ILC2
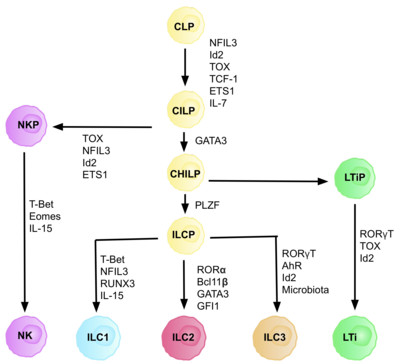
Клетки ILC2 , или врожденные лимфоидные клетки 2 типа, представляют собой тип врожденных лимфоидных клеток . Не путать с КМП. Они происходят от общего лимфоидного предшественника и принадлежат к лимфоидной линии . Эти клетки лишены антигенспецифических В- или Т-клеточных рецепторов из-за отсутствия гена, активирующего рекомбинацию . [2] ILC2 продуцируют цитокины 2 типа (например, IL-4 , IL-5 , IL-9 , IL-13 ) и участвуют в реакциях на гельминтов , аллергены , [3] некоторые вирусы, такие как вирус гриппа [4] и рак. [5] [6] [7]
Этот тип клеток был впервые описан в 2001 году как не-B/не-T-клетки, которые продуцируют IL-5 и IL-13 в ответ на IL-25 и экспрессируют MHC класса II и CD11c . [8] В 2006 г. аналогичная популяция клеток была выявлена при глистной инфекции. [9] Название «ILC2» не предлагалось до 2013 года. [10] Ранее в литературе они были идентифицированы как естественные клетки-хелперы. [11] нуоциты , [12] или врожденный хелпер 2 клетки. [13] Считается, что ILC2 представляют собой довольно старый тип клеток, предковые популяции которого появились у миноги и костистых рыб . [14]
Паразитарная инфекция
[ редактировать ]ILC2 играют решающую роль в секреции цитокинов 2 типа в ответ на крупных внеклеточных паразитов. Они экспрессируют характерные поверхностные маркеры и рецепторы хемокинов , которые участвуют в распределении лимфоидных клеток по определенным участкам органов. Для их развития им необходим IL-7 , который активирует два фактора транскрипции (оба необходимы этим клеткам) — RORα и GATA3 . После стимуляции Th2- поляризующими цитокинами, которые секретируются в основном эпителием (например, IL-25 , IL-33 , TSLP , простагландином D2 и лейкотриеном D4 ), ILC2 начинают продуцировать IL-5 , IL-13 , IL-9 , IL- 4 быстро. ILC2 имеют решающее значение для первичного ответа на местные антигены Th2, например, на гельминты и вирусы, и именно поэтому ILC2 в изобилии присутствуют в тканях кожи. [15] [16] легкие, печень и кишечник. [17] Было замечено, что ILC2 возникают в кишечнике, попадают в лимфатические сосуды, а затем циркулируют в кровотоке, откуда могут мигрировать в другие органы, помогая бороться с паразитарной инфекцией. Незаконный оборот частично зависит от сфингозин-1-фосфата . [16] Например, во время инфекции Nippostrongylus brasiliensis ILC2 способствуют выведению гельминтов путем выработки незаменимого цитокина IL-13 . [18] IL-13, секретируемый ILC2, также способствует миграции активированных дендритных клеток легких в дренирующий лимфатический узел , что затем приводит к наивному праймированию Т-клеток и дифференцировке в клетки Th2 . [19]
Респираторная вирусная инфекция
[ редактировать ]Было замечено, что ILC2 активируются при инфекциях респираторных вирусов у мышей и людей. Например, во время инфекции вирусом гриппа А , который индуцирует выработку IL-33 , ILC2 активируются и вызывают гиперреактивность дыхательных путей. Другим примером является инфекция респираторно-синцитиального вируса , где ILC2 вносят свой вклад, будучи основным источником IL-13 на ранних стадиях инфекции, что приводит к гиперреактивности дыхательных путей и увеличению выработки слизи. [18]
Аллергия, атопический дерматит и астма
[ редактировать ]ILC2 играют разнообразную роль при аллергии . [19] Прежде всего, они являются источником цитокинов 2-го типа, которые управляют аллергическим иммунным ответом. Они производят профиль сигналов в ответ на проаллергенные цитокины IL-25 и IL-33 , аналогичный тем, которые вырабатываются в ответ на глистную инфекцию. Их вклад в эту передачу сигналов, по-видимому, сравним с вкладом Т-клеток. В ответ на воздействие аллергена в легких ILC2 вырабатывают IL-13, необходимый цитокин в патогенезе аллергических реакций. Этот ответ, по-видимому, не зависит от Т- и В-клеток. Кроме того, у мышей, у которых отсутствуют Т- и В-клетки, с помощью IL-33 были вызваны аллергические реакции, напоминающие астматические симптомы. Также было обнаружено, что ILC2 присутствуют в более высоких концентрациях в тканях, где присутствуют аллергические симптомы, например, в носовых полипах пациентов с хроническим риносинуситом и коже пациентов с атопическим дерматитом . [20] [21] [22]
Барьерная функция
[ редактировать ]Известно, что ILC2 обогащены жировыми лимфоидными кластерами (FALC) внутри брыжейки . IL-5, секретируемый ILC2, является важным фактором роста для B1 B-клеток и, следовательно, важен для производства антител IgA . Помимо цитокинов 2 типа, ILC2 также могут продуцировать IL-6 , который индуцирует выработку антител В-клетками, действует как фактор роста плазмобластов и способствует регуляции Т-фолликулярных хелперных клеток . [23] [24]
Также известно, что ILC2 присутствуют в FALC в плевральной полости . После стимуляции IL-33 во время инфекции они начинают секретировать IL-5, что приводит к активации B1 B-клеток и выработке антител IgM . [25] ILC2 являются доминирующей популяцией ILC в легких. Продуцируя IL-13 , они могут инициировать сокращение гладких мышц и секрецию слизи , а также гиперплазию бокаловидных клеток , если IL-13 сверхэкспрессируется. Кроме того, ILC2 помогают заживлению ран легких после инфекции гриппа, секретируя амфирегулин . Помимо легких, популяции ILC2 также можно обнаружить в тканях носа и миндалин человека . [18]
Гомеостаз жировой ткани
[ редактировать ]ILC2 необходимы для поддержания гомеостаза в сухой и здоровой жировой ткани . ILC2, резидентные висцеральной жировой ткани, продуцируют IL-5, IL-13 и пептиды метионин-энкефалин после длительного воздействия IL-33 . IL-5, секретируемый ILC2 в жировой ткани, имеет решающее значение для рекрутирования и поддержания эозинофилов . Кроме того, продукция IL-13 и IL-4 ILC2 и эозинофилами поддерживает поддержание альтернативно активированных макрофагов M2 и гомеостаз глюкозы . [24]
Исследования выявили нарушение регуляции ответов ILC2 в жировой ткани как фактор развития ожирения у мышей, поскольку ILC2 также играют важную роль в энергетическом гомеостазе . Пептиды метионин-энкефалина, продуцируемые ILC2, действуют непосредственно на адипоциты , активируя UCP1 и способствуя появлению бежевых адипоцитов в белой жировой ткани . [26] Бежевая и бурая жировая ткань специализируются на термогенезе . [24] Процесс старения приводит к увеличению затрат энергии и уменьшению ожирения. [26]
Ссылки
[ редактировать ]- ^ Вивье Э., Артис Д., Колонна М., Дифенбах А., Ди Санто Дж. П., Эберл Г. и др. (август 2018 г.). «Врожденные лимфоидные клетки: 10 лет спустя» . Клетка . 174 (5): 1054–1066. дои : 10.1016/j.cell.2018.07.017 . ПМИД 30142344 .
- ^ Спитс Х, Купедо Т (2012). «Врожденные лимфоидные клетки: новые знания о развитии, родственных отношениях и функциях». Ежегодный обзор иммунологии . 30 : 647–75. doi : 10.1146/annurev-immunol-020711-075053 . ПМИД 22224763 .
- ^ Доэрти Т.А., Бройд Д.Х. (январь 2019 г.). «Врожденные лимфоидные клетки дыхательных путей в индукции и регуляции аллергии» . Международная аллергология . 68 (1): 9–16. дои : 10.1016/j.alit.2018.11.001 . ПМК 6614863 . ПМИД 30473412 .
- ^ Монтичелли Л.А., Зонненберг Г.Ф., Абт М.К., Аленгат Т., Зиглер К.Г., Деринг Т.А. и др. (ноябрь 2011 г.). «Врожденные лимфоидные клетки способствуют гомеостазу легочной ткани после заражения вирусом гриппа» . Природная иммунология . 12 (11): 1045–54. дои : 10.1038/ni.2131 . ПМК 3320042 . ПМИД 21946417 .
- ^ Вагнер, Марек; Или, Кофе Н.; Тецу, Хироэ; Кинива, Цуёси; Мотомура, Ясутака; Моро, Кадзуё; Коясу, Сигео (февраль 2020 г.). «Молочная кислота, полученная из опухоли, способствует недостаточности внутриопухолевых ILC2» . о ячейках Отчеты 30 (8): 2743–2757.e5. дои : 10.1016/j.cellrep.2020.01.103 . HDL : 11250/2763785 . ПМИД 32101749 .
- ^ Вагнер, Марек; Коясу, Сигео (май 2019 г.). «Иммуноредактирование рака с помощью врожденных лимфоидных клеток» . Тенденции в иммунологии . 40 (5): 415–430. дои : 10.1016/j.it.2019.03.004 . ПМИД 30992189 . S2CID 119093972 .
- ^ Вагнер, Марек; Моро, Кадзуё; Коясу, Сигео (май 2017 г.). «Пластическая гетерогенность врожденных лимфоидных клеток при раке» . Тенденции рака . 3 (5): 326–335. дои : 10.1016/j.trecan.2017.03.008 . ПМИД 28718410 .
- ^ Форт ММ, Чунг Дж., Йен Д., Ли Дж., Журавски С.М., Ло С. и др. (декабрь 2001 г.). «IL-25 индуцирует IL-4, IL-5, IL-13 и Th2-ассоциированные патологии in vivo» . Иммунитет . 15 (6): 985–95. дои : 10.1016/S1074-7613(01)00243-6 . ПМИД 11754819 .
- ^ Фэллон П.Г., Баллантайн С.Дж., Манган Н.Е., Барлоу Дж.Л., Дасварма А., Хьюитт Д.Р. и др. (апрель 2006 г.). «Идентификация зависимой от интерлейкина (IL)-25 клеточной популяции, которая обеспечивает IL-4, IL-5 и IL-13 в начале изгнания гельминтов» . Журнал экспериментальной медицины . 203 (4): 1105–16. дои : 10.1084/jem.20051615 . ПМК 2118283 . ПМИД 16606668 .
- ^ Лунд С., Уолфорд Х.Х., Доэрти Т.А. (ноябрь 2013 г.). «Врожденные лимфоидные клетки 2-го типа при аллергических заболеваниях» . Текущие обзоры иммунологии . 9 (4): 214–221. дои : 10.2174/1573395510666140304235916 . ПМК 4033554 . ПМИД 24876829 .
- ^ Моро К., Ямада Т., Танабэ М., Такеучи Т., Икава Т., Кавамото Х. и др. (январь 2010 г.). «Врожденная продукция цитокинов T(H)2 лимфоидными клетками c-Kit(+)Sca-1(+) жировой ткани». Природа . 463 (7280): 540–4. дои : 10.1038/nature08636 . ПМИД 20023630 . S2CID 4420895 .
- ^ Нил Д.Р., Вонг С.Х., Беллоси А., Флинн Р.Дж., Дейли М., Лэнгфорд Т.К. и др. (апрель 2010 г.). «Нуоциты представляют собой новый врожденный эффекторный лейкоцит, который опосредует иммунитет 2 типа» . Природа . 464 (7293): 1367–70. Бибкод : 2010Natur.464.1367N . дои : 10.1038/nature08900 . ПМЦ 2862165 . ПМИД 20200518 .
- ^ Прайс А.Е., Лян Х.Э., Салливан Б.М., Рейнхардт Р.Л., Эйсли С.Дж., Эрл Дж.Д. и др. (июнь 2010 г.). «Системно рассеянные врожденные клетки, экспрессирующие IL-13, при иммунитете 2 типа» . Proc Natl Acad Sci . 107 (25): 11489–94. дои : 10.1073/pnas.1003988107 . ПМК 2895098 . ПМИД 20534524 .
- ^ Вивье, Эрик; ван де Павер, Серж А; Купер, Макс Д.; Белз, Габриэль Т (21 июня 2016 г.). «Эволюция врожденных лимфоидных клеток» . Природная иммунология . 17 (7): 790–794. дои : 10.1038/ni.3459 . ISSN 1529-2908 . ПМК 5287353 . ПМИД 27328009 .
- ^ Рёдигер Б., Кайл Р., Йип К.Х., Сумария Н., Гай Т.В., Ким Б.С. и др. (июнь 2013 г.). «Кожный иммунонадзор и регуляция воспаления врожденными лимфоидными клетками 2 группы» . Природная иммунология . 14 (6): 564–73. дои : 10.1038/ni.2584 . ПМЦ 4282745 . ПМИД 23603794 .
- ^ Перейти обратно: а б Фань Х., Ван А., Ван Ю., Сунь Ю., Хань Дж., Чен В. и др. (20 декабря 2019 г.). «Врожденные лимфоидные клетки: регуляторы барьерной функции кишечника и иммунного гомеостаза» . Журнал иммунологических исследований . 2019 : 2525984. дои : 10.1155/2019/2525984 . ПМЦ 6942837 . ПМИД 31930146 .
- ^ Нил Д.Р., Вонг С.Х., Беллоси А., Флинн Р.Дж., Дейли М., Лэнгфорд Т.К. и др. (апрель 2010 г.). «Нуоциты представляют собой новый врожденный эффекторный лейкоцит, который опосредует иммунитет 2 типа» . Природа . 464 (7293): 1367–70. Бибкод : 2010Natur.464.1367N . дои : 10.1038/nature08900 . ПМЦ 2862165 . ПМИД 20200518 .
- ^ Перейти обратно: а б с Миндт, Барбара С.; Фриц, Йорг Х.; Дюрр, Клаудия У. (30 апреля 2018 г.). «Группа 2 врожденных лимфоидных клеток в легочном иммунитете и гомеостазе тканей» . Границы в иммунологии . 9 : 840. дои : 10.3389/fimmu.2018.00840 . ISSN 1664-3224 . ПМК 5937028 . ПМИД 29760695 .
- ^ Перейти обратно: а б Халим Т.Ю., Стир К.А., Матха Л., Голд М.Дж., Мартинес-Гонсалес И., МакНагни К.М. и др. (март 2014 г.). «Врожденные лимфоидные клетки группы 2 имеют решающее значение для инициации адаптивного аллергического воспаления легких, опосредованного Т-хелперами 2» . Иммунитет . 40 (3): 425–35. doi : 10.1016/j.immuni.2014.01.011 . ПМК 4210641 . ПМИД 24613091 .
- ^ Ким Б.С., Сиракуза М.К., Саенс С.А., Ноти М., Монтичелли Л.А., Зонненберг Г.Ф. и др. (январь 2013 г.). «TSLP вызывает IL-33-независимые врожденные реакции лимфоидных клеток, способствующие воспалению кожи» . Наука трансляционной медицины . 5 (170): 170ра16. doi : 10.1126/scitranslmed.3005374 . ПМЦ 3637661 . ПМИД 23363980 .
- ^ Обоки К., Накае С., Мацумото К., Сайто Х. (апрель 2011 г.). «IL-33 и воспаление дыхательных путей» . Исследования аллергии, астмы и иммунологии . 3 (2): 81–8. дои : 10.4168/aair.2011.3.2.81 . ПМК 3062800 . ПМИД 21461246 .
- ^ Кондо Х., Итикава Ю., Имокава Г. (март 1998 г.). «Чрескожная сенсибилизация аллергенами через кожу с нарушенным барьером вызывает Th2-доминантный цитокиновый ответ» . Европейский журнал иммунологии . 28 (3): 769–79. doi : 10.1002/(SICI)1521-4141(199803)28:03<769::AID-IMMU769>3.0.CO;2-H . ПМИД 9541570 . S2CID 21654970 .
- ^ Бенезек, Сесиль; Джексон-Джонс, Люси Хелен (2019). «ILC2 Оркестрация функции местного иммунитета в жировой ткани» . Границы в иммунологии . 10 : 171. дои : 10.3389/fimmu.2019.00171 . ISSN 1664-3224 . ПМК 6374325 . ПМИД 30792718 .
- ^ Перейти обратно: а б с Бенезек С., Джексон-Джонс Л.Х. (2019). «ILC2 Оркестрация функции местного иммунитета в жировой ткани» . Границы в иммунологии . 10 : 171. дои : 10.3389/fimmu.2019.00171 . ПМК 6374325 . ПМИД 30792718 .
- ^ Джексон-Джонс Л.Х., Дункан С.М., Магальяес М.С., Кэмпбелл С.М., Майзелс Р.М., МакСорли Х.Дж. и др. (сентябрь 2016 г.). «Связанные с жиром лимфоидные скопления контролируют локальную секрецию IgM во время плевральной инфекции и воспаления легких» . Природные коммуникации . 7 (1): 12651. Бибкод : 2016NatCo...712651J . дои : 10.1038/ncomms12651 . ПМК 5025788 . ПМИД 27582256 .
- ^ Перейти обратно: а б Брестофф Дж.Р., Ким Б.С., Саенц С.А., Стайн Р.Р., Монтичелли Л.А., Зонненберг Г.Ф. и др. (март 2015 г.). «Врожденные лимфоидные клетки группы 2 способствуют образованию белой жировой ткани и ограничивают ожирение» . Природа . 519 (7542): 242–6. Бибкод : 2015Natur.519..242B . дои : 10.1038/nature14115 . ПМЦ 4447235 . ПМИД 25533952 .