Жерминальный центр
Эта статья нуждается в дополнительных цитатах для проверки . ( май 2016 г. ) |
| Жерминальный центр | |
|---|---|
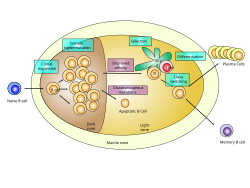 Зародышевый центр лимфатического узла, показывающий стадии пролиферации и развития B-клетки. | |
| Идентификаторы | |
| МеШ | D018858 |
| Анатомическая терминология | |
Зародышевые центры или зародышевые центры ( ГЦ ) представляют собой временно образующиеся структуры в зоне В-клеток (фолликулах) во вторичных лимфоидных органах – лимфатических узлах , пейеровых бляшках подвздошной кишки и селезенке . [1] – где зрелые В-клетки активируются, пролиферируют, дифференцируются и мутируют свои антител гены (посредством соматической гипермутации , направленной на достижение более высокого сродства) во время нормального иммунного ответа; большая часть В-клеток зародышевого центра (BGC ) удаляется макрофагами тела . [2] Существует несколько ключевых различий между наивными B-клетками и GC B-клетками, включая уровень пролиферативной активности, размер, метаболическую активность и производство энергии. [3] В-клетки динамически развиваются после активации фолликулярных В-клеток Т-зависимым антигеном . Инициация формирования зародышевого центра включает взаимодействие между В- и Т-клетками в межфолликулярной области лимфатического узла, лигирование CD40-CD40L, передачу сигналов NF-kB и экспрессию IRF4 и BCL6 . [4]
GC B-клетки циклически проходят через две отдельные зоны зародышевого центра: светлую зону и темную зону. [3] [4] [5] [6] Поскольку В-клетки темной зоны зародышевого центра подвергаются быстрому и мутационному клеточному делению, они известны как центробласты . Как только эти B-клетки перестают пролиферировать в темной зоне и перемещаются в светлую зону, они называются центроцитами и подвергаются селекции фолликулярными Т-хелперными клетками (T FH ) в присутствии фолликулярных дендритных клеток (FDC). [3] [4] [5] [6] Существует три возможных судьбы GC B-клеток, которые были положительно отобраны в светлой зоне: плазматическая клетка, B-клетка памяти или B-клетка, получившая лицензию на возвращение в темную зону для пролиферации и мутации. [4] [6] Эти три судьбы достигаются с помощью различных механизмов, описанных ниже. Герминальные центры являются важной частью гуморального иммунного ответа В-клеток , действуя как центральные фабрики для генерации созревших по аффинности В-клеток, специализирующихся на производстве улучшенных антител, которые эффективно распознают антиген (например, инфекционные агенты), а также на производство долгоживущей плазмы. клетки и В-клетки долговременной памяти .
Наивные В-клетки против В-клеток зародышевого центра
[ редактировать ]Существует несколько ключевых различий между наивными B-клетками и GC B-клетками. Наивные В-клетки не подвергаются интенсивному клеточному делению. С другой стороны, B-клетки в GC имеют тенденцию быстро и часто делиться, и их клеточный цикл может составлять всего пять часов. В результате высокой пролиферативности GC B-клетки больше по размеру и более метаболически активны по сравнению с наивными B-клетками. Хотя GC B-клетки имеют большую потребность в энергии, чем наивные B-клетки, они в основном производят энергию в процессе окисления жирных кислот , тогда как наивные B-клетки зависят от гликолиза . [3]
Инициация зародышевого центра
[ редактировать ]Зародышевые центры инициируются в В-клеточном фолликуле лимфатического узла . После активации наивных В-клеток в фолликулах лимфатических узлов В-клетки мигрируют в межфолликулярные области, где они могут взаимодействовать с Т-клетками. Когда В- и Т-клетки взаимодействуют, антигенспецифические рецепторы Т-клеток связывают антиген + MHC, представленный В-клетками. Кроме того, Т-клетки способны помогать В-клеткам за счет взаимодействия лиганда CD40 Т-клеток с молекулой CD40 В-клеток , что вызывает сигнальный каскад , полезный для выживания и пролиферации В-клеток. Активация В-клеточного рецептора приводит к активации сигнального пути NF-kB , который необходим для инициации реакции зародышевого центра. В частности, экспрессия факторов транскрипции IRF4 и BCL6 необходима для развития зародышевого центра и регулируется передачей сигналов NF-kB. Например, BCL6 контролирует расположение B-клеток в лимфатическом узле и позволяет им иметь более высокую толерантность к повреждению ДНК, тем самым способствуя пролиферации GC B-клеток. Все В-клетки начинаются с совместной экспрессии антител, Константные области IgM и IgD , но позже они способны обменивать эти константные области на константные области IgA , IgG или IgE и экспрессировать антитела другого типа класса посредством рекомбинации переключения классов. Рекомбинация переключения классов происходит во время фазы инициации зародышевого центра. Предшественники В-клеток зародышевого центра начинают размножаться через четыре дня после иммунизации и поляризуются на темные и светлые зоны через неделю после иммунизации. [4]
Две отдельные зоны зародышевого центра: темная зона и светлая зона.
[ редактировать ]Есть две отдельные области зародышевого центра: светлая зона (LZ) и темная зона (DZ). [3] [4] [5] [6] Эти две зоны образуются из В-клеток pre-GC, которые пролиферируют и поляризуются через семь дней после иммунизации. [3] [4] GC B-клетки чередуются между темной зоной и светлой зоной и подвергаются нескольким раундам мутации и селекции соответственно. [5] [6]
Темная зона
[ редактировать ]Темная зона зародышевого центра находится проксимальнее зоны Т-клеток в лимфатическом узле и состоит из GC B-клеток и ретикулярных клеток , которые напоминают фолликулярные дендритные клетки . [3] В-клетки в темной зоне зародышевого центра называются центробластами . [3] Они крупнее клеток светлой зоны зародышевого центра и более пролиферативны (т.е. подвергаются большему клеточному делению). [3] [5] В темной зоне происходит соматическая гипермутация — процесс, при котором индуцируемый активацией фермент цитидиндезаминаза (AID) случайным образом мутирует вариабельные области антитела и изменяет их сродство к антигену. [3] [4] [5] [6] Кроме того, В-клетки, которые были положительно отобраны в светлой зоне, поскольку они экспрессируют В-клеточные рецепторы с высоким сродством к антигену, активно пролиферируют в темной зоне, что представляет собой процесс, называемый клональной экспансией. [3] [6] После соматической гипермутации и перед входом в световую зону старые рецепторы В-клеток на поверхности В-клеток заменяются новыми, мутировавшими рецепторами В-клеток. [4] В-клетки, экспрессирующие антитела, у которых после соматической гипермутации снижено сродство к антигену, подвергаются апоптозу , тогда как В-клетки, экспрессирующие антитела, обладающие повышенным сродством к антигену после соматической гипермутации, мигрируют в световую зону для дальнейшей селекции. [4]
Светлая зона
[ редактировать ]Светлая зона состоит из GC B-клеток и Т-фолликулярных хелперных клеток . [3] Он расположен проксимальнее лимфатического узла и рядом с сетью фолликулярных дендритных клеток . [3] GC B-клетки в светлой зоне, известные как центроциты , меньше по размеру, менее многочисленны и меньше делятся по сравнению с GC B-клетками в темной зоне. [3] [4] [5] Соседние фолликулярные дендритные клетки представляют антиген GC B-клеткам светлой зоны, которые ранее мутировали в темной зоне, а клетки с самым высоким сродством к антигену способны связываться и получать помощь от Т-фолликулярных хелперных клеток, которые имеют Т-клеточные рецепторы. специфичны для одного и того же антигена. [3] [4] [5] [6] Следовательно, GC B-клетки в светлой зоне конкурируют за антиген и стимуляцию Т-фолликулярными хелперными клетками. [3] [4] [5] [6] Механизм, посредством которого это происходит, заключается в том, что, когда рецептор В-клеток связывает антиген, представленный фолликулярными дендритными клетками, антиген интернализуется. Затем антиген связывается с MHC класса II и презентируется на поверхности Т-клетки, что позволяет В-клетке оказывать помощь Т-фолликулярным хелперным клеткам. [6] GC B-клетки, которые лучше всего способны представлять антиген Т-фолликулярным хелперным клеткам и продуцировать самый сильный сигнал рецептора B-клеток, положительно отбираются в светлой зоне зародышевого центра. [4] Таким образом, положительная селекция GC B-клеток в светлой зоне приводит к появлению B-клеток, которые экспрессируют антитела с высоким сродством к антигену. [3] В-клетки, прошедшие положительную селекцию в светлой зоне, начинают экспрессировать cMyc , который регулирует зародышевый центр и пролиферацию В-клеток в зародышевом центре. [3] Наконец, положительно отобранные GC B-клетки (cMyc + ) «лицензированы», что означает, что они готовы быть отправлены обратно в темную зону зародышевого центра, где они будут дальше размножаться и мутировать путем соматической гипермутации . [6]
Процесс
[ редактировать ]

- Центроциты от малого до среднего размера с угловатыми, удлиненными, расщепленными или скрученными ядрами.
- Центробласты – это более крупные клетки, содержащие везикулярные ядра с одним-тремя базофильными ядрышками, прилегающими к ядерной мембране.
- Фолликулярные дендритные клетки имеют округлые ядра, центрально расположенные ядрышки, мягкий и рассеянный хроматин и уплощенную прилежащую ядерную мембрану.
- Внутри лимфатических узлов зрелые периферические В-клетки, известные как фолликулярные (Fo) В-клетки, приобретают антиген из FDC и, в свою очередь, представляют его родственным CD4 + Т FH -клеткам на границе, которая разграничивает межфолликулярную область Т-клеток и зону В-клеток (также известную как лимфоидные фолликулы).
- После нескольких циклов деления клеток В-клетки подвергаются соматической гипермутации — процессу, в ходе которого они мутируют свою антитела , кодирующую ДНК , и таким образом генерируют множество клонов в зародышевом центре. Это включает псевдослучайные замены, смещенные в сторону областей, кодирующих поверхность распознавания антигенов антител, которые продуцируют В-клетки. Это явление подчеркивает процесс созревания аффинности , в результате которого вырабатываются и отбираются антитела с большей аффинностью после распознавания антигена.
- При получении неопознанного стимула созревающие В-клетки (центробласты) мигрируют из темной зоны в светлую зону и начинают экспрессировать свои отредактированные BCR на поверхности клетки и на этом этапе называются центроцитами . Центроциты находятся в состоянии активированного апоптоза и конкурируют за сигналы выживания, полученные от FDC и Т -ФГ- клеток . Считается, что этот процесс спасения, известный как отбор зародышевых центров, зависит от сродства их поверхностных антител к антигену. Таким образом, В-клетка, которая успешно получила мутации, которые придают поверхностным антителам с более высоким сродством к антигену, получает преимущество в выживании по сравнению с клонами В-клеток с более низким сродством и теми, которые получили вредные мутации. Циклический повторный вход в темную зону в виде центробластов дает шанс невыбранным в противном случае мутантам В-клеток получить больше мутаций, чтобы улучшить сродство к антигену. Считается также, что взаимодействие с Т-клетками предотвращает образование аутореактивных В-клеток зародышевого центра. [7]
- На какой-то неясной стадии цикла центробласт-центроциты созревающие В-клетки получают окончательный сигнал дифференцировки для выхода из зародышевого центра в виде плазматических клеток , продуцирующих антитела , которые представляют собой клетки, секретирующие большие количества антител, или В-клетки памяти , которые могут быть реактивированы в последующем. контакты с одним и тем же антигеном. Выбранные В-клетки также могут перезапустить весь цикл мутативного деления центробластов и отбора центроцитов. Таким образом, адаптивная иммунная система, отчасти благодаря реакциям зародышевого центра, со временем может постепенно лучше распознавать антигены.
Роль Т-фолликулярных хелперных клеток в зародышевом центре
[ редактировать ]имеются Т-хелперные клетки, называемые Т-фолликулярными хелперными клетками, которые способствуют образованию зародышевого центра и дифференцировке GC B-клеток в плазматические клетки и B-клетки памяти. В фолликулах лимфатических узлов [5] Т-фолликулярные хелперные клетки опосредуют реакцию зародышевого центра двумя ключевыми способами. Во-первых, Т-фолликулярные хелперные клетки экспрессируют CD40L , который представляет собой цитокин фактора некроза опухоли (TNF), который связывает молекулу CD40, экспрессируемую на GC B-клетках. Это взаимодействие активирует сигнальный путь NF-kB, который стимулирует деление GC B-клеток. Во-вторых, Т-фолликулярные хелперные клетки секретируют цитокин IL-21 , который служит сигналом для пролиферации GC B-клеток и создания плазматических клеток с длительной продолжительностью жизни. [3] [5]
Судьба положительно отобранных В-клеток зародышевого центра
[ редактировать ]После положительного отбора есть три возможных судьбы В-клеток, подвергающихся реакции зародышевого центра: стать плазматической клеткой , стать В-клеткой памяти или войти в темную зону зародышевого центра. [4] [6] Процессы, инициирующие каждую из этих трёх судеб, описаны ниже:
Дифференциация плазматических клеток
[ редактировать ]GC B-клетки, которые дифференцируются в плазматические клетки, представляют собой B-клетки, демонстрирующие высокое сродство к антигену. [3] [6] Когда GC B-клетки получают помощь от Т-фолликулярных хелперных клеток, происходит взаимодействие между CD40 (экспрессируемым на В-клетке) и CD40L (экспрессируемым на Т-фолликулярных хелперных клетках), что увеличивает активацию NF-kB в В-клетке. Повышение регуляции сигнального пути NF-kB приводит к большей экспрессии IRF4, фактора транскрипции, который необходим для дифференцировки плазматических клеток. [6] Прогрессирование реакции зародышевого центра приводит к тому, что плазматические клетки, секретирующие антитела с более высоким сродством, имеют увеличенную продолжительность жизни и отправляются в костный мозг . [5]
Дифференциация B-клеток памяти
[ редактировать ]GC B-клетки, которые дифференцируются в B-клетки памяти , отличаются от предшественников плазматических клеток, поскольку они демонстрируют более низкое сродство к антигену. [3] [6] и не нуждаются в особой помощи со стороны Т-фолликулярных хелперных клеток. Из-за этого многие ученые полагают, что предшественниками В-клеток памяти являются В-клетки из светлой зоны, которые были «неположительно отобраны». Предшественники В-клеток памяти экспрессируют транскрипционный фактор, называемый гематопоэтически экспрессируемым гомеобоксным белком (Hhex) , который управляет дифференцировкой В-клеток памяти из GC В-клеток. [6]
Вход в темную зону зародышевого центра
[ редактировать ]Любые В-клетки, которые были положительно отобраны в светлой зоне зародышевого центра, но не дифференцировались в плазматические клетки или В-клетки памяти, отправляются в темную зону зародышевого центра для дальнейшей пролиферации. Это В-клетки, обладающие промежуточным сродством к антигену. [3] Программа пролиферации темной зоны регулируется FoxO1 и циклином D3 . Эти два гена подавляются сильными сигналами BCR. Следовательно, когда присутствуют слабые сигналы BCR и GC B-клетка не имеет высокого сродства к антигену, она будет отправлена в темную зону зародышевого центра, чтобы она могла продолжать делиться, а не секретироваться как плазматическая клетка или В-клетка памяти. [6]
Морфология на разных стадиях
[ редактировать ]Морфология ГЦ весьма специфична и проявляет свойства , характерные для разных стадий реакции.
- На ранней стадии реакции сеть ФДК полностью заполнена пролиферирующими В-клетками.
- Позже, на 4-й день реакции, ГХ показывают разделение на две зоны: темную и светлую. [8] Первый по-прежнему содержит преимущественно пролиферирующие и мутирующие В-клетки, а второй является областью селекции В-клеток.
- Эти зоны рассасываются через 10 дней развития РЖ, которое заканчивается примерно через 3 недели.
Медицинская значимость
[ редактировать ]Поскольку зародышевые центры являются важными структурами адаптивной иммунной системы , их нарушение регуляции подразумевается при многих иммунных заболеваниях, например, ревматоидном артрите , иммунодефиците и многих лимфомах, таких как DLBCL и лимфома Беркитта .
Зародышевые центры в эволюции
[ редактировать ]Несмотря на то, что рекомбинация V(D)J наблюдается у всех позвоночных появилась GC , у гомеотермных животных . В эволюционно новых условиях, когда повышенная температура тела способствовала увеличению скорости пролиферации микроорганизмов, диссеминации в тканях и их антигенной диверсификации. [9] Эти временные, но постоянно наблюдаемые гистологические структуры оказались полезными, поскольку их уникальное микроокружение могло обеспечить условия, благоприятные для перехода от первоначального широкого к последующему специфическому иммунному ответу, приводящему к дифференцировке клеток линии B в клетки, продуцирующие высокоаффинные антитела и поддерживающие длительную иммунную реакцию. стойкая гуморальная иммунная память. [10]
Среди хладнокровных позвоночных рыбы, по-видимому, имеют функционально аналогичные структуры, представленные «скоплениями Aicda». + клетки, окруженные пигментированными «мелано-макрофагами». [11]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Наткунам Ю. (1 января 2007 г.). «Биология зародышевого центра» . Гематология. Американское общество гематологии. Образовательная программа . 2007 : 210–215. doi : 10.1182/asheducation-2007.1.210 . ПМИД 18024632 .
- ^ Агуцци А., Кранич Дж., Краутлер, Нью-Джерси (март 2014 г.). «Фолликулярные дендритные клетки: происхождение, фенотип и функции в здоровье и болезни». Тенденции в иммунологии . 35 (3): 105–113. дои : 10.1016/j.it.2013.11.001 . ПМИД 24315719 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v Виктора, Габриэль Д.; Нусенцвейг, Мишель К. (26 апреля 2022 г.). «Герминальные центры» . Ежегодный обзор иммунологии . 40 (1): 413–442. doi : 10.1146/annurev-immunol-120419-022408 . ISSN 0732-0582 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Кеннеди, Доменик Э.; Кларк, Маркус Р. (31 марта 2021 г.). «Отделения и связи внутри зародышевого центра» . Границы в иммунологии . 12 . дои : 10.3389/fimmu.2021.659151 . ISSN 1664-3224 . ПМК 8045557 . ПМИД 33868306 .
- ^ Перейти обратно: а б с д и ж г час я дж к л Чхве, Сын Чул; Морель, Лоуренс (март 2020 г.). «Регуляция иммунного метаболизма реакции зародышевого центра» . Экспериментальная и молекулярная медицина . 52 (3): 348–355. дои : 10.1038/s12276-020-0392-2 . ISSN 2092-6413 . ПМК 7156389 . ПМИД 32132626 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д Лау, Анжелика, Вайоминг; Бринк, Роберт (1 апреля 2020 г.). «Отбор в зародышевом центре» . Современное мнение в иммунологии . Развитие и активация лимфоцитов. 63 : 29–34. дои : 10.1016/j.coi.2019.11.001 . ISSN 0952-7915 .
- ^ Торбеке Г.Дж., Амин А.Р., Циагбе В.К. (август 1994 г.). «Биология зародышевых центров лимфоидной ткани» . Журнал ФАСЭБ . 8 (11): 832–840. дои : 10.1096/fasebj.8.11.8070632 . ПМИД 8070632 . S2CID 83999556 .
- ^ Мейер-Германн М. (июнь 2002 г.). «Математическая модель морфологии зародышевого центра и созревания аффинности». Журнал теоретической биологии . 216 (3): 273–300. arXiv : физика/0203021 . Бибкод : 2002JThBi.216..273M . дои : 10.1006/jtbi.2002.2550 . ПМИД 12183119 . S2CID 2141888 .
- ^ Мансер, Тим (15 марта 2004 г.). «Зародышевые центры из учебника?» . Дж Иммунол . 172 (6): 3369–3375. дои : 10.4049/jimmunol.172.6.3369 .
- ^ Бринк, Р.; Фан, Т.Г. (2018). «Самореактивные B-клетки в реакции зародышевого центра». Ежегодный обзор иммунологии . 36 : 339–357. doi : 10.1146/annurev-immunol-051116-052510 .
- ^ Вали, Д.; Мутупандиан, А.; Фан, CW; Анцингер, Х.; Магор, Б.Г. (8 декабря 2022 г.). «Репертуар иммуноглобулинов VDJ обнаруживает признаки зародышевых центров в уникальных кластерах клеток, выделенных из лимфоидных тканей рыбок данио (Danio rerio)» . Границы в иммунологии . 13 : 1058877. дои : 10.3389/fimmu.2022.1058877 . ПМЦ 9772432 . PMID 36569890 .
{{cite journal}}: CS1 maint: дата и год ( ссылка )
Внешние ссылки
[ редактировать ]- Гистология UIUC, предмет 563
- Гистологическое изображение: 07103loa - Система обучения гистологии в Бостонском университете - «Лимфоидные ткани и органы: лимфатический узел, зародышевый центр»
- Гиперссылка на гистологию человека
- MedEd в Loyola Histo/practical/lymph/hp12-42.html