Микробный симбиоз и иммунитет
Эта статья требует внимания эксперта в области молекулярной и клеточной биологии или микробиологии . Конкретная проблема заключается в следующем: содержание не сфокусировано, плохая структура, большая часть написана неопытным студентом-редактором. ( февраль 2017 г. ) |

Долгосрочные тесные взаимодействия между симбиотическими микробами и их хозяином могут изменить реакцию иммунной системы хозяина на другие микроорганизмы, включая патогены , и необходимы для поддержания надлежащего гомеостаза . [1] Иммунная система — это система защиты хозяина, состоящая из анатомических физических барьеров, а также физиологических и клеточных реакций, которые защищают хозяина от вредных микроорганизмов, одновременно ограничивая реакцию хозяина на безвредных симбионтов . У человека насчитывается 10 13 до 10 14 бактерий , примерно эквивалентных количеству клеток человека, [2] и хотя эти бактерии могут быть патогенными для своего хозяина, большинство из них взаимовыгодны как для хозяина, так и для бактерий.
Иммунная система человека состоит из двух основных типов иммунитета: врожденного и адаптивного. Врожденная иммунная система состоит из неспецифических защитных механизмов против чужеродных клеток внутри хозяина, включая кожу как физический барьер для проникновения, активацию каскада комплемента для идентификации чужеродных бактерий и активацию необходимых клеточных реакций, а также лейкоцитов , которые удаляют чужеродные вещества. . Адаптивная иммунная система , или приобретенная иммунная система, представляет собой патоген-специфический иммунный ответ, который осуществляется лимфоцитами посредством презентации антигена на молекулах MHC для различения собственных и чужих антигенов .
Микробы могут способствовать развитию иммунной системы хозяина в кишечнике и коже и могут помочь предотвратить патогенов проникновение . Некоторые выделяют противовоспалительные продукты, защищающие от паразитарных кишечных микробов. Комменсалы способствуют развитию В-клеток , которые вырабатывают защитные антитела — иммуноглобулин А (IgA). Это может нейтрализовать патогены и экзотоксины , а также способствовать развитию иммунных клеток и иммунного ответа слизистой оболочки. Однако микробы вовлечены в развитие заболеваний человека, включая воспалительные заболевания кишечника , ожирение и рак.
Общие принципы
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( март 2017 г. ) |
Микробный симбиоз основан на межвидовом общении . [3] между хозяином и микробными симбионтами. иммунитет определялся Исторически у многоклеточных организмов как контролируемый иммунной системой хозяина, когда воспринимаемое чужеродное вещество или клетка стимулирует иммунный ответ. Конечный результат этой реакции может варьироваться от уничтожения вредного патогена до толерантности к полезному микробу и аутоиммунной реакции , которая наносит вред самому хозяину.
Совсем недавно было показано, что симбиотические микроорганизмы также участвуют в этом иммунном ответе, что указывает на то, что иммунный ответ не ограничивается только клетками-хозяевами. Эти полезные микроорганизмы, помимо других реакций, участвуют в подавлении роста патогенов в кишечнике и противораковом иммунитете.
Желудочно-кишечный тракт
[ редактировать ]
человека Желудочно-кишечный тракт (ЖКТ) состоит из рта , глотки , пищевода , желудка , тонкой и толстой кишки и представляет собой непрерывную трубку длиной 9 метров; наибольшая площадь поверхности тела, подверженная воздействию внешней среды. Кишечник обеспечивает микробы питательными веществами и защитой, позволяя им процветать в кишечном микробном сообществе, состоящем из 10 14 полезные и патогенные бактерии , археи , вирусы и эукариоты . В свою очередь, многие из этих микробов выполняют важные функции для хозяина, включая расщепление волокон. [4] и производство витаминов [5] по крайней мере, роль в производстве витаминов, таких как A , B2 , B3 , B5 , B12 , C , D и K. где кишечные микробы играют ,
В кишечнике человека иммунная система контактирует с большим количеством чужеродных микробов, как полезных, так и патогенных. Иммунная система способна защитить хозяина от этих патогенных микробов, не вызывая ненужных и вредных иммунных реакций на раздражители. желудочно-кишечного тракта Микробиота оказывает прямое влияние на иммунные реакции организма человека. Это означает, что регулярная микробиота необходима для здоровой иммунной системы хозяина, поскольку организм более восприимчив к инфекционным и неинфекционным заболеваниям .
Регуляция иммунных ответов
[ редактировать ]Комменсальные бактерии в желудочно-кишечном тракте выживают, несмотря на обилие местных иммунных клеток. [6] Гомеостаз в кишечнике требует стимуляции толл-подобных рецепторов комменсальными микробами. [6] Когда мыши выращиваются в стерильных условиях, у них отсутствуют циркулирующие антитела, и они не могут производить слизь, антимикробные белки или Т-клетки слизистой оболочки. [6] Кроме того, мыши, выращенные в стерильных условиях, лишены толерантности и часто страдают от реакций гиперчувствительности . [6] Созревание желудочно-кишечного тракта опосредуется рецепторами распознавания образов (PRR) не связанные с собственными , которые распознают молекулярные паттерны, патогенами (PAMP), включая компоненты бактериальной клеточной стенки и нуклеиновые кислоты. [7] Эти данные свидетельствуют о том, что комменсальные микробы способствуют гомеостазу кишечника и развитию иммунной системы. [6]
Чтобы предотвратить постоянную активацию иммунных клеток и возникающее в результате воспаление, хозяева и бактерии эволюционировали, чтобы поддерживать гомеостаз кишечника и развитие иммунной системы. [8] Например, человеческий симбионт Bacteroides fragilis вырабатывает полисахарид А (PSA), который связывается с толл-подобным рецептором 2 (TLR-2) на CD4. + Т-клетки . [9] Хотя передача сигналов TLR2 может активировать клиренс пептидов, ПСА индуцирует противовоспалительный ответ, когда он связывается с TLR2 на CD4. + Т-клетки. [9] Посредством связывания TLR2 ПСА подавляет провоспалительные реакции TH17, способствуя толерантности и создавая комменсальную колонизацию кишечника. [9]
Комменсальные кишечные микробы создают множество метаболитов, которые связывают арилуглеводородные рецепторы (AHR) . AHR представляет собой индуцируемый лигандом фактор транскрипции, обнаруженный в иммунных и эпителиальных клетках, и связывание AHR необходимо для нормальной иммунной активации, поскольку было показано, что отсутствие связывания AHR вызывает чрезмерную активацию иммунных клеток. [1] Эти микробные метаболиты имеют решающее значение для защиты хозяина от ненужного воспаления в кишечнике.
Развитие изолированных лимфоидных тканей
[ редактировать ]Микробы вызывают развитие изолированных лимфоидных фолликулов в тонком кишечнике человека и мышей, которые являются участками иммунного ответа слизистой оболочки. Изолированные лимфоидные фолликулы (ILF) собирают антигены через М-клетки , развивают зародышевые центры и содержат множество В-клеток. [10] Грамотрицательные комменсальные бактерии запускают развитие индуцибельных лимфоидных фолликулов, высвобождая пептидогилканы, содержащие диаминопимелиновую кислоту, во время клеточного деления. [10] Пептидогликаны связываются с рецептором NOD1 на эпителиальных клетках кишечника . [10] В результате эпителиальные клетки кишечника экспрессируют хемокиновый лиганд 20 (CCL20) и бета-дефенсин 3 . [10] CCL20 и бета-дефенсин 3 активируют клетки, которые опосредуют развитие изолированных лимфоидных тканей, включая клетки-индукторы лимфоидной ткани и клетки-организаторы лимфоидной ткани. [10]
Кроме того, существуют и другие механизмы, с помощью которых комменсалы способствуют созреванию изолированных лимфоидных фолликулов. Например, продукты комменсальных бактерий связываются с TLR2 и TLR4 , что приводит к опосредованной NF-κB транскрипции TNF , которая необходима для созревания зрелых изолированных лимфоидных фолликулов. [11]
Защита от патогенов
[ редактировать ]Микробы могут предотвращать рост вредных патогенов, изменяя pH, потребляя питательные вещества, необходимые для выживания патогенов, и выделяя токсины и антитела, которые подавляют рост патогенов. [12]
Иммуноглобулин А
[ редактировать ]IgA предотвращает проникновение и колонизацию патогенных бактерий в кишечнике. Его можно найти в виде мономера, димера или тетрамера, что позволяет ему связывать несколько антигенов одновременно. [13] IgA покрывает патогенные бактериальные и вирусные поверхности (иммунное исключение), предотвращая колонизацию, блокируя их прикрепление к клеткам слизистой оболочки, а также может нейтрализовать PAMP. [8] [14] IgA способствует развитию регуляторных Т-клеток TH17 и FOXP3+. [15] [16] Учитывая его важнейшую функцию в желудочно-кишечном тракте, количество IgA-секретирующих плазматических клеток в тощей кишке превышает общую популяцию плазматических клеток костного мозга , лимфы и селезенки вместе взятых. [13]
Сигналы, полученные от микробиоты, привлекают плазматические клетки, секретирующие IgA, к участкам слизистой оболочки. [8] Например, бактерии на апикальных поверхностях эпителиальных клеток фагоцитируются дендритными клетками, расположенными под пейеровыми бляшками и в собственной пластинке слизистой оболочки , что в конечном итоге приводит к дифференцировке В-клеток в плазматические клетки, секретирующие IgA, специфичные для кишечных бактерий. [17] Роль сигналов микробиоты в привлечении IgA-секретирующих плазматических клеток была подтверждена в экспериментах с обработанными антибиотиками мышами, свободными от специфических патогенов, и мышами MyD88 KO , которые имеют ограниченное количество комменсалов и пониженную способность реагировать на комменсалов. Количество кишечных CD11b + IgA + У этих мышей было снижено количество плазматических клеток , что позволяет предположить роль комменсалов в привлечении IgA-секретирующих плазматических клеток. [18] Основываясь на этих данных, комменсальные микробы могут защищать хозяина от вредных патогенов, стимулируя выработку IgA.
Антимикробные пептиды
[ редактировать ]
Члены микробиоты способны вырабатывать антимикробные пептиды, защищающие человека от чрезмерного воспаления кишечника и заболеваний, связанных с микробами. Различные комменсалы (в первую очередь грамположительные бактерии ) секретируют бактериоцины , пептиды, которые связываются с рецепторами близкородственных клеток-мишеней, образуя проницаемые для ионов каналы и поры в клеточной стенке. [19] Возникающий в результате отток метаболитов и клеточного содержимого, а также рассеивание ионных градиентов вызывают гибель бактериальных клеток. [19] Однако бактериоцины также могут вызывать смерть, перемещаясь в периплазматическое пространство и неспецифически расщепляя ДНК (колицин Е2), инактивируя рибосому (колицин Е3), ингибируя синтез пептидогликана , основного компонента бактериальной клеточной стенки (колицин М). [19]
Бактериоцины обладают огромным потенциалом для лечения заболеваний человека. Например, диарея у людей может быть вызвана множеством факторов, но часто ее вызывают такие бактерии, как Clostridium difficile . [19] Штамм Microbispora ATCC PTA-5024 секретирует бактериоцин микробиспорицин, который убивает клостридии, воздействуя на синтез простагландинов . [20] Кроме того, бактериоцины особенно многообещающи из-за различий в механизмах их действия по сравнению с антибиотиками, что означает, что многие устойчивые к антибиотикам бактерии не устойчивы к этим бактериоцинам. Например, in vitro рост метициллин-резистентного S. aureus (MRSA) ингибировался бактериоцином низином А, продуцируемым Lactococcus Lactis . [19] [21] Низин А ингибирует устойчивый к метициллину S. aureus путем связывания с предшественником синтеза бактериальной клеточной стенки, липидом II . Это препятствует способности синтезировать клеточную стенку, что приводит к увеличению проницаемости мембран, нарушению электрохимических градиентов и возможной смерти. [22]
Обогащение фукозой
[ редактировать ]Эпителий кишечника у людей усилен углеводами, такими как фукоза, экспрессируемыми на апикальной поверхности эпителиальных клеток. [23] Bacteroides thetaiotaomicron , вид бактерий, обитающий в подвздошной и толстой кишке , стимулирует ген, кодирующий фукозу , Fut2, в эпителиальных клетках кишечника. [23] В этом мутуалистическом взаимодействии кишечный эпителиальный барьер укрепляется, и люди защищаются от вторжения разрушительных микробов, в то время как B. thetaiotaomicron получает выгоду, поскольку может использовать фукозу для производства энергии и ее роли в регуляции бактериальных генов. [23]
Кожа
[ редактировать ]
Микробиота кожи жизненно важна как линия защиты от инфекции, физический барьер между окружающей средой и внутренней частью организма хозяина. Комменсальные микробы, живущие на коже, такие как Staphylococcus epidermidis , производят антимикробные пептиды (АМП), которые помогают иммунной системе хозяина. [24] Эти АМФ сигнализируют об иммунных реакциях и поддерживают воспалительный гомеостаз , модулируя высвобождение цитокинов . [24] S. epidermidis секретирует небольшую молекулу АМФ, которая приводит к усилению экспрессии β-дефензинов человека. [24] S. epidermidis также стимулирует выработку IL-17A+ CD8+ Т-клеток, что повышает иммунитет хозяина. [25]
Воздействие этих кожных комменсальных бактерий на ранних стадиях развития имеет решающее значение для толерантности хозяина к этим микробам, поскольку встречи с Т-клетками делают презентацию комменсальных антигенов обычным явлением во время развития. [26] S. epidermidis и другая важная микрофлора аналогичным образом поддерживают гомеостаз и общее состояние здоровья во всех частях человеческого тела, таких как полость рта , влагалище , желудочно-кишечный тракт и ротоглотка . [24]
Роль в болезни
[ редактировать ]Равновесие симбионтов и патобионтов имеет решающее значение для борьбы с внешними патогенами и предотвращения многих вредных расстройств. Дисбиоз , или дисбаланс бактериального состава кишечника, связан с воспалительными заболеваниями кишечника, ожирением и аллергическими заболеваниями у людей и других животных. [27]
Рак
[ редактировать ]
Кишечные микробы могут играть роль в развитии рака посредством различных механизмов. Сульфатредуцирующие бактерии производят сероводород, что приводит к повреждению геномной ДНК . [28] Более высокие показатели рака толстой кишки связаны с большим количеством сульфатвосстанавливающих бактерий в кишечнике. [28] Кроме того, анаэробные бактерии в толстой кишке преобразуют первичные желчные кислоты во вторичные желчные кислоты, которые участвуют в колоректальном канцерогенезе. [28] кишечных бактерий, Метаболиты такие как короткоцепочечные жирные кислоты (КЦЖК) , витамины группы В и N. 1 , Н 12 -диацетилспермин также участвует в подавлении колоректального рака. [1] Грамотрицательные бактерии вырабатывают липополисахарид (ЛПС) , который связывается с TLR-4 и посредством передачи сигналов TGF-β приводит к экспрессии факторов роста и медиаторов воспаления, которые способствуют неоплазии . [28]
представители здорового кишечного микробиома Было показано, что интерферон-γ продуцирующих увеличивают количество Т-клеток CD8, , и инфильтрирующих опухоль дендритных клеток (TIL) в кишечнике. [29] Эти CD8 Т-клетки не только повышают устойчивость к внутриклеточным патогенам, таким как Listeria monocytogenes , но также было показано, что они играют важную роль в противораковом иммунитете, особенно против аденокарциномы MC38 , где они вместе с TIL увеличивают MHC I. экспрессию [29]
Аллергические и иммунные нарушения
[ редактировать ]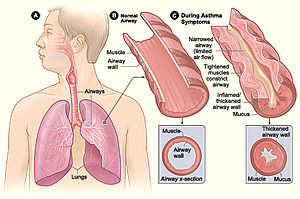
Микробиом человека позитивно модулирует иммунитет хозяина, помогая защититься от потенциальных патогенов, но также может привести к чрезмерным иммунным реакциям на чужеродные вещества, иногда даже атакуя самого хозяина. Воспалительные заболевания кишечника (ВЗК) и астма — это два заболевания, на которые, как было обнаружено, влияют метаболиты микробиоты, вызывающие иммунные реакции. Короткоцепочечные жирные кислоты (КЦЖК) связаны с уменьшением аллергического воспаления при астме. [30] в то время как было показано, что и SCFAs, и витамины группы B уменьшают воспаление воспалительного заболевания кишечника. [31]
SCFAs ( ацетат , бутират и пропионат ) представляют собой метаболиты, вырабатываемые бактериями в кишечнике. Эти молекулы затем ингибируют деацетилазы гистонов (HDAC), а также рецепторы, связанные с G-белком , действуя как сигнальные молекулы . [1] Ингибирование HDAC подавляет ядерный фактор-κB (NF-κB) и провоспалительный фактор некроза опухоли (TNF), а также оказывает противовоспалительное действие на макрофаги и дендритные клетки . [1]
Активация иммунитета слизистой оболочки и кишечной микробиоты может способствовать воспалительному заболеванию кишечника. Многие бактерии вызывают воспаление в кишечнике, включая Escherichia coli , которая реплицируется в макрофагах и секретирует цитокин фактор некроза опухоли. [32] Однако некоторые бактерии, в том числе симбионт человека B. fragilis, могут предотвращать колит , производя полисахарид А (ПСА). [33] ПСА индуцирует выработку IL-10 , иммуносупрессивного цитокина, подавляющего воспаление. [34] Лечение дендритных клеток костномозгового происхождения и наивного CD4 + Т-клетки с очищенным ПСА приводили к увеличению продукции IL-10. [34]
Чтобы имитировать колит и активировать воспалительные Т-клетки в экспериментальных условиях, мышей дикого типа лечили тринитробензолсульфоновой кислотой (TNBS). [34] После этого этим мышам вводили ПСА перорально. Экспрессия провоспалительных цитокинов ( IL-17a и TNFα ) в CD4 + клеток измеряли с помощью ELISA . Исследователи обнаружили, что по сравнению с CD4 + клетки контрольных мышей, CD4 + клетки у мышей, получавших ПСА, продуцировали сниженные уровни провоспалительных цитокинов IL-17a и TNFα. [34] Более того, после колонизации кишечника B. fragilis IL-23 экспрессия спленоцитами заметно снижалась. [34] Эти данные свидетельствуют о том, что ПСА, секретируемый B. fragilis, подавляет воспалительный процесс при колите, приводя к увеличению продукции IL-10 и снижению продукции IL-17, TNFα и IL-23. [34]
Комменсальные бактерии также могут регулировать иммунные реакции, вызывающие аллергию. Например, комменсальные бактерии стимулируют TLR4 , который может подавлять аллергические реакции на пищу. [35]
Метаболические нарушения
[ редактировать ]Было обнаружено, что метаболиты кишечной микробиоты влияют на основные метаболические заболевания, включая болезни сердца , заболевания почек , диабет 2 типа и ожирение . [1] Расщепление L-карнитина из красного мяса кишечными микробами на N-оксид триметиламина (ТМАО) связано с атеросклерозом , который может привести к ожирению, болезням сердца и диабету 2 типа. [36] в то время как события, связанные с заболеваниями сердца и почек, можно предсказать по высоким уровням свободного п-крезола . [37] SCFAs модулируют секрецию ренина путем связывания Olfr78, снижая кровяное давление и уменьшая риск заболеваний почек. [38]
Исследования на незараженных мышах показали, что отсутствие кишечных микробов защищает от ожирения. [39] Хотя точный механизм, посредством которого микробы играют роль в ожирении, еще предстоит выяснить, была выдвинута гипотеза, что кишечная микробиота участвует в преобразовании пищи в полезную энергию и накопление жира. [39]
Неврологические расстройства
[ редактировать ]Микробиота кишечника влияет на многие аспекты здоровья человека, даже на неврологические расстройства, которые могут быть вызваны дисбалансом молекул или гормонов. Расстройство аутистического спектра (РАС) , [1] дисфункция центральной нервной системы [1] и депрессия [40] Было обнаружено, что все они подвергаются воздействию микробиоты.
Хотя РАС обычно характеризуется поведенческими различиями, он также может проявляться желудочно-кишечными симптомами. [41] У некоторых лиц с РАС отмечен дисбиоз желудочно-кишечного тракта, приводящий к повышенной проницаемости кишечника. [41] У мышей-модельных мышей с РАС и дисбиозом желудочно-кишечного тракта (активация материнского иммунитета) было обнаружено повышенная проницаемость кишечника, что было скорректировано введением бактериального симбионта кишечника человека B. fragilis . [41]
Развитие микроглии играет ключевую роль в дисфункции центральной нервной системы, бактериальные метаболиты SCFAs регулируют гомеостаз микроглии , что имеет решающее значение для нормального развития ЦНС. [42] Также решающее значение для развития мозга имеет создание плотных соединений на гематоэнцефалическом барьере , чтобы контролировать проход между кровью и мозгом. Безмикробные мыши имеют повышенную проницаемость гематоэнцефалического барьера из-за снижения экспрессии белков плотных соединений окклюдина и клаудина-5 по сравнению с мышами с нормальной микробиотой кишечника. [43]
бутират Бактерии, продуцирующие дофамина , и метаболит 3,4-дигидроксифенилуксусная кислота связаны с более высокими показателями качества жизни, а γ-аминомасляная кислота связана с более высоким уровнем депрессии. [40]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час Рукс, Мишель Г.; Гарретт, Венди С. (27 мая 2016 г.). «Кишечная микробиота, метаболиты и иммунитет хозяина» . Обзоры природы Иммунология . 16 (6): 341–352. дои : 10.1038/nri.2016.42 . ISSN 1474-1733 . ПМЦ 5541232 . ПМИД 27231050 .
- ^ Мазманян, Саркис (2006). «Отношения любви и ненависти между бактериальными полисахаридами и иммунной системой хозяина». Обзоры природы Иммунология . 849–858 (11): 849–858. дои : 10.1038/nri1956 . ПМИД 17024229 . S2CID 20380038 .
- ^ Маккенни Дэвид; Браун Кэтрин; Эллисон Дэвид (1995). «Влияние экзопродуктов Pseudomonas aeruginosa на продукцию факторов вирулентности у Burkholderia cepacia: доказательства межвидовой связи» . Журнал бактериологии . 177 (23): 6989–6991. дои : 10.1128/jb.177.23.6989-6992.1995 . ПМК 177571 . ПМИД 7592496 .
- ^ Холшер, Ханна Д. (04 марта 2017 г.). «Пищевая клетчатка, пребиотики и микробиота желудочно-кишечного тракта» . Кишечные микробы . 8 (2): 172–184. дои : 10.1080/19490976.2017.1290756 . ISSN 1949-0984 . ПМК 5390821 . ПМИД 28165863 .
- ^ Леблан, Жан Ги; Милани, Кристиан; де Джори, Грасиела Савойя; Сесма, Фернандо; ван Синдерен, Доуве; Вентура, Марко (апрель 2013 г.). «Бактерии как поставщики витаминов для своего хозяина: взгляд на кишечную микробиоту». Современное мнение в области биотехнологии . 24 (2): 160–168. дои : 10.1016/j.copbio.2012.08.005 . hdl : 11336/2561 . ISSN 1879-0429 . ПМИД 22940212 .
- ^ Jump up to: а б с д и Браун, Э.М. (2013). «Свежий взгляд на гигиеническую гипотезу: как воздействие микробов в кишечнике влияет на иммунные эффекторные реакции при атопическом заболевании». Семинары по иммунологии . 25 (5): 378–387. дои : 10.1016/j.smim.2013.09.003 . ПМИД 24209708 .
- ^ Палм, Ной В.; де Зете, Марсель Р.; Флавелл, Ричард А. (август 2015 г.). «Взаимодействие иммунитета и микробиоты в здоровье и болезни» . Клиническая иммунология . 159 (2): 122–127. дои : 10.1016/j.clim.2015.05.014 . ISSN 1521-6616 . ПМЦ 4943041 . ПМИД 26141651 .
- ^ Jump up to: а б с Серф-Бенсуссан, Надин; Габорио-Рутио, Валери (01 октября 2010 г.). «Иммунная система и микробиота кишечника: друзья или враги?». Обзоры природы Иммунология . 10 (10): 735–744. дои : 10.1038/nri2850 . ПМИД 20865020 . S2CID 13257259 .
- ^ Jump up to: а б с Раунд, Джун Л.; Ли, С. Мелани; Ли, Дженнифер; Тран, Глория; Джабри, Бана; Чатила, Талал А.; Мазманян, Саркис К. (20 мая 2011 г.). «Путь Toll-подобных рецепторов обеспечивает комменсальную колонизацию кишечника» . Наука . 332 (6032): 974–977. дои : 10.1126/science.1206095 . ПМК 3164325 . ПМИД 21512004 .
- ^ Jump up to: а б с д и Эберл, Г.; Лохнер, М. (9 сентября 2009 г.). «Развитие лимфоидной ткани кишечника на стыке личности и микробиоты» . Иммунология слизистой оболочки . 2 (6): 478–485. дои : 10.1038/ми.2009.114 . ПМИД 19741595 .
- ^ Бускра, Джахида; Брезийон, Кристоф; Берар, Марион; Вертс, Кэтрин; Варона, Роза; Бонека, Иво Гомпертс; Эберль, Жерар (27 ноября 2008 г.). «Генезис лимфоидной ткани, индуцированный комменсалами через NOD1, регулирует гомеостаз кишечника». Природа . 456 (7221): 507–510. Бибкод : 2008Natur.456..507B . дои : 10.1038/nature07450 . ПМИД 18987631 . S2CID 205215248 .
- ^ Камада, Н. (2013). «Контроль патогенов и патобионтов микробиотой кишечника» . Природная иммунология . 14 (7): 685–690. дои : 10.1038/ni.2608 . ПМЦ 4083503 . ПМИД 23778796 .
- ^ Jump up to: а б Киндт, Томас Дж.; Голдсби, Ричард А.; Осборн, Барбара А.; Куби, Янис (23 октября 2006 г.). Куби Иммунология . У. Х. Фриман. стр. 90–92. ISBN 9781429203944 .
- ^ Мантис, Нью-Джерси; Рол, Н.; Кортези, Б. (1 ноября 2011 г.). «Сложная роль секреторного IgA в иммунитете и гомеостазе слизистой оболочки кишечника» . Иммунология слизистой оболочки . 4 (6): 603–611. дои : 10.1038/ми.2011.41 . ПМЦ 3774538 . ПМИД 21975936 .
- ^ Макферсон, Эй Джей (2008). «Иммунная география индукции и функции IgA» . Иммунология слизистой оболочки . 1 (1): 11–22. дои : 10.1038/ми.2007.6 . ПМИД 19079156 .
- ^ Камада, Н. (2013). «Роль микробиоты кишечника в иммунитете и воспалительных заболеваниях». Обзоры природы Иммунология . 13 (5): 321–335. дои : 10.1038/nri3430 . ПМИД 23618829 . S2CID 205491968 .
- ^ Хупер Лора В., Брай Линн, Фальк Пер Г., Гордон Джеффри И. (1998). «Симбиоз хозяина и микроба в кишечнике млекопитающих: исследование внутренней экосистемы». Биоэссе . 20 (4): 336–343. doi : 10.1002/(sici)1521-1878(199804)20:4<336::aid-bies10>3.3.co;2-j . ПМИД 9619105 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Кунисава, Джун; Года, Масаси; Хашимото, Эри; Исикава, Идзуми; Хигучи, Морио; Сузуки, Юджи; Гото, Ёсиюки; Панеа, Касандра; Иванов, Ивайло И. (23 апреля 2013 г.). «Микробно-зависимые CD11b+ IgA+ плазматические клетки опосредуют устойчивые реакции IgA кишечника на ранней стадии у мышей» . Природные коммуникации . 4 : 1772. Бибкод : 2013NatCo...4.1772K . дои : 10.1038/ncomms2718 . ПМК 3644083 . ПМИД 23612313 .
- ^ Jump up to: а б с д и Хаммами, Риад; Фернандес, Бенуа; Лакруа, Кристоф; Флисс, Исмаил (30 октября 2012 г.). «Противоинфекционные свойства бактериоцинов: обновленная информация» . Клеточные и молекулярные науки о жизни . 70 (16): 2947–2967. дои : 10.1007/s00018-012-1202-3 . ПМЦ 11113238 . ПМИД 23109101 . S2CID 16228657 .
- ^ Кастильоне, Франка; Лаццарини, Америка; Каррано, Люсия; Корти, Эмилиана; Чичильято, Исмаэла; Гастальдо, Лучано; Кандиани, Паоло; Лоси, Даниэле; Маринелли, Флавия (25 января 2008 г.). «Определение структуры и механизма действия микробиспорицина, мощного лантибиотика, активного против полирезистентных патогенов» . Химия и биология . 15 (1): 22–31. doi : 10.1016/j.chembiol.2007.11.009 . ПМИД 18215770 .
- ^ Пайпер, К.; Дрейпер, Луизиана; Коттер, PD; Росс, РП; Хилл, К. (1 сентября 2009 г.). «Сравнение активности лакцина 3147 и низина против лекарственно-устойчивых видов Staphylococcus aureus и Enterococcus» . Журнал антимикробной химиотерапии . 64 (3): 546–551. дои : 10.1093/jac/dkp221 . ПМИД 19561147 .
- ^ Сюй, Шан-Те Д.; Брейкинк, Эфьян; Тищенко Евгений; Латтерс, Мэнди АГ; де Крейфф, Бен; Каптейн, Роберт; Бонвен, Александр MJJ; ван Нуланд, Нико Эй Джей (01 октября 2004 г.). «Комплекс низин-липид II обнаруживает пирофосфатную клетку, которая обеспечивает основу для новых антибиотиков». Структурная и молекулярная биология природы . 11 (10): 963–967. дои : 10.1038/nsmb830 . hdl : 1874/385449 . ПМИД 15361862 . S2CID 13181577 .
- ^ Jump up to: а б с Гото, Ёсиюки; Киёно, Хироши (2012). «Эпителиальный барьер: интерфейс для перекрестной связи между кишечной флорой и иммунной системой». Иммунологические обзоры . 245 (1): 147–163. дои : 10.1111/j.1600-065X.2011.01078.x . ПМИД 22168418 . S2CID 22187069 .
- ^ Jump up to: а б с д Галло Ричард Л., Накацудзи Теруаки (2011). «Микробный симбиоз с системой врожденной иммунной защиты кожи» . Журнал исследовательской дерматологии . 131 (10): 1974–1980. дои : 10.1038/jid.2011.182 . ПМК 3174284 . ПМИД 21697881 .
- ^ Наик, Шрути ; Буладу, Николя; Линехан, Джонатан Л.; Хан, Сон Джи; Харрисон, Оливер Дж.; Вильгельм, Кристоф; Конлан, Шон; Химмельфарб, Сара; Берд, Эллисон Л.; Деминг, Клейтон; Хиноны, Мариам (2 апреля 2015 г.). «Взаимодействие комменсала и дендритных клеток определяет уникальную защитную иммунную сигнатуру кожи» . Природа . 520 (7545): 104–108. Бибкод : 2015Natur.520..104N . дои : 10.1038/nature14052 . ISSN 0028-0836 . ПМЦ 4667810 . ПМИД 25539086 .
- ^ Шаршмидт, Тиффани К. (январь 2017 г.). «Установление толерантности к комменсальным бактериям кожи: время решает все» . Дерматологические клиники . 35 (1): 1–9. дои : 10.1016/j.det.2016.07.007 . ISSN 0733-8635 . ПМК 5130113 . ПМИД 27890233 .
- ^ ДеГруттола, Арианна К.; Лоу, Дарен; Мидзогучи, Ацуши; Мидзогучи, Эмико (25 февраля 2017 г.). «Современное понимание дисбактериоза при заболеваниях на моделях человека и животных» . Воспалительные заболевания кишечника . 22 (5): 1137–1150. doi : 10.1097/MIB.0000000000000750 . ПМЦ 4838534 . ПМИД 27070911 .
- ^ Jump up to: а б с д Халлар, Мередит Эй Джей; Бернетт-Хартман, Андреа Н.; Лампе, Йоханна В. (1 января 2014 г.). «Кишечные микробы, диета и рак». Достижения в области питания и рака . Лечение и исследования рака. Том. 159. стр. 377–399. дои : 10.1007/978-3-642-38007-5_22 . ISBN 978-3-642-38006-8 . ISSN 0927-3042 . ПМК 4121395 . ПМИД 24114492 .
- ^ Jump up to: а б Таноуэ, Такеши; Морита, Сатору; Плихта, Дамиан Р.; Скелли, Эшвин Н.; Суда, Ватару; Сугиура, Юки; Нарушима, Сейко; Вламакис, Гера; Мотоо, Иори; Сугита, Кайоко; Сиота, Ацуши (январь 2019 г.). «Определенный комменсальный консорциум вызывает Т-клетки CD8 и противораковый иммунитет». Природа . 565 (7741): 600–605. Бибкод : 2019Natur.565..600T . дои : 10.1038/s41586-019-0878-z . ISSN 1476-4687 . ПМИД 30675064 . S2CID 59159425 .
- ^ Мин, Буки (18 февраля 2014 г.). «Оценка факультета 1000 микробиоты кишечника, метаболизм пищевых волокон влияет на аллергические заболевания дыхательных путей и кроветворение» . дои : 10.3410/ф.718228193.793491060 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Сингх, Нагендра; Гурав, Ашиш; Шивапракасам, Сатиш; Брэди, Эван; Падия, Рави; Ши, Хуэйдун; Тангараджу, Мутусами; Прасад, Путтур Д.; Маникассами, Сантакумар; Манн, Дэвид Х.; Ли, Джеффри Р. (январь 2014 г.). «Активация Gpr109a, рецептора ниацина и комменсального метаболита бутирата, подавляет воспаление толстой кишки и канцерогенез» . Иммунитет . 40 (1): 128–139. doi : 10.1016/j.immuni.2013.12.007 . ISSN 1074-7613 . ПМК 4305274 . ПМИД 24412617 .
- ^ Сартор, Р. Бальфур; Мазманян, Саркис К. (01 июля 2012 г.). «Кишечные микробы при воспалительных заболеваниях кишечника» . Американский журнал гастроэнтерологических добавок . 1 (1): 15–21. дои : 10.1038/ajgsup.2012.4 .
- ^ Раунд, Джун Л.; Мазманян, Саркис К. (16 февраля 2017 г.). «Микробиом кишечника формирует иммунные реакции кишечника во время здоровья и болезней» . Обзоры природы Иммунология . 9 (5): 313–323. дои : 10.1038/nri2515 . ПМК 4095778 . ПМИД 19343057 .
- ^ Jump up to: а б с д и ж Мазманян, Саркис К.; Раунд, Джун Л.; Каспер, Деннис Л. (2008). «Фактор микробного симбиоза предотвращает воспалительные заболевания кишечника». Природа . 453 (7195): 620–625. Бибкод : 2008Natur.453..620M . дои : 10.1038/nature07008 . ПМИД 18509436 . S2CID 205213521 .
- ^ Башир, Мохамед Эльфатих Х.; Луи, Стив; Ши, Хай Нин; Наглер-Андерсон, Кэтрин (1 июня 2004 г.). «Передача сигналов Toll-подобного рецептора 4 кишечными микробами влияет на восприимчивость к пищевой аллергии» . Журнал иммунологии . 172 (11): 6978–6987. doi : 10.4049/jimmunol.172.11.6978 . ПМИД 15153518 .
- ^ Терри, Пол; Чен, Цзянган (11 апреля 2013 г.). «Оценка факультета 1000 кишечной микробиоты: метаболизм L-карнитина, питательного вещества в красном мясе, способствует развитию атеросклероза» . дои : 10.3410/f.717998892.793474469 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Мейерс, Бьёрн К.И.; Клаас, Кэтлин; Бамменс, Берт; де Лор, Генриетта; Виэна, Лисбет; Вербеке, Кристин; Кайперс, Дирк; Ванрентергем, Ив; Эвенепол, Питер (июль 2010 г.). «П-крезол и сердечно-сосудистый риск при легкой и умеренной болезни почек» . Клинический журнал Американского общества нефрологов . 5 (7): 1182–1189. дои : 10.2215/CJN.07971109 . ISSN 1555-9041 . ПМЦ 2893077 . ПМИД 20430946 .
- ^ Перссон, Эрик Г; Карлстрем, Маттиас (21 мая 2013 г.). «Оценка факультета 1000 обонятельных рецепторов, реагирующих на сигналы кишечной микробиоты, играет роль в секреции ренина и регуляции артериального давления» . дои : 10.3410/f.717978217.793476920 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Jump up to: а б Кардинг, Саймон; Вербеке, Кристин; Випонд, Дэниел Т.; Корф, Бернард М.; Оуэн, Лорен Дж. (2 февраля 2015 г.). «Дисбиоз микробиоты кишечника при заболевании» . Микробная экология в здоровье и болезнях . 26 : 26191. дои : 10.3402/mehd.v26.26191 . ISSN 0891-060X . ПМЦ 4315779 . ПМИД 25651997 .
- ^ Jump up to: а б Валлес-Коломер, Мирейя; Фалони, Гвен; Дарзи, Юсеф; Тигчелаар, Эттье Ф.; Ван, Цзюнь; Тито, Рауль Ю.; Шивек, Кармен; Курильщиков, Александр; Йоссенс, Мари; Вейменга, Сиска; Клаас, Стефан (апрель 2019 г.). «Нейроактивный потенциал микробиоты кишечника человека в отношении качества жизни и депрессии» (PDF) . Природная микробиология . 4 (4): 623–632. дои : 10.1038/s41564-018-0337-x . ISSN 2058-5276 . ПМИД 30718848 . S2CID 59603019 .
- ^ Jump up to: а б с Сяо, Элейн Ю.; Макбрайд, Сара В.; Сянь, София; Шэрон, Гил; Хайд, Эмбриетт Р.; МакКью, Тайлер; Коделли, Джулиан А.; Чоу, Джанет; Райсман, Сара Э.; Петрозино, Джозеф Ф.; Паттерсон, Пол Х. (19 декабря 2013 г.). «Микробиота модулирует физиологию кишечника и поведенческие отклонения, связанные с аутизмом» . Клетка . 155 (7): 1451–1463. дои : 10.1016/j.cell.2013.11.024 . ISSN 0092-8674 . ПМЦ 3897394 . ПМИД 24315484 .
- ^ Давидович, Летиция (22 июня 2015 г.). «Факультет 1000 оценки микробиоты хозяина постоянно контролирует созревание и функцию микроглии в ЦНС» . дои : 10.3410/f.725528105.793507643 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Браниште, Виорика; Аль-Асмах, Маха; Коваль, Чеслава; Ануар, Фархана; Аббаспур, Афруз; Тот, Миклош; Корецка, Агата; Бакочевич, Надя; Нг, Лай Гуань; Кунду, Параг; Гуляш, Балаж (19 ноября 2014 г.). «Микробиота кишечника влияет на проницаемость гематоэнцефалического барьера у мышей» . Наука трансляционной медицины . 6 (263): 263ра158. doi : 10.1126/scitranslmed.3009759 . ISSN 1946-6234 . ПМЦ 4396848 . ПМИД 25411471 .