Хеликаза синдрома Вернера
| WRN | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | WRN , RECQ3, RECQL2, RECQL3, синдром Вернера RecQ-подобная геликаза, WRN RecQ-подобная геликаза | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 604611 ; МГИ : 109635 ; Гомологен : 6659 ; GeneCards : WRN ; ОМА : WRN – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Номер ЕС | 3.1.-.- | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Синдром Вернера АТФ-зависимая геликаза , также известная как ДНК-геликаза, RecQ-подобный тип 3 , представляет собой фермент , который у человека кодируется WRN геном . WRN является членом семейства RecQ Helicase . [ 5 ] Ферменты хеликазы обычно раскручивают и разделяют двухцепочечную ДНК . Эти действия необходимы до того, как ДНК сможет быть скопирована при подготовке к делению клеток ( репликация ДНК ). Ферменты хеликазы также имеют решающее значение для создания проекта гена для производства белка — процесса, называемого транскрипцией . Дальнейшие данные свидетельствуют о том, что белок Вернера играет решающую роль в восстановлении ДНК . В целом этот белок помогает поддерживать структуру и целостность ДНК человека.
Ген WRN расположен на коротком (p) плече хромосомы 8 между положениями 12 и 11,2, от пары оснований 31 010 319 до пары оснований 31 150 818.
Структура и функции
[ редактировать ]WRN является членом семейства RecQ Helicase . Это единственная геликаза RecQ, которая содержит экзонуклеазную активность от 3' до 5'. Эта экзонуклеазная активность включает деградацию утопленных 3'-концов и инициацию деградации ДНК из-за разрыва в дцДНК. WRN важен для восстановления двухцепочечных разрывов путем гомологичной рекомбинации. [ 6 ] [ 7 ] или негомологическое соединение концов , [ 8 ] восстановление одиночных нуклеотидных повреждений путем эксцизионной репарации оснований , [ 9 ] [ 10 ] [ 5 ] и эффективен при восстановлении после остановки репликации. [ 11 ] WRN также может быть важен для поддержания и репликации теломер, особенно репликации G-богатых последовательностей. [ 12 ]
WRN представляет собой олигомер , который может действовать как мономер при раскручивании ДНК, но как димер в растворе или как тетрамер при образовании комплекса с ДНК, а также наблюдался в гексамерных формах. Диффузия WRN была измерена как 1,62. в нуклеоплазме и 0,12 у ядрышек. [ 13 ] Ортологи WRN были обнаружены у ряда других организмов, включая Drosophila , Xenopus и C. elegans . WRN важен для стабильности генома, а клетки с мутациями WRN более восприимчивы к повреждениям ДНК и разрывам ДНК. [ 14 ]


Аминоконец , WRN участвует как в активности геликазы так и в активности нуклеазы , тогда как карбоксильный конец взаимодействует с р53 , важным супрессором опухоли. [ 15 ] WRN может действовать как экзонуклеаза при репарации, рекомбинации или репликации ДНК, а также при разрешении вторичных структур ДНК. Он участвует в миграции ветвей в соединениях Холлидея и взаимодействует с другими промежуточными продуктами репликации ДНК. [ 11 ] мРНК, кодирующая WRN, была идентифицирована в большинстве тканей человека. [ 15 ]
Посттрансляционная модификация
[ редактировать ]Фосфорилирование WRN по серину/треонину ингибирует активность геликазы и экзонуклеазы, которые важны для пострепликационной репарации ДНК. Дефосфорилирование этих сайтов усиливает каталитическую активность WRN. Фосфорилирование может влиять на другие посттрансляционные модификации, включая сумойилирование и ацетилирование. [ 12 ]
Метилирование WRN приводит к отключению гена. Это подавляет выработку белка WRN и его функции в репарации ДНК. [ 16 ]
Клиническое значение
[ редактировать ]Синдром Вернера вызван мутациями в гене WRN . [ 15 ] Известно, что более 20 мутаций в гене WRN вызывают синдром Вернера. Многие из этих мутаций приводят к аномально укороченному белку Вернера. Имеющиеся данные свидетельствуют о том, что измененный белок не транспортируется в ядро клетки , где он обычно взаимодействует с ДНК. [ 17 ] Этот укороченный белок также может расщепляться слишком быстро, что приводит к потере белка Вернера в клетке. Без нормального белка Вернера в ядре клетки не могут выполнять задачи репликации, репарации и транскрипции ДНК. [ 18 ] Исследователи все еще выясняют, как эти мутации вызывают преждевременное старение, наблюдаемое при синдроме Вернера.
Роль в путях восстановления ДНК
[ редактировать ]Гомологичная рекомбинационная репарация
[ редактировать ]WRN активен в гомологичной рекомбинации . Клетки, дефектные по гену WRN , имеют 23-кратное снижение спонтанной митотической рекомбинации с особенным дефицитом событий конверсионного типа. [ 19 ] Дефектные WRN клетки при воздействии рентгеновских лучей имеют больше хромосомных разрывов и микроядер, чем клетки с WRN дикого типа. [ 20 ] Клетки, дефектные по гену WRN , не более чувствительны, чем клетки дикого типа, к гамма-облучению, УФ-свету, 4–6 циклобутанпиримидинам или митомицину С, но чувствительны к ингибиторам топоизомеразы I и II типов. [ 21 ] Эти данные позволяют предположить, что белок WRN принимает участие в гомологичной рекомбинационной репарации и в обработке остановившихся репликационных вилок. [ 22 ]
негомологичное соединение концов
[ редактировать ]WRN играет важную роль в репарации ДНК негомологичного соединения концов (NHEJ). Как показали Шаманна и др., [ 8 ] WRN рекрутируется в двухцепочечные разрывы (DSB) и участвует в NHEJ, выполняя ферментативные и неферментативные функции. В DSB в сочетании с Ku (белком) он способствует стандартному или каноническому NHEJ (c-NHEJ), восстанавливая двухцепочечные разрывы в ДНК своими ферментативными функциями и с достаточной степенью точности. WRN ингибирует альтернативную форму NHEJ, называемую alt-NHEJ или микрогомологическое соединение концов (MMEJ). MMEJ — это неточный способ восстановления двухцепочечных разрывов.
Базовый иссеченный ремонт
[ редактировать ]WRN играет роль в эксцизионной репарации оснований (BER) ДНК. Как показали Дас и др., [ 9 ] WRN взаимодействует с NEIL1 на раннем этапе BER по обнаружению повреждений. WRN стимулирует NEIL1 при удалении окислительных поражений. NEIL1 представляет собой ДНК-гликозилазу , которая инициирует первый этап BER путем расщепления оснований, поврежденных активными формами кислорода (АФК), и создания разрыва цепи ДНК посредством связанной с NEIL1 лиазной активности. [ 23 ] NEIL1 распознает (нацеливается) и удаляет определенные ROS- поврежденные основания, а затем разрезает абазический сайт путем β,δ-элиминирования, оставляя 3'- и 5'-фосфатные концы. NEIL1 распознает окисленные пиримидины , формамидопиримидины, остатки тимина , окисленные по метильной группе, и оба стереоизомера тимингликоля . [ 24 ]
WRN также участвует в BER посредством взаимодействия с Polλ . [ 10 ] WRN связывается с каталитическим доменом Polλ и специфически стимулирует заполнение пробелов в ДНК с помощью Polλ над 8-оксо-G с последующим синтезом замещения цепи. Это позволяет WRN стимулировать синтез репарации ДНК с длинными участками с помощью Polλ во время MUTYH -инициируемой репарации неправильных пар 8-оксо-G:A.
Восстановление после остановки репликации
[ редактировать ]WRN также участвует в восстановлении после остановки репликации. Если WRN дефектен, остановка репликации приводит к накоплению DSB и усилению фрагментации хромосом. [ 25 ] Как показали Пикьерри и др., [ 25 ] WRN взаимодействует с комплексом RAD9 - RAD1 - HUS1 (9.1.1), одним из центральных факторов контрольной точки репликации. Это взаимодействие опосредовано связыванием субъединицы RAD1 с N-концевой областью WRN и играет важную роль в релокализации WRN в ядерные очаги и его фосфорилировании в ответ на арест репликации. (При отсутствии повреждения ДНК или остановки репликационной вилки белок WRN остается локализованным в ядрышках. [ 26 ] ) Взаимодействие WRN с комплексом 9.1.1 приводит к предотвращению образования DSB на остановившихся репликационных вилках. [ 25 ]
Роль в апоптозе
[ редактировать ]Белок p53 и хеликаза WRN участвуют в прямом белок-белковом взаимодействии. [ 27 ] Повышенные клеточные уровни WRN вызывают повышение клеточных уровней p53, а также потенцируют p53-опосредованный апоптоз . [ 27 ] Это открытие предполагает, что WRN-хеликаза участвует в активации р53 в ответ на определенные типы повреждений ДНК . [ 27 ] p53-опосредованный апоптоз ослаблен в клетках пациентов с синдромом Вернера. [ 28 ]
И восстановление повреждений ДНК , и апоптоз являются ферментативными процессами, необходимыми для поддержания целостности генома человека . Клетки с недостаточной репарацией ДНК имеют тенденцию накапливать повреждения ДНК, а когда такие клетки также имеют дефекты апоптоза, они имеют тенденцию выживать, даже если присутствуют чрезмерные повреждения ДНК. [ 29 ] Репликация ДНК в таких дефектных клетках имеет тенденцию приводить к мутациям , а такие мутации могут вызывать рак. Таким образом, геликаза синдрома Вернера, по-видимому, выполняет две роли, связанные с профилактикой рака: первая роль заключается в содействии восстановлению определенных типов повреждений, а вторая роль заключается в индукции апоптоза, если уровень такого повреждения ДНК выходит за пределы способности клетки к восстановлению. [ 29 ]
Дефицит WRN при раке
[ редактировать ]Клетки, экспрессирующие ограниченное количество WRN, имеют повышенную частоту мутаций по сравнению с клетками дикого типа. [ 30 ] Повышенная мутация может привести к раку. Пациенты с синдромом Вернера с гомозиготными мутациями в гене WRN имеют повышенную заболеваемость раком, включая саркомы мягких тканей, остеосаркому, рак щитовидной железы и меланому. [ 31 ]
Мутации WRN в общей популяции встречаются редко. Частота гетерозиготных мутаций с потерей функции при WRN составляет примерно одну на миллион. В Японии этот показатель составляет 6 на 1000, что выше, но все же встречается нечасто. [ 32 ]
Мутационные дефекты гена WRN относительно редки в раковых клетках по сравнению с частотой эпигенетических изменений в WRN , которые снижают экспрессию WRN и могут способствовать канцерогенезу. Ситуация аналогична другим генам репарации ДНК, экспрессия которых снижается при раке главным образом из-за эпигенетических изменений, а не мутаций (см. Частоты эпимутаций в генах репарации ДНК ). [ нужна ссылка ]
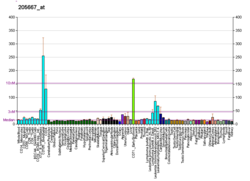
В таблице представлены результаты анализа 630 первичных опухолей человека на гиперметилирование CpG-островков WRN . [ 33 ] Это гиперметилирование вызывало снижение экспрессии белка WRN, что является обычным явлением при онкогенезе. [ 33 ]
| Рак | Частота снижения заболеваемости раком [ 33 ] |
|---|---|
| Колоректальный рак | 37.9% |
| Немелкоклеточный рак легкого | 37.5% |
| Рак желудка | 25% |
| Рак простаты | 20% |
| Рак молочной железы | 17.2% |
| Рак щитовидной железы | 12.5% |
| Неходжкинская лимфома | 23.7% |
| Острый миелобластный лейкоз | 4.8% |
| Хондросаркомы | 33.3% |
| Остеосаркомы | 11.1% |
Взаимодействия
[ редактировать ]Было показано, что АТФ-зависимая геликаза синдрома Вернера взаимодействует с:
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000165392 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000031583 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Перейти обратно: а б Моннат Р.Ж. (октябрь 2010 г.). «Человеческие RECQ-хеликазы: роль в метаболизме ДНК, мутагенезе и биологии рака» . Семин. Рак Биол . 20 (5): 329–39. дои : 10.1016/j.semcancer.2010.10.002 . ПМК 3040982 . ПМИД 20934517 .
- ^ Сентиньи И., Макиенко К., Суонсон С., Эмонд М.Дж., Моннат Р.Дж. (2002). «Дефект разрешения гомологичной рекомбинации при синдроме Вернера» . Мол. Клетка. Биол . 22 (20): 6971–8. дои : 10.1128/mcb.22.20.6971-6978.2002 . ПМК 139822 . ПМИД 12242278 .
- ^ Стурценеггер А, Бурдова К, Канагарадж Р, Левикова М, Пинто С, Чейка П, Янскак П (2014). «ДНК2 взаимодействует с хеликазами WRN и BLM RecQ, обеспечивая резекцию концов ДНК на большие расстояния в клетках человека» . Ж. Биол. Хим . 289 (39): 27314–26. дои : 10.1074/jbc.M114.578823 . ПМЦ 4175362 . ПМИД 25122754 .
- ^ Перейти обратно: а б Шаманна Р.А., Лу Х., де Фрейтас Дж.К., Тиан Дж., Крото Д.Л., Бор В.А. (2016). «WRN регулирует выбор пути между классическим и альтернативным негомологичным соединением концов» . Нат Коммун . 7 : 13785. Бибкод : 2016NatCo...713785S . дои : 10.1038/ncomms13785 . ПМК 5150655 . ПМИД 27922005 .
- ^ Перейти обратно: а б Дас А., Болдог И., Ли Дж.В., Харриган Дж.А., Хегде М.Л., Пиотровски Дж., де Соуза Пинто Н., Рамос В., Гринберг М.М., Хазра Т.К., Митра С., Бор В.А. (2007). «Белок синдрома Вернера человека стимулирует восстановление окислительных повреждений оснований ДНК с помощью ДНК-гликозилазы NEIL1» . Ж. Биол. Хим . 282 (36): 26591–602. дои : 10.1074/jbc.M703343200 . ПМИД 17611195 .
- ^ Перейти обратно: а б Канагарадж Р., Парасураман П., Михалевич Б., ван Лун Б., Бурдова К., Кениг С., Фуррер А., Бор В.А., Хюбшер У., Янскак П. (2012). «Участие белка синдрома Вернера в MUTYH-опосредованном восстановлении окислительных повреждений ДНК» . Нуклеиновые кислоты Рез . 40 (17): 8449–59. дои : 10.1093/nar/gks648 . ПМЦ 3458577 . ПМИД 22753033 .
- ^ Перейти обратно: а б Пикьерри П., Аммаззалорсо Ф., Биньями М., Франчитто А. (2011). «Белок синдрома Вернера: связь реакции контрольной точки репликации со стабильностью генома» . Старение . 3 (3): 311–8. дои : 10.18632/aging.100293 . ПМК 3091524 . ПМИД 21389352 .
- ^ Перейти обратно: а б Дин С.Л., Шен С.И. (2008). «Модель старения человека: последние данные о синдромах прогерии Вернера и Хатчинсона-Гилфорда» . Клин Интервальное старение . 3 (3): 431–44. дои : 10.2147/CIA.S1957 . ПМЦ 2682376 . ПМИД 18982914 .
- ^ Бендцен К.М., Йенсен М.Б., Мэй А., Расмуссен Л.Дж., Трусина А., Бор В.А., Йенсен М.Х. (2014). «Динамика белков репарации ДНК WRN и BLM в нуклеоплазме и ядрышках» . Европейский биофизический журнал . 43 (10–11): 509–16. дои : 10.1007/s00249-014-0981-x . ПМЦ 5576897 . ПМИД 25119658 .
- ^ Росси М.Л., Гош А.К., Бор В.А. (2010). «Роль белка синдрома Вернера в защите целостности генома» . Восстановление ДНК (Амст.) . 9 (3): 331–44. дои : 10.1016/j.dnarep.2009.12.011 . ПМЦ 2827637 . ПМИД 20075015 .
- ^ Перейти обратно: а б с Осима Дж (2000). «Белок синдрома Вернера: обновление». Биоэссе . 22 (10): 894–901. doi : 10.1002/1521-1878(200010)22:10<894::AID-BIES4>3.0.CO;2-B . ПМИД 10984715 . S2CID 36746466 .
- ^ «ВРН» . Национальная медицинская библиотека США . Проверено 18 марта 2014 г.
- ^ Хуан С., Ли Л., Хансон Н.Б., Ленартс С., Хоэн Х., Пут М., Рубин К.Д., Чен Д.Ф., Ян CC, Джух Х., Дорн Т., Шпигель Р., Орал Э.А., Абид М., Баттисти С., Луччи-Кордиско Е, Нери Дж., Стид Э.Х., Кидд А., Айсли В., Шоуолтер Д., Виттоне Дж.Л., Константинов А., Ринг Дж., Мейер П., Венгер С.Л., фон Хербей А., Воллина У., Шуэльке М., Хуйзенга Ч.Р., Лейстриц Д.Ф., Мартин Г.М., Миан И.С., Осима Дж (2006). «Спектр мутаций WRN у пациентов с синдромом Вернера» . Хм. Мутат . 27 (6): 558–67. дои : 10.1002/humu.20337 . ПМК 1868417 . ПМИД 16673358 .
- ^ Лебель М. (2001). «Синдром Вернера: генетическая и молекулярная основа расстройства преждевременного старения». Клетка. Мол. Наука о жизни . 58 (7): 857–67. дои : 10.1007/s00018-001-8398-y . ПМИД 11497235 . S2CID 24801894 .
- ^ Принц П.Р., Эмонд М.Дж., Моннат Р.Дж. (2001). «Потеря функции белка синдрома Вернера способствует аберрантной митотической рекомбинации» . Генс Дев . 15 (8): 933–8. дои : 10.1101/gad.877001 . ПМК 312674 . ПМИД 11316787 .
- ^ Вейрих-Швайгер Х, Вейрих ХГ, Грубер Б, Швайгер М, Хирш-Кауфманн М (1994). «Корреляция между старением и восстановлением ДНК в клетках молодых и старых людей, а также при синдромах преждевременного старения». Мутат. Рез . 316 (1): 37–48. дои : 10.1016/0921-8734(94)90006-х . ПМИД 7507567 .
- ^ Лебель М., Ледер П. (1998). «Делекция в мышиной хеликазе синдрома Вернера вызывает чувствительность к ингибиторам топоизомеразы и потерю клеточной пролиферативной способности» . Учеб. Натл. акад. наук. США . 95 (22): 13097–102. Бибкод : 1998PNAS...9513097L . дои : 10.1073/pnas.95.22.13097 . ПМК 23722 . ПМИД 9789047 .
- ^ Сакамото С., Нисикава К., Хо С.Дж., Гото М., Фуруичи Ю., Симамото А. (2001). «Геликаза Вернера перемещается в ядерные очаги в ответ на агенты, повреждающие ДНК, и локализуется совместно с RPA и Rad51». Генные клетки . 6 (5): 421–30. дои : 10.1046/j.1365-2443.2001.00433.x . ПМИД 11380620 . S2CID 26078155 .
- ^ Джейкобс А.С., Калкинс М.Дж., Джадхав А., Дорджсурен Д., Мэлони Д., Симеонов А., Яруга П., Диздароглу М., Маккалоу А.К., Ллойд Р.С. (2013). «Ингибирование ДНК-гликозилаз с помощью низкомолекулярных аналогов пурина» . ПЛОС ОДИН . 8 (12): е81667. Бибкод : 2013PLoSO...881667J . дои : 10.1371/journal.pone.0081667 . ПМЦ 3857224 . ПМИД 24349107 .
- ^ Немец А.А., Уоллес С.С., Суизи Дж.Б. (октябрь 2010 г.). «Варианты базовых белков эксцизионной репарации: факторы нестабильности генома» . Семинары по биологии рака . 20 (5): 320–8. дои : 10.1016/j.semcancer.2010.10.010 . ПМЦ 3254599 . ПМИД 20955798 .
- ^ Перейти обратно: а б с Пикьерри П., Николай С., Чиньоло Л., Биньями М., Франчитто А. (2012). «Комплекс RAD9-RAD1-HUS1 (9.1.1) взаимодействует с WRN и имеет решающее значение для регулирования его реакции на остановку репликационной вилки» . Онкоген . 31 (23): 2809–23. дои : 10.1038/onc.2011.468 . ПМЦ 3272477 . ПМИД 22002307 .
- ^ Константину А., Тарсунас М., Каров Дж.К., Брош Р.М., Бор В.А., Хиксон И.Д., West SC (2000). «Белок синдрома Вернера (WRN) мигрирует по соединениям Холлидея и локализуется совместно с RPA при аресте репликации» . Представитель ЭМБО . 1 (1): 80–4. дои : 10.1093/embo-reports/kvd004 . ПМЦ 1083680 . ПМИД 11256630 .
- ^ Перейти обратно: а б с Бландер Г., Залле Н., Леал Дж.Ф., Бар-Ор Р.Л., Ю.С.Э., Орен М. (ноябрь 2000 г.). «Белок синдрома Вернера способствует индукции р53 за счет повреждения ДНК» . ФАСЕБ Дж . 14 (14): 2138–40. doi : 10.1096/fj.00-0171fje . ПМИД 11023999 .
- ^ Спилларе Э.А., Ван XW, фон Коббе С., Бор В.А., Хиксон И.Д., Харрис CC (март 2006 г.). «Избыточность ДНК-хеликаз при p53-опосредованном апоптозе» . Онкоген . 25 (14): 2119–23. дои : 10.1038/sj.onc.1209242 . ПМК 1420682 . ПМИД 16288211 .
- ^ Перейти обратно: а б Бернштейн С., Бернштейн Х., Пейн С.М., Гаревал Х. (июнь 2002 г.). «Репарация ДНК / проапоптотические белки двойной роли в пяти основных путях репарации ДНК: надежная защита от канцерогенеза». Мутат Рес . 511 (2): 145–78. дои : 10.1016/s1383-5742(02)00009-1 . ПМИД 12052432 .
- ^ Камат-Лёб А.С., Шен Дж.К., Шмитт М.В., Леб Л.А. (2012). «Экзонуклеаза синдрома Вернера способствует деградации ДНК и высокоточной полимеризации ДНК с помощью ДНК-полимеразы δ человека» . Ж. Биол. Хим . 287 (15): 12480–90. дои : 10.1074/jbc.M111.332577 . ПМК 3320997 . ПМИД 22351772 .
- ^ Гото М., Миллер Р.В., Исикава Ю., Сугано Х. (1996). «Превышение редких видов рака при синдроме Вернера (прогерия у взрослых)». Эпидемиол рака. Биомаркеры Пред . 5 (4): 239–46. ПМИД 8722214 .
- ^ Чун С.Г., Шеффер Д.С., Брайант-Гринвуд ПК (2011). «Геликаза/экзонуклеаза RecQ синдрома Вернера на стыке рака и старения» . Гавайи Мед Дж . 70 (3): 52–5. ПМК 3071901 . ПМИД 21365542 .
- ^ Перейти обратно: а б с Агрело Р., Ченг В.Х., Сетьен Ф., Роперо С., Эспада Х., Фрага М.Ф., Херранц М., Пас М.Ф., Санчес-Сеспедес М., Артига М.Дж., Герреро Д., Кастельс А., фон Коббе С., Бор В.А., Эстеллер М. (2006) . «Эпигенетическая инактивация гена синдрома преждевременного старения Вернера при раке человека» . Учеб. Натл. акад. наук. США . 103 (23): 8822–7. Бибкод : 2006PNAS..103.8822A . дои : 10.1073/pnas.0600645103 . ПМЦ 1466544 . ПМИД 16723399 .
- ^ фон Коббе С., Кармакар П., Давут Л., Опреско П., Зенг X, Брош Р.М., Хиксон И.Д., Бор В.А. (июнь 2002 г.). «Колокализация, физическое и функциональное взаимодействие между белками синдромов Вернера и Блума» . Ж. Биол. Хим . 277 (24): 22035–44. дои : 10.1074/jbc.M200914200 . ПМИД 11919194 .
- ^ Ким С.Т., Лим Д.С., Канман CE, Кастан М.Б. (декабрь 1999 г.). «Специфичность субстратов и идентификация предполагаемых субстратов членов семейства ATM-киназ» . Ж. Биол. Хим . 274 (53): 37538–43. дои : 10.1074/jbc.274.53.37538 . ПМИД 10608806 .
- ^ Кармакар П., Пиотровски Дж., Брош Р.М., Соммерс Дж.А., Миллер С.П., Ченг В.Х., Сноуден К.М., Рамсден Д.А., Бор В.А. (май 2002 г.). «Белок Вернера является мишенью ДНК-зависимой протеинкиназы in vivo и in vitro, а его каталитическая активность регулируется фосфорилированием» . Ж. Биол. Хим . 277 (21): 18291–302. дои : 10.1074/jbc.M111523200 . ПМИД 11889123 .
- ^ Шарма С., Соммерс Дж.А., Ву Л., Бор В.А., Хиксон И.Д., Брош Р.М. (март 2004 г.). «Стимуляция лоскутной эндонуклеазы-1 белком синдрома Блума» . Ж. Биол. Хим . 279 (11): 9847–56. дои : 10.1074/jbc.M309898200 . ПМИД 14688284 .
- ^ Брош Р.М., фон Коббе С., Соммерс Дж.А., Кармакар П., Опреско П.Л., Пиотровски Дж., Дианова И., Дианов Г.Л., Бор В.А. (октябрь 2001 г.). «Белок синдрома Вернера взаимодействует с эндонуклеазой 1 лоскута человека и стимулирует его расщепляющую активность» . ЭМБО Дж . 20 (20): 5791–801. дои : 10.1093/emboj/20.20.5791 . ПМК 125684 . ПМИД 11598021 .
- ^ Перейти обратно: а б Кармакар П., Сноуден К.М., Рамсден Д.А., Бор В.А. (август 2002 г.). «Гетеродимер Ku связывается с обоими концами белка Вернера, и функциональное взаимодействие происходит на N-конце Вернера» . Нуклеиновые кислоты Рез . 30 (16): 3583–91. дои : 10.1093/nar/gkf482 . ПМЦ 134248 . ПМИД 12177300 .
- ^ Перейти обратно: а б Ли Б, Комай Л (сентябрь 2000 г.). «Функциональное взаимодействие между Ku и белком синдрома Вернера при процессинге концов ДНК» . Ж. Биол. Хим . 275 (37): 28349–52. дои : 10.1074/jbc.C000289200 . ПМИД 10880505 .
- ^ Ян Кью, Чжан Р., Ван XW, Спилларе Э.А., Линке С.П., Субраманиан Д., Гриффит Дж.Д., Ли Дж.Л., Хиксон И.Д., Шен Дж.К., Леб Л.А., Мазур С.Дж., Аппелла Е., Брош Р.М., Кармакар П., Бор В.А., Харрис CC (август 2002 г.). «Обработка соединений Холлидея хеликазами BLM и WRN регулируется р53» . Ж. Биол. Хим . 277 (35): 31980–7. дои : 10.1074/jbc.M204111200 . hdl : 10026.1/10341 . ПМИД 12080066 .
- ^ Брош Р.М., Кармакар П., Соммерс Дж.А., Ян К., Ван XW, Спилларе Э.А., Харрис CC, Бор В.А. (сентябрь 2001 г.). «p53 модулирует экзонуклеазную активность белка синдрома Вернера» . Ж. Биол. Хим . 276 (37): 35093–102. дои : 10.1074/jbc.M103332200 . ПМИД 11427532 .
- ^ Родригес-Лопес А.М., Джексон Д.А., Нелин Д.О., Иборра Ф., Уоррен А.В., Кокс Л.С. (февраль 2003 г.). «Характеристика взаимодействия между WRN, дефектом геликазы/экзонуклеазы при синдроме прогероида Вернера и важным фактором репликации PCNA». Мех. Стареющий Дев . 124 (2): 167–74. дои : 10.1016/S0047-6374(02)00131-8 . ПМИД 12633936 . S2CID 37287691 .
- ^ Хуанг С., Берестен С., Ли Б., Осима Дж., Эллис Н.А., Кампизи Дж. (июнь 2000 г.). «Характеристика 3'->5'-экзонуклеазы WRN человека и мыши» . Нуклеиновые кислоты Рез . 28 (12): 2396–405. дои : 10.1093/нар/28.12.2396 . ПМЦ 102739 . ПМИД 10871373 .
- ^ Опреско П.Л., фон Коббе С., Лейн Дж.П., Харриган Дж., Хиксон И.Д., Бор В.А. (октябрь 2002 г.). «Теломерсвязывающий белок TRF2 связывается и стимулирует хеликазы синдрома Вернера и Блума» . Ж. Биол. Хим . 277 (43): 41110–9. дои : 10.1074/jbc.M205396200 . ПМИД 12181313 .
- ^ Бранзей Д., Хаяши Т., Сузуки Х., Масуко Т., Онода Ф., Хо С.Дж., Икеда Х., Симамото А., Фуруичи Ю., Секи М., Эномото Т. (июнь 2001 г.). «Новый белок физически и функционально взаимодействует с продуктом гена синдрома Вернера» . Ж. Биол. Хим . 276 (23): 20364–9. дои : 10.1074/jbc.C100035200 . ПМИД 11301316 .
Дальнейшее чтение
[ редактировать ]- Комай Л, Ли Б (2004). «Белок синдрома Вернера на перекрестке репарации ДНК и апоптоза» . Разработчик старения мехов . 125 (8): 521–8. дои : 10.1016/j.mad.2004.06.004 . ПМИД 15336909 . S2CID 30529954 .
- Ли Дж.В., Харриган Дж., Опреско П.Л., Бор В.А. (2005). «Пути и функции белка синдрома Вернера». Разработчик старения мехов . 126 (1): 79–86. дои : 10.1016/j.mad.2004.09.011 . ПМИД 15610765 . S2CID 39834357 .
- Моннат Р.Ж. младший; Сентиньи Ю (2004). «Белок синдрома Вернера — функция разматывания, объясняющая болезнь» (PDF) . Научные знания о старении. Окружающая среда . 2004 (13): re3. дои : 10.1126/sageke.2004.13.re3 . ПМИД 15056797 . S2CID 15789751 .
- Озгенц А., Леб Л.А. (2005). «Современные достижения в расшифровке функции белка синдрома Вернера». Мутат Рес . 577 (1–2): 237–51. дои : 10.1016/j.mrfmmm.2005.03.020 . ПМИД 15946710 .
- Суонсон С., Сентиньи И., Эмонд М.Дж., Моннат Р.Дж. младший (2004). «Белок синдрома Вернера обладает раздельными функциями рекомбинации и выживания» (PDF) . Репарация ДНК (Амст) . 3 (5): 475–82. дои : 10.1016/j.dnarep.2004.01.002 . ПМИД 15084309 . S2CID 21780379 .
- Мозер М.Дж., Осима Дж., Моннат Р.Дж. (1999). «Мутации WRN при синдроме Вернера» . Хм. Мутат . 13 (4): 271–9. doi : 10.1002/(SICI)1098-1004(1999)13:4<271::AID-HUMU2>3.0.CO;2-Q . ПМИД 10220139 . S2CID 35814236 .
- Кастан М.Б., Лим Д.С. (2001). «Множество субстратов и функций банкомата». Нат. Преподобный мол. Клеточная Биол . 1 (3): 179–86. дои : 10.1038/35043058 . ПМИД 11252893 . S2CID 10691352 .
Внешние ссылки
[ редактировать ]- Осима Дж., Мартин ГМ, Хисама FM (февраль 2012 г.). Синдром Вернера . Вашингтонский университет, Сиэтл. ПМИД 20301687 . НБК1514. В Адам М.П., Эверман Д.Б., Мирза Г.М., Пагон Р.А., Уоллес С.Е., Бин Л.Дж.Х, Грипп К.В., Амемия А. (1993). Пагон Р.А., Берд Т.Д., Долан Ч.Р. и др. (ред.). GeneReviews [Интернет] . Сиэтл, Вашингтон: Вашингтонский университет, Сиэтл. ПМИД 20301295 .
- GeneCard
- База данных мутаций синдрома Вернера, заархивированная 21 июля 2012 г. на Wayback Machine.
галерея PDB |
|---|