ЧЕК1
| ЧЕК1 | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | CHEK1 , CHK1, киназа контрольной точки 1 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 603078 ; МГИ : 1202065 ; Гомологен : 975 ; Генные карты : CHEK1 ; ОМА : ЧЕК1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||




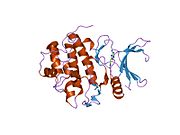
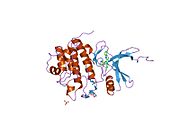
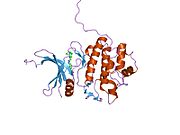
Киназа контрольной точки 1 , обычно называемая Chk1 , представляет собой серин/треонин-специфическую протеинкиназу , которая у человека кодируется CHEK1 геном . [5] [6] Chk1 координирует реакцию на повреждение ДНК (DDR) и реакцию контрольной точки клеточного цикла. [7] Активация Chk1 приводит к инициированию контрольных точек клеточного цикла, остановке клеточного цикла, восстановлению ДНК и гибели клеток, чтобы предотвратить продвижение поврежденных клеток по клеточному циклу.
Открытие
[ редактировать ]В 1993 году Бич и его коллеги первоначально идентифицировали Chk1 как серин/треониновую киназу, которая регулирует фазовый переход G2/M у делящихся дрожжей. [8] Было показано, что конститутивная экспрессия Chk1 в делящихся дрожжах вызывает остановку клеточного цикла. Тот же ген под названием Rad27 был обнаружен у почкующихся дрожжей Карром и его коллегами. В 1997 году гомологи были идентифицированы у более сложных организмов, включая плодовую мушку, человека и мышь. [9] Эти результаты показывают, что Chk1 высоко консервативен от дрожжей до человека. [5]
Структура
[ редактировать ]Человеческий Chk1 расположен на хромосоме 11 на цитогенной полосе 11q22-23. Chk1 имеет N-концевой киназный домен, линкерную область, регуляторный домен SQ/TQ и C-концевой домен. [9] Chk1 содержит четыре остатка Ser/Gln. [8] Активация Chk 1 происходит главным образом за счет фосфорилирования консервативных сайтов Ser-317, Ser-345 и реже Ser-366. [8] [10]
Функция
[ редактировать ]Киназы контрольных точек (Chks) представляют собой протеинкиназы, которые участвуют в контроле клеточного цикла. Были идентифицированы два подтипа киназы контрольной точки: Chk1 и Chk2. Chk1 является центральным компонентом путей надзора за геномом и ключевым регулятором клеточного цикла и выживания клеток. Chk1 необходим для инициации контрольных точек повреждения ДНК , и недавно было показано, что он играет роль в нормальном (ненарушенном) клеточном цикле. [9] Chk1 влияет на различные стадии клеточного цикла, включая фазу S, переход G2/M и фазу M. [8]
Помимо опосредования контрольных точек клеточного цикла, Chk1 также способствует процессам восстановления ДНК, транскрипции генов, производству яйцеклеток, развитию эмбрионов, клеточным ответам на ВИЧ-инфекцию и жизнеспособности соматических клеток. [8] [11]
S-фаза
[ редактировать ]Chk1 необходим для поддержания целостности генома. Chk1 контролирует репликацию ДНК в ненарушенных клеточных циклах и реагирует на генотоксический стресс, если он присутствует. [9] Chk1 распознает нестабильность цепи ДНК во время репликации и может остановить репликацию ДНК, чтобы дать время механизмам репарации ДНК для восстановления генома. [8] Недавно было показано, что Chk1 опосредует механизмы репарации ДНК, активируя различные факторы репарации. Более того, Chk1 связан с тремя конкретными аспектами S-фазы, которые включают регуляцию активации позднего происхождения, контроль процесса элонгации и поддержание стабильности репликационной вилки ДНК. [8]
Переход G2/M
[ редактировать ]В ответ на повреждение ДНК Chk1 является важным преобразователем сигнала для активации контрольной точки G2/M. Активация Chk1 удерживает клетку в фазе G2 до тех пор, пока она не будет готова перейти в митотическую фазу. Эта задержка дает время ДНК для восстановления или гибели клеток, если повреждение ДНК необратимо. [12] Chk1 должен инактивироваться, чтобы клетка могла перейти из фазы G2 в митоз. Уровень экспрессии Chk1 опосредуется регуляторными белками.
Фаза М
[ редактировать ]Chk1 играет регуляторную роль в контрольной точке веретена, однако взаимосвязь менее ясна по сравнению с контрольными точками на других стадиях клеточного цикла. Во время этой фазы активирующий элемент Chk1 оцДНК не может генерироваться, что предполагает альтернативную форму активации. Исследования на клетках куриной лимфомы с дефицитом Chk1 показали повышенный уровень геномной нестабильности и невозможность остановки во время фазы контрольной точки веретена в митозе. [8] Кроме того, гаплонедостаточность эпителиальных клеток молочной железы свидетельствует о смещении хромосом и аномальной сегрегации. Эти исследования показывают, что истощение Chk1 может привести к дефектам контрольной точки веретена, что приводит к митотическим аномалиям.
Взаимодействия
[ редактировать ]Повреждение ДНК вызывает активацию Chk1, что облегчает инициацию реакции на повреждение ДНК (DDR) и контрольных точек клеточного цикла. Реакция на повреждение ДНК представляет собой сеть сигнальных путей, которая приводит к активации контрольных точек, восстановлению ДНК и апоптозу, препятствуя прохождению поврежденных клеток через клеточный цикл.
Активация Chk1
[ редактировать ]Chk1 регулируется ATR посредством фосфорилирования, образуя путь ATR-Chk1. Этот путь распознает одноцепочечную ДНК (оцДНК), которая может быть результатом повреждения, вызванного УФ-излучением, репликационного стресса и межцепочечных перекрестных связей. [8] [9] Часто оцДНК может быть результатом аномальной репликации во время S-фазы из-за разобщения ферментов репликации геликазы и ДНК-полимеразы. [8] Эти структуры оцДНК привлекают ATR и в конечном итоге активируют путь контрольной точки.
Однако активация Chk1 зависит не только от ATR, часто необходимы промежуточные белки, участвующие в репликации ДНК. Регуляторные белки, такие как белок репликации A, Claspin, Tim/Tipin, Rad 17, TopBP1, могут участвовать в активации Chk1. Дополнительные белковые взаимодействия участвуют в индукции максимального фосфорилирования Chk1. Активация Chk1 также может быть ATR-независимой за счет взаимодействия с другими протеинкиназами, такими как PKB/AKT, MAPKAPK и p90/RSK. [8]
Кроме того, было показано, что Chk1 активируется субъединицей Scc1 белка когезина в зиготах. [13]
Остановка клеточного цикла
[ редактировать ]Chk1 взаимодействует со многими нижестоящими эффекторами, вызывая остановку клеточного цикла. В ответ на повреждение ДНК Chk1 в первую очередь фосфорилирует Cdc25, что приводит к его протеасомной деградации. [9] Деградация оказывает ингибирующее действие на образование циклин-зависимых киназных комплексов, которые являются ключевыми движущими силами клеточного цикла. [14] Благодаря нацеливанию на Cdc25 остановка клеточного цикла может происходить в несколько моментов времени, включая переход G1/S, фазу S и переход G2/M. [8] Более того, Chk1 может косвенно нацеливаться на Cdc25 посредством фосфорилирования Nek11.
Киназа WEE1 и PLK1 также подвергаются воздействию Chk1, вызывая остановку клеточного цикла. Фосфорилирование киназы WEE1 ингибирует cdk1, что приводит к остановке клеточного цикла в фазе G2. [8]
Chk1 играет роль в контрольной точке веретена во время митоза, таким образом, взаимодействует с белками сборки веретена Aurora A-киназой и Aurora B-киназой. [9]
восстановление ДНК
[ редактировать ]Недавно было показано, что Chk1 опосредует механизмы репарации ДНК, активируя факторы репарации, такие как ядерный антиген пролиферирующих клеток (PCNA), FANCE, Rad51 и TLK. [8] Chk1 способствует стабилизации репликационной вилки во время репликации и восстановления ДНК, однако необходимы дополнительные исследования для определения основных взаимодействий. [9]
Клиническая значимость
[ редактировать ]Chk1 играет центральную роль в координации реакции на повреждение ДНК и поэтому представляет собой область большого интереса в онкологии и разработке методов лечения рака. [15] Первоначально считалось, что Chk1 действует как супрессор опухоли из-за регуляторной роли, которую он выполняет среди клеток с повреждением ДНК. Однако не было обнаружено доказательств гомозиготной потери функции мутантов Chk1 в опухолях человека. [8] Вместо этого было показано, что Chk1 сверхэкспрессируется во многих опухолях, включая рак молочной железы, толстой кишки, печени, желудка и носоглотки. [8] Существует положительная корреляция с экспрессией Chk1, степенью опухоли и рецидивом заболевания, что позволяет предположить, что Chk1 может способствовать росту опухоли. [8] [9] [15] Chk1 необходим для выживания клеток, и благодаря высокому уровню экспрессии в опухолях эта функция может вызывать пролиферацию опухолевых клеток. Кроме того, исследование показало, что нацеливание на Chk1 реактивирует опухолесупрессирующую активность комплекса протеинфосфатазы 2А (PP2A) в раковых клетках. [16] Исследования показали, что полная потеря Chk1 подавляет химически индуцирующий канцерогенез, однако гаплонедостаточность Chk1 приводит к прогрессированию опухоли. [9] Из-за возможности участия Chk1 в продвижении опухоли киназа и родственные ей сигнальные молекулы могут быть потенциально эффективными терапевтическими мишенями. В терапии рака используются методы, повреждающие ДНК, такие как химиотерапия и ионизирующее излучение, чтобы ингибировать пролиферацию опухолевых клеток и вызывать остановку клеточного цикла. [17] Опухолевые клетки с повышенным уровнем Chk1 приобретают преимущества выживания благодаря способности переносить более высокий уровень повреждения ДНК. Следовательно, Chk1 может способствовать устойчивости к химиотерапии. [18] Чтобы оптимизировать химиотерапию, Chk1 необходимо ингибировать, чтобы уменьшить преимущество выживаемости. [7] Ген Chk1 можно эффективно заглушить путем нокдауна siRNA для дальнейшего анализа на основе независимой проверки. [19] Ингибируя Chk1, раковые клетки теряют способность восстанавливать поврежденную ДНК, что позволяет химиотерапевтическим агентам работать более эффективно. Сочетание методов лечения, повреждающих ДНК, таких как химиотерапия или лучевая терапия, с ингибированием Chk1 усиливает целенаправленную гибель клеток и обеспечивает синтетическую летальность. [20] Многие виды рака в значительной степени зависят от остановки клеточного цикла, опосредованной Chk1, особенно если рак имеет дефицит р53. [21] Примерно 50% случаев рака обладают мутациями р53, что иллюстрирует зависимость, которую многие виды рака могут иметь от пути Chk1. [22] [23] [24] Ингибирование Chk1 позволяет избирательно воздействовать на мутантные клетки p53, поскольку уровни Chk1 с большей вероятностью будут высоко экспрессироваться в опухолевых клетках с дефицитом p53. [15] [25] Несмотря на то, что этот метод ингибирования является узконаправленным, недавние исследования показали, что Chk1 также играет роль в нормальном клеточном цикле. [26] Следовательно, при разработке новых методов лечения необходимо учитывать нецелевые эффекты и токсичность, связанные с комбинированной терапией с использованием ингибиторов Chk1. [27]
В рамках комбинированного вычислительного подхода для анализа был выбран набор полусинтетических аминоарилбензосубереновых молекул растительного происхождения, из которых Bch10 рассматривается как потенциальный ингибитор CHK1 по сравнению с пятью лучшими сокристаллизованными ингибиторами на основе их сродства к связыванию и профиля токсичности. . [28]
Мейоз
[ редактировать ]Во время мейоза у человека и мыши протеинкиназа CHEK1 важна для интеграции восстановления повреждений ДНК с остановкой клеточного цикла. [29] CHEK1 экспрессируется в семенниках и связывается с мейотическими синаптонемными комплексами на стадиях зигонемы и пахинемы . [29] CHEK1, вероятно, действует как интегратор сигналов ATM и ATR и может участвовать в мониторинге мейотической рекомбинации . [29] В ооцитах мышей CHEK1, по-видимому, незаменим для остановки профазы I и функционирования в контрольной точке G2/M . [30]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000149554 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000032113 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б Санчес И., Вонг С., Тома Р.С., Ричман Р., Ву З., Пивника-Вормс Х. и др. (сентябрь 1997 г.). «Сохранение пути контрольной точки Chk1 у млекопитающих: связь повреждения ДНК с регуляцией Cdk через Cdc25». Наука . 277 (5331): 1497–501. дои : 10.1126/science.277.5331.1497 . ПМИД 9278511 .
- ^ Флэггс Дж., Плаг А.В., Данкс К.М., Мундт К.Э., Форд Дж.К., Куиггл М.Р. и др. (декабрь 1997 г.). «АТМ-зависимые взаимодействия гомолога chk1 млекопитающих с мейотическими хромосомами» . Современная биология . 7 (12): 977–86. Бибкод : 1997CBio....7..977F . дои : 10.1016/S0960-9822(06)00417-9 . ПМИД 9382850 . S2CID 14734991 .
- ^ Jump up to: а б Макнили С., Бекманн Р., Бенс Лин А.К. (апрель 2014 г.). «Снова CHEK: новый взгляд на разработку ингибиторов Chk1 для терапии рака». Фармакология и терапия . 142 (1): 1–10. doi : 10.1016/j.pharmthera.2013.10.005 . ПМИД 24140082 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Чжан Ю, Хантер Т. (март 2014 г.). «Роль Chk1 в клеточной биологии и терапии рака» . Международный журнал рака . 134 (5): 1013–23. дои : 10.1002/ijc.28226 . ПМК 3852170 . ПМИД 23613359 .
- ^ Jump up to: а б с д и ж г час я дж Патил М., Пабла Н., Донг З. (ноябрь 2013 г.). «Киназа контрольной точки 1 в реакции на повреждение ДНК и регуляции клеточного цикла» . Клеточные и молекулярные науки о жизни . 70 (21): 4009–21. дои : 10.1007/s00018-013-1307-3 . ПМЦ 3731415 . ПМИД 23508805 .
- ^ Капарелли М.Л., О'Коннелл М.Дж. (март 2013 г.). «Регуляторные мотивы в Чк1» . Клеточный цикл . 12 (6): 916–22. дои : 10.4161/cc.23881 . ПМЦ 3637350 . ПМИД 23422000 .
- ^ Рут К.С., Дэй Ф.Р., Хуссейн Дж., Мартинес-Маршал А., Айкен С.Э., Азад А. и др. (август 2021 г.). «Генетическое понимание биологических механизмов, управляющих старением яичников человека» . Природа . 596 (7872): 393–397. Бибкод : 2021Natur.596..393R . дои : 10.1038/s41586-021-03779-7 . ПМЦ 7611832 . ПМИД 34349265 .
- ^ Мейт М. (сентябрь 2010 г.). «Chk1 подавлял гибель клеток» . Отделение клеток . 5:21 . дои : 10.1186/1747-1028-5-21 . ПМЦ 2939633 . ПМИД 20813042 .
- ^ Ладштеттер С., Тачибана-Конвальски К. (декабрь 2016 г.). «Механизм наблюдения обеспечивает восстановление повреждений ДНК во время зиготического перепрограммирования» . Клетка . 167 (7): 1774–1787.e13. дои : 10.1016/j.cell.2016.11.009 . ПМК 5161750 . ПМИД 27916276 .
- ^ Лю Q, Гунтуку С., Цуй XS, Мацуока С., Кортес Д., Тамай К. и др. (июнь 2000 г.). «Chk1 представляет собой незаменимую киназу, которая регулируется Atr и необходима для контрольной точки повреждения ДНК G (2) / M» . Гены и развитие . 14 (12): 1448–59. дои : 10.1101/gad.840500 . ПМК 316686 . ПМИД 10859164 .
- ^ Jump up to: а б с Гото Х, Идзава И, Ли П, Инагаки М (июль 2012 г.). «Новая регуляция киназы 1 контрольной точки: является ли киназа 1 контрольной точки хорошим кандидатом для противораковой терапии?» . Раковая наука . 103 (7): 1195–200. дои : 10.1111/j.1349-7006.2012.02280.x . ПМЦ 7659239 . ПМИД 22435685 . S2CID 205237831 .
- ^ Ханна А., Кауко О., Бёкельман С., Лайне А., Шрек И., Партанен Дж.И. и др. (ноябрь 2013 г.). «Нацеливание на Chk1 реактивирует активность опухолевого супрессора PP2A в раковых клетках» . Исследования рака . 73 (22): 6757–69. дои : 10.1158/0008-5472.CAN-13-1002 . ПМК 3870284 . ПМИД 24072747 .
- ^ Смит Дж., То Л.М., Сюй Н., Гиллеспи Д.А. (2010). Пути ATM-Chk2 и ATR-Chk1 в передаче сигналов о повреждении ДНК и раке . Том. 108. С. 73–112. дои : 10.1016/B978-0-12-380888-2.00003-0 . ISBN 978-0-12-380888-2 . ПМИД 21034966 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Лян Ю, Линь С.Ю., Бруникади, Госс Дж., Ли К. (апрель 2009 г.). «Пути реакции на повреждение ДНК при подавлении опухоли и лечении рака». Всемирный журнал хирургии . 33 (4): 661–6. дои : 10.1007/s00268-008-9840-1 . ПМИД 19034564 . S2CID 13599990 .
- ^ Мункачи Г., Штупински З., Херман П., Бан Б., Пенцвалло З., Сарвас Н. и др. (сентябрь 2016 г.). «Подтверждение эффективности подавления РНКи с использованием данных генного массива показывает 18,5% частоты неудач в 429 независимых экспериментах» . Молекулярная терапия: нуклеиновые кислоты . 5 (9): е366. дои : 10.1038/mtna.2016.66 . ПМК 5056990 . ПМИД 27673562 .
- ^ Толедо Л.И., Мурга М., Фернандес-Капетильо О. (август 2011 г.). «Нацеливание на киназы ATR и Chk1 для лечения рака: новая модель новых (и старых) лекарств» . Молекулярная онкология . 5 (4): 368–73. дои : 10.1016/j.molonc.2011.07.002 . ПМК 3590794 . ПМИД 21820372 .
- ^ Чен З, Сяо З, Гу ВЗ, Сюэ Дж, Буй МХ, Ковар П и др. (декабрь 2006 г.). «Селективные ингибиторы Chk1 по-разному повышают чувствительность раковых клеток с дефицитом р53 к противораковым препаратам» . Международный журнал рака . 119 (12): 2784–94. дои : 10.1002/ijc.22198 . ПМИД 17019715 . S2CID 22922827 .
- ^ Могери-Сакка М., Бартуччи М., Де Мария Р. (август 2013 г.). «Ингибиторы контрольной точки киназы 1 для усиления системной противораковой терапии». Обзоры лечения рака . 39 (5): 525–33. дои : 10.1016/j.ctrv.2012.10.007 . ПМИД 23207059 .
- ^ Тао З.Ф., Лин Н.Х. (июль 2006 г.). «Ингибиторы Chk1 для нового лечения рака». Противораковые агенты в медицинской химии . 6 (4): 377–88. дои : 10.2174/187152006777698132 . ПМИД 16842237 .
- ^ Ма CX, Джанетка Дж.В., Пивница-Вормс Х (февраль 2011 г.). «Смерть от разрывов: ингибиторы CHK1 как средство лечения рака» . Тенденции молекулярной медицины . 17 (2): 88–96. doi : 10.1016/j.molmed.2010.10.009 . ПМК 6905465 . ПМИД 21087899 .
- ^ Зенвирт С., Кравченко-Балаша Н., Левицкий А. (ноябрь 2010 г.). «Статус р53 в раковых клетках человека не предсказывает эффективность ингибиторов киназы Chk1 в сочетании с химиотерапевтическими агентами» . Онкоген . 29 (46): 6149–59. дои : 10.1038/onc.2010.343 . ПМИД 20729914 .
- ^ Томпсон Р., Истман А. (сентябрь 2013 г.). «Терапевтический потенциал ингибиторов Chk1 против рака: как механистические исследования влияют на дизайн клинических исследований» . Британский журнал клинической фармакологии . 76 (3): 358–69. дои : 10.1111/bcp.12139 . ПМЦ 3769664 . ПМИД 23593991 .
- ^ Дент П., Тан Ю., Якуб А., Дай Ю., Фишер П.Б., Грант С. (апрель 2011 г.). «Ингибиторы CHK1 в комбинированной химиотерапии: мышление за пределами клеточного цикла» . Молекулярные вмешательства . 11 (2): 133–40. дои : 10.1124/ми.11.2.11 . ПМК 3109860 . ПМИД 21540473 .
- ^ Сингх Р., Бхардвадж В.К., Шарма Дж., Дас П., Пурохит Р. (январь 2021 г.). «Открытие и оценка in silico молекул аминоарилбензосуберена как новых детерминант ингибитора контрольной точки киназы 1» . Геномика . 113 (1): 707–715. дои : 10.1016/j.ygeno.2020.10.001 . ПМИД 33065246 . S2CID 223556335 .
- ^ Jump up to: а б с Флэггс Дж., Плаг А.В., Данкс К.М., Мундт К.Э., Форд Дж.К., Куиггл М.Р. и др. (декабрь 1997 г.). «АТМ-зависимые взаимодействия гомолога chk1 млекопитающих с мейотическими хромосомами» . Современная биология . 7 (12): 977–86. Бибкод : 1997CBio....7..977F . дои : 10.1016/s0960-9822(06)00417-9 . ПМИД 9382850 . S2CID 14734991 .
- ^ Чен Л., Чао С.Б., Ван З.Б., Ци С.Т., Чжу С.Л., Ян С.В. и др. (май 2012 г.). «Киназа контрольной точки 1 необходима для регуляции мейотического клеточного цикла в ооцитах мыши» . Клеточный цикл . 11 (10): 1948–55. дои : 10.4161/cc.20279 . ПМИД 22544319 .
Дальнейшее чтение
[ редактировать ]- Джачча А.Дж., Кастан М.Б. (октябрь 1998 г.). «Сложность модуляции p53: возникающие закономерности из расходящихся сигналов» . Гены и развитие . 12 (19): 2973–83. дои : 10.1101/gad.19.12.2973 . ПМИД 9765199 .
- Кастан М.Б., Лим Д.С. (декабрь 2000 г.). «Множество субстратов и функций банкомата». Обзоры природы. Молекулярно-клеточная биология . 1 (3): 179–86. дои : 10.1038/35043058 . ПМИД 11252893 . S2CID 10691352 .
- Чини CC, Чен Дж (2005). «Класпин, регулятор Chk1 в пути стресса репликации ДНК». Восстановление ДНК . 3 (8–9): 1033–7. дои : 10.1016/j.dnarep.2004.03.001 . ПМИД 15279790 .
- Пэн С.И., Грейвс П.Р., Тома Р.С., Ву З., Шоу А.С., Пивиника-Вормс Х. (сентябрь 1997 г.). «Контроль контрольных точек митоза и G2: регуляция связывания белка 14-3-3 путем фосфорилирования Cdc25C на серине-216». Наука . 277 (5331): 1501–5. дои : 10.1126/science.277.5331.1501 . ПМИД 9278512 .
- Оуян Б., Ли В., Пан Х., Медоуз Дж., Хоффманн И., Дай В. (октябрь 1999 г.). «Физическая ассоциация и фосфорилирование протеинфосфатазы Cdc25C с помощью Prk» . Онкоген . 18 (44): 6029–36. дои : 10.1038/sj.onc.1202983 . ПМИД 10557092 .
- Ким С.Т., Лим Д.С., Канман С.Э., Кастан М.Б. (декабрь 1999 г.). «Специфичность субстратов и идентификация предполагаемых субстратов членов семейства ATM-киназ» . Журнал биологической химии . 274 (53): 37538–43. дои : 10.1074/jbc.274.53.37538 . ПМИД 10608806 .
- Ши С.Ю., Ан Дж., Тамай К., Тая Ю., Привес К. (февраль 2000 г.). «Человеческие гомологи киназ контрольных точек Chk1 и Cds1 (Chk2) фосфорилируют p53 в нескольких сайтах, индуцирующих повреждение ДНК» . Гены и развитие . 14 (3): 289–300. дои : 10.1101/gad.14.3.289 . ПМК 316358 . ПМИД 10673501 .
- Грейвс П.Р., Ю.Л., Шварц Дж.К., Гейлс Дж., Сосвилл Э.А., О'Коннор П.М. и др. (февраль 2000 г.). «Протеинкиназа Chk1 и регуляторные пути Cdc25C являются мишенями противоракового агента UCN-01» . Журнал биологической химии . 275 (8): 5600–5. дои : 10.1074/jbc.275.8.5600 . ПМИД 10681541 .
- Семба С., Оуян Х., Хан С.Ю., Като Ю., Хории А. (апрель 2000 г.). «Анализ генов-кандидатов-мишеней для мутаций при микросателлитном нестабильном раке колоректума, желудка и эндометрия». Международный журнал онкологии . 16 (4): 731–7. дои : 10.3892/ijo.16.4.731 . ПМИД 10717241 .
- Чен П., Луо С., Дэн Ю., Райан К., Регистр Дж., Маргосиак С. и др. (март 2000 г.). «Кристаллическая структура 1,7 A киназы контрольной точки клеточного цикла человека Chk1: значение для регуляции Chk1» . Клетка . 100 (6): 681–92. дои : 10.1016/S0092-8674(00)80704-7 . ПМИД 10761933 . S2CID 15626948 .
- Лю Q, Гунтуку С., Цуй XS, Мацуока С., Кортес Д., Тамай К. и др. (июнь 2000 г.). «Chk1 представляет собой незаменимую киназу, которая регулируется Atr и необходима для контрольной точки повреждения ДНК G (2) / M» . Гены и развитие . 14 (12): 1448–59. дои : 10.1101/gad.840500 . ПМК 316686 . ПМИД 10859164 .
- Булавин Д.В., Хигасимото Ю., Попов И.Дж., Гаарде В.А., Басрур В., Потапова О. и др. (май 2001 г.). «Для запуска контрольной точки G2 / M после ультрафиолетового излучения требуется киназа p38». Природа . 411 (6833): 102–7. дои : 10.1038/35075107 . ПМИД 11333986 . S2CID 4410763 .
- Чжао Х., Пивница-Вормс Х. (июль 2001 г.). «Пути контрольных точек, опосредованные ATR, регулируют фосфорилирование и активацию Chk1 человека» . Молекулярная и клеточная биология . 21 (13): 4129–39. дои : 10.1128/MCB.21.13.4129-4139.2001 . ПМК 87074 . ПМИД 11390642 .
- Фейджу С., Холл-Джексон С., Ву Р., Дженкинс Д., Лейтч Дж., Гилберт Д.М. и др. (сентябрь 2001 г.). «Активация Chk1 млекопитающих во время остановки репликации ДНК: роль Chk1 в контрольной точке внутри S-фазы, контролирующей запуск начала репликации» . Журнал клеточной биологии . 154 (5): 913–23. дои : 10.1083/jcb.200104099 . ПМК 1255922 . ПМИД 11535615 .
- Се С., Ву Х., Ван К., Когсвелл Дж. П., Хусейн И., Конн С. и др. (ноябрь 2001 г.). «Plk3 функционально связывает повреждение ДНК с остановкой клеточного цикла и апоптозом, по крайней мере частично, через путь p53» . Журнал биологической химии . 276 (46): 43305–12. дои : 10.1074/jbc.M106050200 . ПМИД 11551930 .
- Латонен Л., Тая Ю., Лайхо М. (октябрь 2001 г.). «УФ-излучение индуцирует дозозависимую регуляцию ответа р53 и модулирует взаимодействие p53-HDM2 в фибробластах человека» . Онкоген . 20 (46): 6784–93. дои : 10.1038/sj.onc.1204883 . ПМИД 11709713 .
Внешние ссылки
[ редактировать ]- Местоположение генома человека CHEK1 и CHEK1 страница сведений о гене в браузере генома UCSC .
галерея PDB |
|---|