.). «Стандартизированная номенклатура кинезина» . Журнал клеточной биологии . 167 (1): 19–22. doi : 10.1083/jcb.200408113 . PMC 2041940 . PMID 15479732 .
- ^ Hirokawa N, Pfister KK, Yorifuji H, Wagner MC, Brady St, Bloom GS (март 1989 г.). «Судолекулярные домены кинезина бычьего мозга, идентифицированных с помощью электронной микроскопии и моноклонального антитела». Клетка . 56 (5): 867–78. doi : 10.1016/0092-8674 (89) 90691-0 . PMID 2522351 . S2CID 731898 .
- ^ ВВП : 1BG2 ; Kull FJ, Sablin EP, Lau R, Fletterick RJ, Vale Rd (апрель 1996 г.). «Кристаллическая структура кинезинового моторного домена выявляет структурное сходство с миозином» . Природа . 380 (6574): 550–5. Bibcode : 1996natur.380..550J . doi : 10.1038/380550A0 . PMC 2851642 . PMID 8606779 .
- ^ Jump up to: а беременный Kaan HY, Hackney DD, Kozielski F (август 2011 г.). «Структура комплекса двигателя кинезина-1 выявляет механизм аутоингибирования» . Наука . 333 (6044): 883–5. Bibcode : 2011sci ... 333..883k . doi : 10.1126/science.1204824 . PMC 3339660 . PMID 21836017 .
- ^ Стюарт Р.Дж., Талер Дж.П., Гольдштейн Л.С. (июнь 1993 г.). «Направление движения микротрубочек является внутренним свойством моторных доменов кинезиновой тяжелой цепи и белка Drosophila NCD» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (11): 5209–13. Bibcode : 1993pnas ... 90.5209S . doi : 10.1073/pnas.90.11.5209 . PMC 46685 . PMID 8506368 .
- ^ Verhey KJ, Hammond JW (ноябрь 2009 г.). «Управление движением: регулирование кинезиновых двигателей». Природные обзоры. Молекулярная клеточная биология . 10 (11): 765–77. doi : 10.1038/nrm2782 . PMID 19851335 . S2CID 10713993 .
- ^ Siddiqui N, Zwetsloot AJ, Bachmann A, Roth D, Hussain H, Brandt J, et al. (Июнь 2019). «PTPN21 и Hook3 облегчают аутоингибирование KIF1C и активируют внутриклеточный транспорт» . Природная связь . 10 (1): 2693. Bibcode : 2019natco..10.2693S . doi : 10.1038/s41467-019-10644-9 . PMC 6584639 . PMID 31217419 .
- ^ Blasius TL, Cai D, Jih GT, Toret CP, Verhey KJ (январь 2007 г.). «Два партнера по связыванию сотрудничают, чтобы активировать молекулярный моторный кинезин-1» . Журнал клеточной биологии . 176 (1): 11–7. doi : 10.1083/jcb.200605099 . PMC 2063617 . PMID 17200414 .
- ^ Hooikaas PJ, Martin M, Mühlethaler T, Kuijntjes GJ, Peeters CA, Kathukha EA, et al. (Апрель 2019). «Семейные белки MAP7 регулируют рекрутирование и активацию кинезина-1» . Журнал клеточной биологии . 218 (4): 1298–1318. doi : 10.1083/jcb.201808065 . PMC 6446838 . PMID 30770434 .
- ^ Schnitzer MJ, Block SM (июль 1997 г.). «Кинеиновые гидролизы Один АТФ на шаг 8 нм» . Природа . 388 (6640): 386–90. Bibcode : 1997natur.388..386s . doi : 10.1038/41111 . PMID 9237757 . S2CID 4363000 .
- ^ Vale Rd, Milligan RA (апрель 2000 г.). «То, как все движется: глядя под капот молекулярных моторных белков». Наука . 288 (5463): 88–95. Bibcode : 2000sci ... 288 ... 88V . doi : 10.1126/science.288.5463.88 . PMID 10753125 .
- ^ Mather WH, Fox RF (октябрь 2006 г.). «Предвзятый механизм ступени Кинезина: амплификация линкера на молнии шеи» . Биофизический журнал . 91 (7): 2416–26. Bibcode : 2006bpj .... 91.2416m . doi : 10.1529/biophysj.106.087049 . PMC 1562392 . PMID 16844749 .
- ^ Гаудин Р., де Аленгар Б.К., Жуве М, Берре С., Ле Будер Э., Шиндлер М., Вартаман А., Гоберт Ф.Х., Бенарох П (октябрь 2012 г.). «Критическая роль кинезина KIF3A в жизненном цикле ВИЧ в первичных человеческих макрофагах» . Журнал клеточной биологии . 199 (3): 467–79. doi : 10.1083/jcb.201201144 . PMC 3483138 . PMID 23091068 .
- ^ Gross SP, Vershinin M, Shubeita GT (июнь 2007 г.). «Транспорт с грузом: два двигателя иногда лучше одного» . Текущая биология . 17 (12): R478–86. doi : 10.1016/j.cub.2007.04.025 . PMID 17580082 . S2CID 8791125 .
- ^ Hancock WO (август 2008 г.). «Внутриклеточный транспорт: кинезины работают вместе» . Текущая биология . 18 (16): R715–7. doi : 10.1016/j.cub.2008.07.068 . PMID 18727910 . S2CID 7540556 .
- ^ Кунвар А., Вершинин М., Сюй Дж., Гросс С.П. (август 2008 г.). «Stepping, Drawing gating и неожиданная кривая силы-скорости для транспорта на основе нескольких мотор» . Текущая биология . 18 (16): 1173–83. doi : 10.1016/j.cub.2008.07.027 . PMC 33855514 . PMID 18701289 .
- ^ Klumpp S, Lipowsky R (ноябрь 2005 г.). «Кооперативный транспорт грузов по нескольким молекулярным двигателям» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (48): 17284–9. arxiv : Q-bio/0512011 . Bibcode : 2005pnas..10217284K . doi : 10.1073/pnas.0507363102 . PMC 1283533 . PMID 16287974 .
- ^ Rice S, Lin AW, SAFER D, Hart CL, Naber N, Carragher Bo, Cain SM, Pechatnikova E, Wilson-Kubalek EM, Whittaker M, Pate E, Cooke R, Taylor EW, Milligan RA, Vale Rd (декабрь 1999 г.) Полем «Структурное изменение в кинезинском моторном белке, который приводит к подвижности». Природа . 402 (6763): 778–84. Bibcode : 1999natur.402..778r . doi : 10.1038/45483 . PMID 10617199 . S2CID 573909 .
- ^ Lodish H, Berk A, Zipursky SL, Matsudaira P, Baltimore D, Darnell J (2000). «Кинезин, Динеин и внутриклеточный транспорт» .
{{cite journal}}: CITE Journal требует|journal=( помощь ) - ^ Roostalu J, Hentrich C, Bieling P, Telley IA, Schiebel E, Surrey T (апрель 2011 г.). «Направленное переключение кинезина CIN8 через моторную связь» . Наука . 332 (6025): 94–9. Bibcode : 2011sci ... 332 ... 94r . doi : 10.1126/science.1199945 . PMID 21350123 . S2CID 90739364 .
- ^ Fallesen T, Roostalu J, Duellberg C, Pruessner G, Surrey T (ноябрь 2017 г.). «Ансамбли двунаправленного кинезина CIN8 производят аддитивные силы в обоих направлениях движения» . Биофизический журнал . 113 (9): 2055–2067. Bibcode : 2017bpj ... 113.2055f . doi : 10.1016/j.bpj.2017.09.006 . PMC 5685778 . PMID 29117528 .
- ^ Edamatsu M (март 2014 г.). «Двунаправленная подвижность делящихся дрожжевых кинезин-5, cut7» . Биохимическая и биофизическая исследовательская коммуникация . 446 (1): 231–4. doi : 10.1016/j.bbrc.2014.02.106 . PMID 24589736 .
- ^ Roostalu J, Hentrich C, Bieling P, Telley IA, Schiebel E, Surrey T (апрель 2011 г.). «Направленное переключение кинезина CIN8 через моторную связь». Наука . 332 (6025): 94–9. Bibcode : 2011sci ... 332 ... 94r . doi : 10.1126/science.1199945 . PMID 21350123 . S2CID 90739364 .
- ^ Jump up to: а беременный Gerson-Gurwitz A, Thiede C, Movshovich N, Fridman V, Podolskaya M, Danieli T, et al. (Ноябрь 2011). «Направленность отдельных двигателей кинезина-5 CIN8 модулируется петлей 8, ионной прочности и геометрией микротрубочек» . Embo Journal . 30 (24): 4942–54. doi : 10.1038/emboj.2011.403 . PMC 3243633 . PMID 22101328 .
- ^ Валентин М.Т., Фордис П.М., Блок С.М. (декабрь 2006 г.). "EG5 снимает это!" Полем Клеточная деление . 1 (1): 31. doi : 10.1186/1747-1028-1-31 . PMC 1716758 . PMID 17173688 .
- ^ Ambrose JC, Li W, Marcus A, Ma H, Cyr R (апрель 2005 г.). «Учебный минус, направленный в энду, с активностью белка плюс-элитного белка участвует в морфогенезе шпинделя» . Молекулярная биология клетки . 16 (4): 1584–92. doi : 10.1091/mbc.e04-10-0935 . PMC 1073643 . PMID 15659646 .
- ^ Фэй, Джинью; Чжоу, Руобу (10 марта 2023 г.). «Наблюдение за биомолекулами шагает в режиме реального времени» . Наука . 379 (6636): 986–987. doi : 10.1126/science.adg8451 . PMC 10318587 . PMID 36893224 .
- ^ Дегучи, Такахиро (10 марта 2023 г.). «Прямое наблюдение за шагом моторного белка в живых клетках с использованием minflux» . Наука . 379 (6636): 1010–1015. doi : 10.1126/science.ade2676 . PMC 7614483 . PMID 36893247 .
- ^ Вольф, Ян; Scheiderer, Lukas; Энгельхардт, Тобиас; Энгельхардт, Иоганн; Матиас, Джессика; Ад, Стефан (10 марта 2023 г.). «Minflux анализирует беспрепятственную ходьбу кинезина-1». Наука . 379 (6636): 1004–1010. doi : 10.1126/science.ade2650 . PMID 36893244 . S2CID 251162014 .
- ^ Yildiz A, Tomishige M, Vale Rd, Selvin PR (январь 2004 г.). «Кинезин ходит вручную». Наука . 303 (5658): 676–8. Bibcode : 2004sci ... 303..676y . doi : 10.1126/science.1093753 . PMID 14684828 . S2CID 30529199 .
- ^ Эсбери Кл (февраль 2005 г.). «Кинезин: самый крошечный в мире двуногнутый». Современное мнение в клеточной биологии . 17 (1): 89–97. doi : 10.1016/j.ceb.2004.12.002 . PMID 15661524 .
- ^ Sindelar CV, Даунинг KH (март 2010 г.). «Механизм атомного уровня для активации кинезиновых молекулярных двигателей» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (9): 4111–6. Bibcode : 2010pnas..107.4111S . doi : 10.1073/pnas.0911208107 . PMC 2840164 . PMID 20160108 .
- ^ Сводка мирян (18 февраля 2010 г.). «Самый маленький двигатель жизни, грузовой носитель клеток, движется как качели» . Physorgg.com . Получено 31 мая 2013 года .
- ^ Atzberger PJ, Peskin CS (январь 2006 г.). «Модель кинезина с коричневой динамикой в трех измерениях, включающая профиль силовой экстенсии в спиральной камере грузового привязки». Бюллетень математической биологии . 68 (1): 131–60. Arxiv : 0910.5753 . doi : 10.1007/s11538-005-9003-6 . PMID 16794924 . S2CID 13534734 .
- ^ Peskin CS, Oster G (апрель 1995 г.). «Координированный гидролиз объясняет механическое поведение кинезина» . Биофизический журнал . 68 (4 Suppl): 202S - 210S, обсуждение 210 -х - 211 с. PMC 1281917 . PMID 7787069 .
- ^ Mogilner A, Fisher AJ, Baskin RJ (июль 2001 г.). «Структурные изменения в линкере кинезина в шее объясняют зависимость нагрузки механического цикла двигателя». Журнал теоретической биологии . 211 (2): 143–57. Bibcode : 2001jthbi.211..143m . doi : 10.1006/jtbi.2001.2336 . PMID 11419956 .
- ^ Jump up to: а беременный Celis-Garza D, Teimouri H, Kolomeisky AB (2015). «Корреляции и симметрия взаимодействий влияют на коллективную динамику молекулярных двигателей». Журнал статистической механики: теория и эксперимент . 2015 (4): P04013. Arxiv : 1503.00633 . Bibcode : 2015jsmte..04..013c . doi : 10.1088/1742-5468/2015/04/p04013 . S2CID 14002728 .
- ^ Jump up to: а беременный Липельт, Штеффен; Липоуски, Рейнхард (20 июня 2007 г.). «Сеть химических моторных циклов Кинезина» . Письма о физическом обзоре . 98 (25): 258102. Bibcode : 2007 Phrvl..98y8102L . doi : 10.1103/physrevlett.98.258102 . PMID 17678059 .
- ^ Сейферт, Дэвид; Соллих, Питер; Клумпп, Стефан (29 декабря 2020 г.). «Грубое зернистые биохимические системы, описанные дискретной стохастической динамикой» . Физический обзор e . 102 (6): 062149. Arxiv : 2102.13394 . BIBCODE : 2020PHRVE.102F2149S . doi : 10.1103/physreve.102.062149 . PMID 33466014 . S2CID 231652939 .
- ^ Seitz A, Surrey T (январь 2006 г.). «Процедурное движение отдельных кинезинов на переполненных микротрубочках, визуализируемых с использованием квантовых точек» . Embo Journal . 25 (2): 267–77. doi : 10.1038/sj.emboj.7600937 . PMC 1383520 . PMID 16407972 .
- ^ Вильфан А., Фрей Е., Швабл Ф., Тормален М., Сонг Й.Х., Мандельков Е (октябрь 2001 г.). «Динамика и кооперативность украшения микротрубочек моторным белком кинезином». Журнал молекулярной биологии . 312 (5): 1011–26. doi : 10.1006/jmbi.2001.5020 . PMID 11580246 .
- ^ Госима Г., Вейл Р.Д. (август 2005 г.). «Зависимая от клеточного цикла динамика и регуляция митотических кинезинов в клетках Drosophila S2» . Молекулярная биология клетки . 16 (8): 3896–907. doi : 10.1091/mbc.e05-02-0118 . PMC 1182325 . PMID 15958489 .
Дальнейшее чтение
[ редактировать ]- Лоуренс К.Дж., Доу Р.К., Кристи К.Р., Кливленд Д.В., Доусон С.К., Эндоу С.А., Гольдштейн Л.С., Гудсон Х.В., Хирокава Н., Ховард Дж., Мальмберг Р.Л., Макинтош младший, Мики Х, Митчисон Т.Дж., Окада Й, Редди А.С. , Schliwa M, Scholey JM, Vale Rd, Walczak CE, Wordeman L (октябрь 2004 г.). «Стандартизированная номенклатура кинезина» . Журнал клеточной биологии . 167 (1): 19–22. doi : 10.1083/jcb.200408113 . PMC 2041940 . PMID 15479732 .
Внешние ссылки
[ редактировать ]- Mbinfo - кинезиновый транспортирует грузы вдоль микротрубочек
- Анимированная модель ходьбы кинезина
- Семинар Рона Вейла: «Молекулярные моторные белки»
- Анимация кинезинного движения ASCB библиотеки изображений
- Мерфи, VF (12 мая 2004 г.). «Движение на основе микротрубочек» . ткань.medicalengineer.co.uk . Архивировано из оригинала 22 июля 2007 года . Получено 10 декабря 2015 года .
- Внутренняя жизнь ячейки, 3D -анимация с кинезином, транспортирующим везикул, архивированный 7 декабря 2008 года на машине Wayback
- КИНЕСИНА ДОМАНА
- Кинезин в Национальной библиотеке медицинской библиотеки США медицинские заголовки (сетка)
- ЕС 3.6.4.4
- ЕС 3.6.4.5
- 3D -электронная микроскопическая структура кинезина из банка данных EM (EMDB)
| Базы данных управления авторитетом : национальный |
|---|