Соединительное разнообразие
Разнообразие соединений описывает изменения последовательности ДНК , вызванные неправильным соединением сегментов гена в процессе рекомбинации V(D)J . Этот процесс рекомбинации V(D)J играет жизненно важную роль для позвоночных иммунной системы , поскольку он способен генерировать огромный набор различных рецепторов Т-клеток (TCR) и молекул иммуноглобулинов, необходимых для патогена распознавания антигена Т-клетками и B-клетками. клетки соответственно.
Процесс
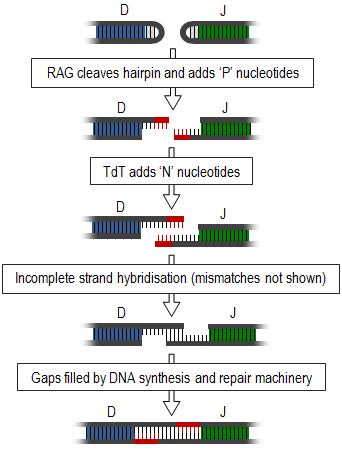
[ редактировать ]Соединительное разнообразие включает процесс соматической рекомбинации или V(D)J-рекомбинации , во время которого различные вариабельные генные сегменты (сегменты, участвующие в распознавании антигена) TCR и иммуноглобулинов перестраиваются, а неиспользуемые сегменты удаляются. Это приводит к появлению двухцепочечных разрывов между необходимыми сегментами. Эти концы образуют петли-шпильки, и их необходимо соединить вместе, чтобы сформировать одну прядь (показано на схеме справа). Это соединение представляет собой очень неточный процесс, который приводит к переменному добавлению или вычитанию нуклеотидов и, таким образом, создает разнообразие соединений. [1]
Генерация разнообразия соединений начинается с белков, активирующих рекомбинацию генов -1 и -2 (RAG1 и RAG2), а также белков репарации ДНК, таких как Artemis , [2] ответственны за одноцепочечное расщепление шпилек и добавление ряда палиндромных нуклеотидов «P». После этого фермент терминальная дезоксинуклеотидилтрансфераза (TdT) добавляет дополнительные случайные N-нуклеотиды. Вновь синтезированные цепи отжигаются друг с другом, но несовпадения являются обычным явлением. Экзонуклеазы удаляют эти неспаренные нуклеотиды, а пробелы заполняются ДНК механизмами синтеза и восстановления . [1] [3] Экзонуклеазы также могут вызывать укорочение этого соединения, однако этот процесс еще плохо изучен. [4]
Разнообразие соединений может вызывать мутации сдвига рамки и, следовательно, образование нефункциональных белков. Поэтому,в этом процессе участвуют значительные отходы. [1]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Джейнвей, Калифорния, Трэверс, П., Уолпорт, М., Шломчик, М.Дж. (2005). Иммунология (6-е изд.). Гирляндная наука.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ма, Ю., Паннике, У., Шварц, К., Либер, М.Р. (2004). «Открытие шпильки и обработка выступа с помощью комплекса Artemis/ДНК-зависимой протеинкиназы при негомологичном соединении концов и рекомбинации V (D) J» . Клетка . 108 (6): 781–794. дои : 10.1016/S0092-8674(02)00671-2 . ПМИД 11955432 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Вайман К., Канаар Р. (2006). «Восстановление двухцепочечного разрыва ДНК: все хорошо, что хорошо кончается». Ежегодный обзор генетики . 40 : 363–383. дои : 10.1146/annurev.genet.40.110405.090451 . ПМИД 16895466 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Крагель, М.С. (2009). «Механика перестройки генов рецепторов Т-клеток» . Современное мнение в иммунологии . 21 (2): 133–139. дои : 10.1016/j.coi.2009.03.009 . ПМК 2676214 . ПМИД 19362456 .