Ячейка памяти B
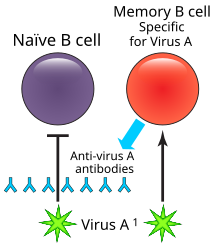
В иммунологии B- клетка памяти ( MBC ) представляет собой тип B-лимфоцита , который является частью адаптивной иммунной системы . Эти клетки развиваются в зародышевых центрах вторичных лимфоидных органов . В-клетки памяти циркулируют в кровотоке в состоянии покоя, иногда в течение десятилетий. [ 1 ] Их функция заключается в запоминании характеристик антигена , который активировал родительскую В-клетку во время первоначальной инфекции, так что, если В-клетка памяти позже встретит тот же антиген , она запускает ускоренный и устойчивый вторичный иммунный ответ . [ 2 ] [ 3 ] В-клетки памяти имеют рецепторы В-клеток (BCR) на клеточной мембране, идентичные рецепторам родительской клетки, которые позволяют им распознавать антиген и вызывать специфический ответ антител. [ 4 ]
Разработка и активация
[ редактировать ]Т-клеточно-зависимые механизмы
[ редактировать ]При Т-клеточно- зависимом пути развития наивные фолликулярные В-клетки активируются антигенпрезентирующими фолликулярными В-хелперными Т-клетками (TFH ) во время начальной инфекции или первичного иммунного ответа . [ 3 ] Наивные В-клетки циркулируют через фолликулы во вторичных лимфоидных органах (например, селезенке и лимфатических узлах ), где они могут быть активированы плавающим чужеродным пептидом, доставленным через лимфу , или антигеном, презентируемым антигенпрезентирующими клетками (АПК), такими как дендритные клетки (ДК) . ). [ 5 ] В-клетки также могут активироваться путем связывания чужеродного антигена на периферии, откуда они затем перемещаются во вторичные лимфоидные органы. [ 3 ] Сигнал, передаваемый при связывании пептида с В-клеткой, заставляет клетки мигрировать к краю фолликула, граничащему с областью Т-клеток. [ 5 ]
В-клетки усваивают чужеродные пептиды, расщепляют их и экспрессируют на главных комплексах гистосовместимости класса II (MHCII), которые представляют собой белки клеточной поверхности. Во вторичных лимфоидных органах большая часть В-клеток попадает в В-клеточные фолликулы, где формируется зародышевый центр. Большинство В-клеток в конечном итоге дифференцируются в плазматические клетки или В-клетки памяти внутри зародышевого центра. [ 3 ] [ 6 ] Т - ФГ , которые экспрессируют Т-клеточные рецепторы (TCR), родственные пептиду (т.е. специфичные для комплекса пептид-MHCII) на границе В-клеточного фолликула и Т-клеточной зоны, будут связываться с лигандом MHCII. Затем Т-клетки будут экспрессировать молекулу лиганда CD40 (CD40L) и начнут секретировать цитокины , которые заставляют В-клетки пролиферировать и подвергаться рекомбинации с переключением классов — мутации в генетическом кодировании В-клеток, которая меняет тип их иммуноглобулина . [ 7 ] [ 8 ] Переключение классов позволяет В-клеткам памяти секретировать различные типы антител в будущих иммунных ответах. [ 3 ] Затем В-клетки дифференцируются либо в плазматические клетки , В-клетки зародышевого центра или В-клетки памяти, в зависимости от экспрессируемых факторов транскрипции . Активированные В-клетки, экспрессирующие фактор транскрипции Bcl-6, проникают в В-клеточные фолликулы и подвергаются реакциям зародышевого центра. [ 7 ]
Попав внутрь зародышевого центра, В-клетки подвергаются пролиферации, за которой следует мутация генетической кодирующей области их BCR — процесс, известный как соматическая гипермутация . [ 3 ] Мутации либо увеличивают, либо уменьшают сродство поверхностного рецептора к определенному антигену, этот прогресс называется созреванием сродства . После приобретения этих мутаций рецепторы на поверхности В-клеток (рецепторы В-клеток) проверяются внутри зародышевого центра на их сродство к текущему антигену. [ 9 ] Клоны В-клеток с мутациями, которые увеличили аффинность их поверхностных рецепторов, получают сигналы выживания посредством взаимодействия с родственными им клетками T FH . [ 2 ] [ 3 ] [ 10 ] В-клетки, которые не обладают достаточно высоким сродством для получения этих сигналов выживания, а также В-клетки, которые потенциально являются аутореактивными, будут отобраны и погибнут в результате апоптоза. [ 6 ] Эти процессы увеличивают вариабельность сайтов связывания антигена, так что каждая вновь созданная В-клетка имеет уникальный рецептор. [ 11 ]
После дифференцировки В-клетки памяти перемещаются на периферию тела, где они с большей вероятностью столкнутся с антигеном в случае будущего воздействия. [ 6 ] [ 2 ] [ 3 ] Многие из циркулирующих В-клеток концентрируются в областях тела, которые имеют высокую вероятность контакта с антигеном, например, в пейеровых пятнах .
Процесс дифференцировки в В-клетки памяти внутри зародышевого центра еще полностью не изучен. [ 3 ] Некоторые исследователи предполагают, что дифференцировка в В-клетки памяти происходит случайным образом. [ 6 ] [ 4 ] Другие гипотезы предполагают, что транскрипционный фактор NF-κB и цитокин IL-24 участвуют в процессе дифференцировки в В-клетки памяти. [ 11 ] [ 3 ] Дополнительная гипотеза гласит, что В-клетки с относительно более низким сродством к антигену станут В-клетками памяти, в отличие от В-клеток с относительно более высоким сродством, которые станут плазматическими клетками.
Независимые от Т-клеток механизмы
[ редактировать ]Не все В-клетки, присутствующие в организме, подверглись соматическим гипермутациям. IgM+ В-клетки памяти, которые не подверглись рекомбинации с переключением классов, демонстрируют, что В-клетки памяти могут продуцироваться независимо от зародышевых центров.
Первичный ответ
[ редактировать ]При заражении патогеном многие В-клетки дифференцируются в плазматические клетки , также называемые эффекторными В-клетками, которые производят первую волну защитных антител и помогают избавиться от инфекции. [ 6 ] [ 2 ] Плазматические клетки секретируют антитела, специфичные для патогенов, но они не могут реагировать на вторичное воздействие. Часть В-клеток с BCR, родственными антигену, дифференцируется в В-клетки памяти, которые долговременно выживают в организме. [ 12 ] В-клетки памяти могут поддерживать экспрессию BCR и смогут быстро реагировать на вторичное воздействие. [ 6 ]
Вторичная реакция и память
[ редактировать ]В-клетки памяти, образующиеся во время первичного иммунного ответа, специфичны к антигену, задействованному во время первого воздействия. При вторичном ответе отреагируют В-клетки памяти, специфичные к антигену или подобным антигенам. [ 3 ] Когда В-клетки памяти повторно встречают свой специфический антиген, они пролиферируют и дифференцируются в плазматические клетки, которые затем реагируют на антиген и удаляют его. [ 3 ] В-клетки памяти, которые на этом этапе не дифференцируются в плазматические клетки, могут повторно войти в зародышевые центры, чтобы подвергнуться дальнейшему переключению классов или соматической гипермутации для дальнейшего созревания аффинности. [ 3 ] Дифференциация В-клеток памяти в плазматические клетки происходит намного быстрее, чем дифференцировка наивными В-клетками, что позволяет В-клеткам памяти производить более эффективный вторичный иммунный ответ. [ 4 ] Эффективность и накопление ответа В-клеток памяти является основой вакцин и повторных прививок. [ 4 ] [ 3 ]
Фенотип клеток памяти, который предсказывает судьбу плазматических клеток или клеток зародышевого центра, был открыт несколько лет назад. На основании сравнения экспрессионных микрочипов В-клеток памяти и наивных В-клеток было выявлено, что существует несколько поверхностных белков, таких как CD80 , PD-L2 и CD73, которые экспрессируются только на В-клетках памяти, поэтому они также служат для деления этих клеток. клетки в нескольких фенотипических подмножествах. [ 13 ] Более того, было показано, что клетки памяти, экспрессирующие CD80, PD-L2 и CD73, с большей вероятностью станут плазматическими клетками. С другой стороны, клетки, не имеющие маркеров такого типа, с большей вероятностью образуют клетки зародышевого центра. IgM + В-клетки памяти не экспрессируют CD80 или CD73, тогда как IgG + выразить их. Более того, IgG + с большей вероятностью дифференцируются в клетки, секретирующие антитела. [ 14 ]
Продолжительность жизни
[ редактировать ]В-клетки памяти могут выживать десятилетиями, что дает им способность реагировать на многократное воздействие одного и того же антигена. [ 3 ] Предполагается, что длительное выживание является результатом определенных генов антиапоптоза, которые более высоко экспрессируются в В-клетках памяти, чем в других подмножествах В-клеток. [ 6 ] Кроме того, В-клетке памяти не требуется постоянного взаимодействия ни с антигеном, ни с Т-клетками, чтобы выжить в долгосрочной перспективе. [ 4 ]
Однако верно то, что продолжительность жизни отдельных В-клеток памяти остается плохо определенной, хотя они играют решающую роль в долговременном иммунитете. В одном исследовании с использованием трансгенной системы B-клеточного рецептора (BCR) (это была модель трансгенной мышиной H-цепи, у которой отсутствовал секретируемый Ig, поэтому она не откладывала Ag-содержащие иммунные комплексы), было показано, что количество B-клеток памяти остаются постоянными в течение примерно 8–20 недель после иммунизации. Было также подсчитано, что период полураспада В-клеток памяти составлял от 8 до 10 недель после проведения эксперимента, в котором клетки обрабатывались in vivo бромдезоксиуридином . [ 15 ] В других экспериментах на мышах было показано, что продолжительность жизни В-клеток памяти как минимум в 9 раз превышает продолжительность жизни фолликулярных наивных В-клеток. [ 16 ]
Маркеры
[ редактировать ]В-клетки памяти обычно отличаются маркером клеточной поверхности CD27, хотя некоторые подгруппы не экспрессируют CD27. В-клетки памяти, в которых отсутствует CD27, обычно связаны с истощением В-клеток или некоторыми аутоиммунными состояниями, такими как ВИЧ, волчанка или ревматоидный артрит. [ 2 ] [ 3 ]
Поскольку В-клетки обычно подвергаются переключению класса, они могут экспрессировать ряд молекул иммуноглобулина . Некоторые специфические свойства конкретных молекул иммуноглобулина описаны ниже:
- IgM: В-клетки памяти, экспрессирующие IgM, сконцентрированы в миндалинах , пейеровых бляшках и лимфатических узлах. [ 3 ] Эта подгруппа В-клеток памяти с большей вероятностью будет пролиферировать и повторно проникать в зародышевый центр во время вторичного иммунного ответа. [ 4 ]
- IgG: В-клетки памяти, которые экспрессируют IgG, обычно дифференцируются в плазматические клетки. [ 4 ]
- IgE: В-клетки памяти, экспрессирующие IgE, у здоровых людей встречаются очень редко. Это может произойти потому, что В-клетки, экспрессирующие IgE, чаще дифференцируются в плазматические клетки, а не в В-клетки памяти. [ 4 ]
- Только IgD: В-клетки памяти, экспрессирующие IgD, встречаются очень редко. В-клетки, содержащие только IgD, обнаруживаются в миндалинах. [ 3 ]
Важно отметить важность интеграции сигнальных путей, связанных с рецепторами BCR и TLR, для модуляции продукции антител за счет расширения В-клеток памяти. Следовательно, существуют разные факторы, которые предоставляют информацию для секреции разных типов антител. Было продемонстрировано, что продукция специфических IgG1, анафилактических IgG1 и общего IgE зависит от сигнала, вырабатываемого TLR2 и Myd88 . Более того, сигнал, вырабатываемый TLR4 при его стимуляции наттеринами (белком, полученным из яда рыб T. nattereri ), ускоряет синтез антитела IgE, действующего как адъювант, как было показано в эксперименте in vivo на мышах. [ 17 ]
Рецептор CCR6 обычно является маркером В-клеток, которые в конечном итоге дифференцируются в MBC. Этот рецептор обнаруживает хемокины , которые являются химическими посланниками, которые позволяют В-клетке перемещаться внутри тела. В-клетки памяти могут иметь этот рецептор, позволяющий им выходить из зародышевого центра в ткани, где у них более высокая вероятность встречи с антигеном. [ 6 ]
Было показано, что В-клетки памяти имеют высокий уровень экспрессии CCR6, а также повышенный хемотаксический ответ на лиганд CCR6 ( CCL20 ) по сравнению с наивными В-клетками. Тем не менее, у мышей с дефицитом CCR6 первичный гуморальный ответ и поддержание В-клеток памяти не затрагиваются. Однако эффективного вторичного ответа со стороны В-клеток памяти не происходит при повторном воздействии антигена, если клетки не экспрессируют CCR6. Таким образом, мы можем подтвердить, что CCR6 необходим для способности В-клеток памяти вызывать родственный антиген, а также для соответствующего анатомического расположения этих клеток. [ 18 ]
Подмножества
[ редактировать ]В-клетки памяти, независимые от зародышевого центра
[ редактировать ]Эта подгруппа клеток дифференцируется из активированных В-клеток в В-клетки памяти перед входом в зародышевый центр. В-клетки, которые имеют высокий уровень взаимодействия с Т -ФГ внутри В-клеточного фолликула, имеют более высокую склонность к проникновению в зародышевый центр. В-клетки, которые развиваются в В-клетки памяти независимо от зародышевых центров, вероятно, испытывают передачу сигналов CD40 и цитокинов от Т-клеток. [ 4 ] Переключение классов все еще может происходить до взаимодействия с зародышевым центром, тогда как соматическая гипермутация происходит только после взаимодействия с зародышевым центром. [ 4 ] Предполагается, что отсутствие соматических гипермутаций полезно; более низкий уровень созревания аффинности означает, что эти В-клетки памяти менее специализированы к конкретному антигену и могут распознавать более широкий спектр антигенов. [ 11 ] [ 19 ] [ 4 ]
Т-независимые В-клетки памяти
Т-независимые В-клетки памяти представляют собой подмножество, называемое клетками В1. Эти клетки обычно находятся в брюшной полости. При повторном введении в антиген некоторые из этих В1-клеток могут дифференцироваться в В-клетки памяти, не взаимодействуя с Т-клетками. [ 4 ] Эти В-клетки производят антитела IgM, помогающие избавиться от инфекции. [ 20 ]
В-клетки памяти T-bet
В-клетки T-bet представляют собой подгруппу, которая, как было обнаружено, экспрессирует фактор транскрипции T-bet. T-ставка связана с переключением класса. Считается, что T-bet B-клетки играют важную роль в иммунном ответе против внутриклеточных бактериальных и вирусных инфекций. [ 21 ]
Вакцинация
[ редактировать ]Вакцины основаны на понятии иммунологической памяти . Превентивное введение в организм непатогенного антигена позволяет организму создать стойкую иммунологическую память . Инъекция антигена приводит к выработке антител с последующей выработкой В-клеток памяти. Эти В-клетки памяти быстро реактивируются при заражении антигеном и могут эффективно защитить организм от болезней. [ 22 ]
Долгоживущие плазматические клетки и В-клетки памяти отвечают за долговременный гуморальный иммунитет, вызываемый большинством вакцин. Был проведен эксперимент с целью наблюдения за долговечностью В-клеток памяти после вакцинации, в данном случае с противооспенной вакциной (DryVax), которая была выбрана в связи с тем, что оспа была ликвидирована, поэтому иммунная память к оспе является полезным. Эталон, позволяющий понять долговечность В-клеток иммунной памяти в отсутствие рестимуляции. Исследование пришло к выводу, что специфические В-клетки памяти сохраняются в течение десятилетий, что указывает на то, что иммунологическая память долговременно сохраняется в компартменте В-клеток после сильного первоначального воздействия антигена. [ 23 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Кротти С., Фельгнер П., Дэвис Х., Глайдвелл Дж., Вильярреал Л., Ахмед Р. (ноябрь 2003 г.). «Авангард: долговременная память В-клеток у людей после вакцинации против оспы» . Журнал иммунологии . 171 (10): 4969–4973. дои : 10.4049/jimmunol.171.10.4969 . ПМИД 14607890 .
- ^ Jump up to: а б с д и Вайзель Ф., Шломчик М. (апрель 2017 г.). «В-клетки памяти мышей и людей» . Ежегодный обзор иммунологии . 35 (1): 255–284. doi : 10.1146/annurev-immunol-041015-055531 . ПМИД 28142324 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р Зайферт М., Кюпперс Р. (декабрь 2016 г.). «В-клетки памяти человека». Лейкемия . 30 (12): 2283–2292. дои : 10.1038/leu.2016.226 . ПМИД 27499139 . S2CID 28936531 .
- ^ Jump up to: а б с д и ж г час я дж к л Куросаки Т., Кометани К., Исэ В. (март 2015 г.). «В-клетки памяти». Обзоры природы. Иммунология . 15 (3): 149–159. дои : 10.1038/nri3802 . ПМИД 25677494 . S2CID 20825732 .
- ^ Jump up to: а б Гарсайд П., Ингулли Э., Мерика Р.Р., Джонсон Дж.Г., Ноэль Р.Дж., Дженкинс М.К. (июль 1998 г.). «Визуализация специфических взаимодействий В- и Т-лимфоцитов в лимфатическом узле». Наука . 281 (5373): 96–99. Бибкод : 1998Sci...281...96G . дои : 10.1126/science.281.5373.96 . ПМИД 9651253 .
- ^ Jump up to: а б с д и ж г час Суан Д., Сандлинг К., Бринк Р. (апрель 2017 г.). «Дифференцировка плазматических клеток и В-клеток памяти из зародышевого центра». Современное мнение в иммунологии . Развитие и активация лимфоцитов * Иммунология опухолей. 45 : 97–102. дои : 10.1016/j.coi.2017.03.006 . ПМИД 28319733 .
- ^ Jump up to: а б Тейлор Дж. Дж., Дженкинс М.К., Папе К.А. (декабрь 2012 г.). «Гетерогенность в дифференцировке и функции В-клеток памяти» . Тенденции в иммунологии . 33 (12): 590–597. дои : 10.1016/j.it.2012.07.005 . ПМЦ 3505266 . ПМИД 22920843 .
- ^ Фан Т.Г., Танье С.Г. (апрель 2017 г.). «В-клетки памяти: полное воспоминание». Современное мнение в иммунологии . Развитие и активация лимфоцитов * Иммунология опухолей. 45 : 132–140. дои : 10.1016/j.coi.2017.03.005 . ПМИД 28363157 .
- ^ Оллман Д., Уилмор-младший, Годетт Б.Т. (март 2019 г.). «Продолжение истории антител, независимых от Т-клеток» . Иммунологические обзоры . 288 (1): 128–135. дои : 10.1111/imr.12754 . ПМК 6653682 . ПМИД 30874357 .
- ^ Виктора Г.Д., Nussenzweig MC (26 марта 2012 г.). «Герминальные центры». Ежегодный обзор иммунологии . 30 (1): 429–457. doi : 10.1146/annurev-immunol-020711-075032 . ПМИД 22224772 . S2CID 20168324 .
- ^ Jump up to: а б с Шиннакасу Р., Куросаки Т. (апрель 2017 г.). «Регуляция памяти B и дифференцировка плазматических клеток». Современное мнение в иммунологии . Развитие и активация лимфоцитов * Иммунология опухолей. 45 : 126–131. дои : 10.1016/j.coi.2017.03.003 . ПМИД 28359033 .
- ^ Гатто Д., Бринк Р. (ноябрь 2010 г.). «Реакция зародышевого центра» . Журнал аллергии и клинической иммунологии . 126 (5): 898–907, викторина 908–9. дои : 10.1016/j.jaci.2010.09.007 . ПМИД 21050940 .
- ^ Цуккарино-Катания, Гризельда V; Садананд, Сахели; Вайзель, Флориан Дж; Томайко, Мэри М; Мэн, Хайлун; Кляйнштейн, Стивен Х; Гуд-Джейкобсон, Ким Л; Шломчик, Марк Дж (июль 2014 г.). «CD80 и PD-L2 определяют функционально различные подмножества B-клеток памяти, которые не зависят от изотипа антитела» . Природная иммунология . 15 (7): 631–637. дои : 10.1038/ni.2914 . ISSN 1529-2908 . ПМК 4105703 . ПМИД 24880458 .
- ^ Чонг, Анита С.; Ансари, М. Джавид (апрель 2018 г.). «Гетерогенность В-клеток памяти» . Американский журнал трансплантологии . 18 (4): 779–784. дои : 10.1111/ajt.14669 . ПМЦ 5962275 . ПМИД 29359404 .
- ^ Андерсон С.М., Ханнум Л.Г., Шломчик М.Ю. (апрель 2006 г.). «Выживание и функционирование B-клеток памяти в отсутствие секретируемых антител и иммунных комплексов на фолликулярных дендритных клетках» . Журнал иммунологии . 176 (8): 4515–4519. дои : 10.4049/jimmunol.176.8.4515 . ПМИД 16585539 . S2CID 85927231 .
- ^ Джонс Д.Д., Уилмор-младший, Оллман Д. (ноябрь 2015 г.). «Клеточная динамика популяций B-клеток памяти: IgM + и IgG + B-клетки памяти сохраняются неопределенно долго в состоянии покоя» . Журнал иммунологии . 195 (10): 4753–4759. doi : 10.4049/jimmunol.1501365 . ПМЦ 4637268 . ПМИД 26438523 .
- ^ Комегаэ, Эвилин Нанаме; Грунд, Лидиана Зито; Лопес-Феррейра, Моника; Лима, Карла (5 августа 2013 г.). Ричард, Иоланда (ред.). «TLR2, TLR4 и передача сигналов MyD88 имеют решающее значение для генерации in vivo и долговечности долгоживущих клеток, секретирующих антитела» . ПЛОС ОДИН . 8 (8): е71185. Бибкод : 2013PLoSO...871185K . дои : 10.1371/journal.pone.0071185 . ISSN 1932-6203 . ПМЦ 3733974 . ПМИД 23940714 .
- ^ Эльгета, Рауль; Маркс, Эллен; Новак, Элизабет; Менезес, Шинель; Бенсон, Мика; Раман, Ванита С.; Ортис, Карла; О'Коннелл, Сэмюэл; Гесс, Генри; Лорд, Грэм М.; Ноэль, Рэндольф (15 января 2015 г.). «CCR6-зависимое расположение B-клеток памяти необходимо для их способности вызывать ответную реакцию на антиген» . Журнал иммунологии . 194 (2): 505–513. doi : 10.4049/jimmunol.1401553 . ISSN 0022-1767 . ПМЦ 4282958 . ПМИД 25505290 .
- ^ Пуповац А., Гуд-Джейкобсон К.Л. (апрель 2017 г.). «Антиген, который следует помнить: регуляция памяти В-клеток в здоровом состоянии и при заболеваниях» . Современное мнение в иммунологии . Развитие и активация лимфоцитов * Иммунология опухолей. 45 : 89–96. дои : 10.1016/j.coi.2017.03.004 . ПМК 7126224 . ПМИД 28319732 .
- ^ Монтесино-Родригес Э., Доршкинд К. (январь 2012 г.). «Развитие B-1 B-клеток у плода и взрослого» . Иммунитет . 36 (1): 13–21. doi : 10.1016/j.immuni.2011.11.017 . ПМК 3269035 . ПМИД 22284417 .
- ^ Нокс Джей-Джей, Майлз А., член парламента от Канкро (март 2019 г.). "Т-ставка + B-клетки памяти: генерация, функции и судьба» . Immunological Reviews . 288 (1): 149–160. : 10.1111 /imr.12736 . PMC 6626622. . PMID 30874358 doi
- ^ Дхенни Р., Фан Т.Г. (июль 2020 г.). «География реактивации В-клеток памяти при вакцино-индуцированном иммунитете и при рецидивах аутоиммунных заболеваний». Иммунологические обзоры . 296 (1): 62–86. дои : 10.1111/imr.12862 . ПМИД 32472583 . S2CID 219103939 .
- ^ Кротти, Шейн; Фельгнер, Фил; Дэвис, Хью; Глайдвелл, Джон; Вильярреал, Луис; Ахмед, Рафи (15 ноября 2003 г.). «Авангард: долговременная память B-клеток у людей после вакцинации от оспы» . Журнал иммунологии . 171 (10): 4969–4973. дои : 10.4049/jimmunol.171.10.4969 . ISSN 0022-1767 . ПМИД 14607890 . S2CID 32215286 .