PIM2 (ген)
| ПИМ2 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | PIM2 , протоонкоген Pim-2, серин/треонин киназа | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 300295 ; МГИ : 97587 ; Гомологен : 17098 ; Генные карты : PIM2 ; ОМА : PIM2 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Серин/треонин-протеинкиназа Pim-2 представляет собой фермент , который у человека кодируется геном PIM2 (провирусная интеграция вируса Молони 2). [ 5 ] [ 6 ] Фермент представляет собой серин/треониновую киназу , которая играет роль в росте клеток , пролиферации, апоптозе и регуляции каскадов сигнальной трансдукции . [ 7 ]
Структура
[ редактировать ]До сих пор большая часть структурной информации, относящейся к семейству киназ PIM, ограничивалась PIM1 . В результате большая часть усилий по разработке ингибиторов также была направлена на PIM1. Последовательность PIM2 на 55% идентична последовательности PIM1, а структура PIM2 довольно тесно связана с PIM1. [ 8 ] Как и PIM1, PIM2 демонстрирует билобальную киназную архитектуру с конститутивно активной закрытой конформацией. Основная цепь обеих молекул идентична, за исключением двух гибких участков в N-концевой доле.

Наиболее значимым структурным различием между PIM1 и PIM2 является отсутствие терминальной αJ-спирали в PIM2. Последние 23 остатка PIM2 сильно отличаются от PIM1, поскольку PIM2 содержит 6 остатков пролина в этой области и, как полагают, не образует одинаковые третичные структуры. В результате отсутствие взаимодействий, присутствующих в этой области, может увеличивать гибкость PIM2 внутри N-концевой доли киназы и способствовать неупорядоченным участкам структуры PIM2. [ 9 ]
Функция
[ редактировать ]PIM2 экспрессируется на высоких уровнях в мозге и лимфоидных клетках . соединения PIM1-3 Мыши с нокаутом , пережившие перинатальный период, показали значительное уменьшение размеров тела. Это говорит о том, что ферменты PIM важны для роста тела . [ 10 ] Эксперименты показали, что PIM1 и PIM2 необходимы для цитокин-зависимой пролиферации и выживания лимфоцитов. [ 11 ] Эксперименты с трансгенными мышами с индуцированными лимфомами выявили повышенные уровни PIM2 как частое, но позднее событие онкогенеза . [ 12 ]
Эксперименты, проведенные по ядерной транслокации ядерного фактора κB ( NFκB (PNI) человека ) при периневральной инвазии , показали, что активация NFκB и его нижестоящей мишени, PIM2, являются компонентами сигнального каскада антиапоптоза, который связан с раковыми клетками в PNI. Этот каскад может регулировать ингибирование апоптоза. Исследование также показало, что повышенные уровни PIM2 связаны с ПНИ. [ 13 ] Таким образом, киназа PIM2 стала ключевой мишенью для восстановления апоптоза при лекарственно-устойчивом раке человека. [ 14 ] [ 15 ] [ 16 ]
Механизм
[ редактировать ]В известных кристаллических структурах PIM1 и PIM2 принимают активную конформацию. Обычно активное состояние киназ характеризуется наличием консервативного лизина , закрытой лепестковой конформации и хорошо структурированного активационного сегмента. Сегмент активации часто требует фосфорилирования для проявления каталитической активности . После фосфорилирования активный сегмент складывается на нижнюю долю и реорганизует сайт связывания пептида , что, следовательно, приводит к ферментативной активации. Однако киназы PIM каталитически активны без фосфорилирования. Кристаллические структуры показывают, что нефосфорилированный активационный сегмент образует множество полярных взаимодействий с нижней долей киназы, что стабилизирует активную конформацию. Хотя киназа PIM осуществляет аутофосфорилирование , функциональные последствия неизвестны. [ 17 ]
Ингибиторы
[ редактировать ]-
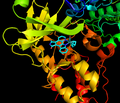 Ингибитор действует как миметик АТФ в кармане связывания АТФ.
Ингибитор действует как миметик АТФ в кармане связывания АТФ. -
 Стауроспорин и соединение рутения 12
Стауроспорин и соединение рутения 12 -
 Общая структура рутениевого ингибитора
Общая структура рутениевого ингибитора



PIM2 (наряду с PIM1) имеет уникальный карман для связывания АТФ с шарнирной областью, что делает его привлекательной мишенью для мощных низкомолекулярных ингибиторов киназы PIM . [ 18 ]
Многие ингибиторы часто более селективны в отношении PIM1 и PIM3, чем PIM2. Другими словами, PIM2 обычно ингибируется с гораздо меньшей эффективностью. Структурные модели пока не могут объяснить это явление. Однако это может быть связано с различиями в динамических свойствах разных изоформ PIM .
В ряду рутенийорганических соединений [ 19 ] на основе стауроспоринового каркаса [ 20 ] соединение 12 давало практически полное ингибирование при концентрации 10 нМ. Однако он был немного менее эффективен против PIM1.
SAR предполагает, что добавление потенциальных групп водородных связей в положениях R1 и R2 резко увеличивает эффективность против обеих киназ. Подобная замена положения R3 была менее эффективной, а замена галогеном была еще более разрушительной.
Клиническое значение
[ редактировать ]В исследовании с участием 48 пациентов с неходжкинской лимфомой (НХЛ) и лимфоцитарным лейкозом экспрессию hPim-2 анализировали с использованием гибридизации in-situ , количественной RT-PCR и анализа FACS. Исследования показали более высокие уровни экспрессии в НХЛ по сравнению с нормальными лимфоцитами, а также при хроническом лимфоцитарном лейкозе по сравнению с нормальными В-клетками . [ 21 ]
Повышенные уровни PIM2 также были обнаружены в первичных бластах пациентов с острым миелолейкозом . PIM2 может быть важной киназой в фосфорилировании 4E-BP1. Конститутивное фосфорилирование 4E-BP1 обычно обнаруживается при раке и способствует устойчивой трансляции транскриптов, связанных со злокачественными новообразованиями, среди которых c-Myc и Cyclin D. Нокдаун PIM2 iRNA сильно снижал накопление онкогенных белков. [ 22 ] В результате PIM2 может стать привлекательной мишенью для острого миелолейкоза.
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000102096 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000031155 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Байтел Д., Шалом С., Мадгар И., Вайсенберг Р., Дон Дж. (декабрь 1998 г.). «Человеческий протоонкоген Pim-2 и его тестикулярная экспрессия». Биохим Биофиз Акта . 1442 (2–3): 274–85. дои : 10.1016/s0167-4781(98)00185-7 . ПМИД 9804974 .
- ^ «Ген Энтрез: онкоген PIM2 pim-2» .
- ^ Альварадо Ю., Джайлз Ф.Дж., Мечи RT (2012). «Киназы PIM при гематологическом раке». Эксперт Рев Гематол . 5 (1): 81–96. дои : 10.1586/эхм.11.69 . ПМИД 22272708 . S2CID 29518294 .
- ^ Дебрецени Дж. Э., Буллок А. Н., Атилла Г. Е., Уильямс Д. С., Брегман Х., Кнапп С., Меггерс Э. (февраль 2006 г.). «Полусэндвич-комплексы рутения, связанные с протеинкиназой Pim-1». Ангеванде Хеми . 45 (10): 1580–5. дои : 10.1002/anie.200503468 . ПМИД 16381041 .
- ^ Буллок А.Н., Руссо С., Амос А., Пагано Н., Брегман Х., Дебрецени Дж.Е., Ли В.Х., фон Делфт Ф., Меггерс Э., Кнапп С. (2005). «Кристаллическая структура киназы PIM2 в комплексе с рутенийовым ингибитором» . ПЛОС ОДИН . 4 (10): е7112. дои : 10.1371/journal.pone.0007112 . ПМЦ 2743286 . ПМИД 19841674 .
- ^ Миккерс Х., Навейн М., Аллен Дж., Брауэрс С., Верховен Э., Джонкерс Дж., Бернс А. (2004). «Мыши с дефицитом всех киназ PIM демонстрируют уменьшенный размер тела и нарушенную реакцию на гемопоэтические факторы роста» . Мол Клеточная Биол . 24 (13): 6104–6115. дои : 10.1128/MCB.24.13.6104-6115.2004 . ПМК 480904 . ПМИД 15199164 .
- ^ Тахванайнен Дж., Кюляниеми М.К., Кандури К., Гупта Б., Лахтеенмяки Х., Каллонен Т., Раджавуори А., Расул О., Коскинен П.Дж., Рао К.В., Ляхдесмяки Х., Лахесмаа Р. (2013). «Сайт провирусной интеграции киназ вируса мышиного лейкоза Молони (PIM) способствует дифференцировке клеток Т-хелперов 1 человека» . Журнал биологической химии . 288 (5): 3048–3058. дои : 10.1074/jbc.M112.361709 . ПМЦ 3561529 . ПМИД 23209281 .
- ^ Брейер М.Л., Кайперс Х.Т., Бернс А. (март 1989 г.). «Доказательства участия pim-2, нового распространенного места вставки провирусов, в прогрессирование лимфом» . Журнал ЭМБО . 8 (3): 743–8. дои : 10.1002/j.1460-2075.1989.tb03434.x . ПМК 400870 . ПМИД 2721500 .
- ^ Аяла Г.Е., Дай Х., Иттманн М., Ли Р., Пауэлл М., Фролов А. и др. (сентябрь 2004 г.). «Механизмы роста и выживания, связанные с периневральной инвазией при раке простаты» . Исследования рака . 64 (17): 6082–90. дои : 10.1158/0008-5472.CAN-04-0838 . ПМИД 15342391 .
- ^ Джайлз Ф (2005). «Ингибитор киназы Пим, пожалуйста» . Кровь . 105 (11): 4158–4159. дои : 10.1182/кровь-2005-03-1150 .
- ^ Мэлоун Т., Шефер Л., Саймон Н., Хиви С., Кафф С., Финн С. и др. (март 2020 г.). «Текущие перспективы использования киназ PIM для преодоления механизмов лекарственной устойчивости и уклонения от иммунитета при раке» (PDF) . Фармакология и терапия . 207 : 107454. doi : 10.1016/j.pharmthera.2019.107454 . ПМИД 31836451 . S2CID 209357486 .
- ^ Лущак С., Кумар С., Сатьядеван В.К., Симпсон Б.С., Гейтли К.А., Уитакер ХК, Хиви С. (2020). «Ингибирование киназы PIM: совместные терапевтические подходы при раке простаты» . Сигнальная трансдукция и таргетная терапия . 5 :7. дои : 10.1038/s41392-020-0109-y . ПМК 6992635 . ПМИД 32025342 .
- ^ Браулт Л., Гассер С., Брахер Ф., Хубер К., Кнапп С., Шваллер Дж. (2010). «Серин/треониновые киназы PIM в патогенезе и терапии гематологических злокачественных опухолей и солидного рака» . Гематологическая . 95 (6): 1004–1015. дои : 10.3324/haematol.2009.017079 . ПМЦ 2878801 . ПМИД 20145274 .
- ^ Погачич В., Буллок А.Н., Федоров О., Филиппакопулос П., Гассер С., Бионди А., Мейер-Монард С., Кнапп С., Шваллер Дж. (июль 2007 г.). «Структурный анализ идентифицирует имидазо[1,2-b]пиридазины как ингибиторы киназы PIM с противолейкемической активностью in vitro» . Исследования рака . 67 (14): 6916–24. дои : 10.1158/0008-5472.CAN-07-0320 . ПМИД 17638903 .
- ^ Пагано Н., Максимоска Дж., Брегман Х., Уильямс Д.С., Вебстер Р.Д., Сюэ Ф., Меггерс Э. (апрель 2007 г.). «Полусэндвичевые комплексы рутения как ингибиторы протеинкиназы: дериватизация фармакофорного лиганда пиридокарбазола». Органическая и биомолекулярная химия . 5 (8): 1218–27. дои : 10.1039/b700433h . ПМИД 17406720 .
- ^ Меггерс Э. (июнь 2007 г.). «Исследование биологически значимого химического пространства с помощью металлокомплексов». Современное мнение в области химической биологии . 11 (3): 287–92. дои : 10.1016/j.cbpa.2007.05.013 . ПМИД 17548234 .
- ^ Коэн А.М., Гринблат Б., Бесслер Х., Кристт Д., Кремер А., Шварц А., Гальперин М., Шалом С., Меркель Д., Дон Дж. (2004). «Повышенная экспрессия гена hPim-2 при хроническом лимфоцитарном лейкозе человека и неходжкинской лимфоме». Лейкемия и лимфома . 45 (5): 951–955. дои : 10.1080/10428190310001641251 . ПМИД 15291354 . S2CID 24369286 .
- ^ Моерке Н.Дж., Актас Х., Чен Х., Кантель С., Рейбарх М.Ю., Фахми А., Гросс Дж.Д., Дегтерев А., Юань Дж., Чорев М., Гальперин Дж.А., Вагнер Г. (январь 2007 г.). «Низкомолекулярное ингибирование взаимодействия факторов инициации трансляции eIF4E и eIF4G» . Клетка . 128 (2): 257–67. дои : 10.1016/j.cell.2006.11.046 . ПМИД 17254965 .
Дальнейшее чтение
[ редактировать ]- Ян Б, Земскова М, Холдер С, Чин В, Крафт А, Коскинен П.Дж., Лилли М (2003). «Киназа PIM-2 фосфорилирует BAD по серину 112 и обращает вспять вызванную BAD гибель клеток» . Ж. Биол. Хим . 278 (46): 45358–67. дои : 10.1074/jbc.M307933200 . ПМИД 12954615 .
- Коэн А.М., Гринблат Б., Бесслер Х., Кристт Д., Кремер А., Шварц А., Гальперин М., Шалом С., Меркель Д., Дон Дж. (2004). «Повышенная экспрессия гена hPim-2 при хроническом лимфоцитарном лейкозе человека и неходжкинской лимфоме». Лейк. Лимфома . 45 (5): 951–5. дои : 10.1080/10428190310001641251 . ПМИД 15291354 . S2CID 24369286 .
- Хаммерман П.С., Фокс С.Дж., Чиналли Р.М., Сюй А., Вагнер Дж.Д., Линдстен Т., Томпсон CB (2005). «Трансформация лимфоцитов с помощью Pim-2 зависит от активации ядерного фактора каппаВ» . Рак Рез . 64 (22): 8341–8. дои : 10.1158/0008-5472.CAN-04-2284 . ПМИД 15548703 .
- Райнио Э.М., Альфорс Х., Картер К.Л., Рууска М., Матикайнен С., Кифф Э., Коскинен П.Дж. (2005). «Киназы Pim активируются во время заражения вирусом Эпштейна-Барра и повышают активность EBNA2». Вирусология . 333 (2): 201–6. дои : 10.1016/j.virol.2005.01.001 . ПМИД 15721354 .
- Дай Х., Ли Р., Уилер Т., Диас де Вивар А., Фролов А., Тахир С., Агульник И., Томпсон Т., Роули Д., Аяла Г. (2005). «Повышение уровня регуляции Pim-2: биологические последствия, связанные с прогрессированием заболевания и перинуеральной инвазией при раке простаты». Простата . 65 (3): 276–86. дои : 10.1002/pros.20294 . ПМИД 16015593 . S2CID 44856689 .
- Руал Дж.Ф., Венкатесан К., Хао Т., Хирозане-Кисикава Т., Дрико А., Ли Н., Берриз Г.Ф., Гиббонс Ф.Д., Дрез М., Айви-Гедехуссу Н., Клитгорд Н., Саймон С., Боксем М., Мильштейн С., Розенберг Дж., Голдберг Д.С., Чжан Л.В., Вонг С.Л., Франклин Г., Ли С., Альбала Дж.С., Лим Дж., Фротон С., Лламосас Е., Чевик С., Бекс С., Ламеш П., Сикорски Р.С., Ванденхауте Дж., Зогби Х.И., Смоляр А., Босак С., Секерра Р., Дусетт-Стамм Л., Кьюсик М.Е., Хилл Д.Е., Рот Ф.П., Видал М. (2005). «К карте сети белок-белковых взаимодействий человека в масштабе протеома». Природа . 437 (7062): 1173–8. Бибкод : 2005Natur.437.1173R . дои : 10.1038/nature04209 . ПМИД 16189514 . S2CID 4427026 .
- Макдональд А., Кэмпбелл Д.Г., Тот Р., Маклаухлан Х., Хасти С.Дж., Артур Дж.С. (2006). «Киназы Pim фосфорилируют несколько сайтов на Bad и способствуют связыванию 14-3-3 и диссоциации от Bcl-XL» . BMC Клеточная Биол . 7 :1. дои : 10.1186/1471-2121-7-1 . ПМК 1368972 . ПМИД 16403219 .
Внешние ссылки
[ редактировать ]- Обзор всей структурной информации, доступной в PDB для UniProt : Q9P1W9 (серин/треонин-протеинкиназа pim-2) в PDBe-KB .
галерея PDB |
|---|
