Лимфопоэз
Тон или стиль этой статьи могут не отражать энциклопедический тон , используемый в Википедии . ( Ноябрь 2021 г. ) |
Лимфопоэз (lĭm'fō-poi-ē'sĭs) (или лимфоцитопоэз ) — образование лимфоцитов , одного из пяти типов лейкоцитов (лейкоцитов). [1] Более формально он известен как лимфоидное кроветворение .
Нарушение лимфопоэза может привести к ряду лимфопролиферативных заболеваний , таких как лимфомы и лимфоидные лейкозы .
| Словарь лимфопоэза |
|---|
| • антиген любая молекула, которая может спровоцировать иммунную защиту |
| • В-клетки- лимфоциты, которые в конечном итоге производят антитела |
| • костный мозг – центр костей, способный вырабатывать все красные и белые кровяные тельца у взрослого человека. |
| • кора — внешняя часть любого органа |
| • цитоплазма часть клетки между ядром и мембраной |
| • дифференциация, постоянные изменения в клетке, развивающиеся с течением времени и при делении клеток. |
| • гранулы, обнаруженные во многих лейкоцитах, состоящие из защитных химических веществ. |
| • кроветворные , которые дают начало любому типу клеток крови |
| • Происхождение типа клетки и ее потомков путем деления и дифференцировки. |
| • лимфоциты – особая «линия» лейкоцитов |
| • потомки миелоидных макрофагов (некоторые могут быть лимфоидными) со способностями к «еде» также взаимодействуют с лимфоцитами |
| • миелоидные предки лейкоцитов с гранулами, а также макрофагов |
| • Т-клетки «управляют» лимфоцитами для иммунитета |
| • (WBC) Лейкоциты в отличие от гораздо более распространенных эритроцитов; ответственный за оборону |
| редактировать |
Терминология
[ редактировать ]Лимфоциты — это клетки крови лимфоидного (а не миелоидного или эритроидного ) происхождения .

Лимфоциты находятся в кровотоке и происходят из костного мозга , однако преимущественно принадлежат к отдельной лимфатической системе , которая взаимодействует с кровообращением. [ нужна ссылка ]
Лимфопоэз сейчас обычно используется взаимозаменяемо с термином «лимфоцитопоэз» – образование лимфоцитов, но некоторые источники различают эти два понятия, заявляя, что «лимфопоэз» дополнительно относится к созданию лимфатической ткани , тогда как «лимфоцитопоэз» относится только к созданию клеток в эту ткань. В настоящее время лимфопоэз редко относится к созданию лимфатических тканей. [ нужна ссылка ]
Миелопоэз относится к «поколению клеток миелоидного ряда», а эритропоэз относится к «поколению клеток эритроидного ряда», поэтому появилось параллельное использование, в котором лимфопоэз относится к «поколению клеток лимфоидного ряда». [ нужна ссылка ]
Наблюдения за исследованиями, продолжавшимися более 100 лет, пролили свет на два класса лейкоцитов – миелоидные и лимфоидные – и в результате этих исследований были достигнуты успехи в медицине и науке. Изучив происхождение этих двух классов клеток, ученые выделили и определили два типа клеток с некоторыми сильными свойствами стволовых клеток — общий миелоидный предшественник (CMP) и общий лимфоидный предшественник (CLP) для мышей. [2] В конечном итоге было обнаружено, что эти прародители не были уникальными и что миелоидный и лимфоидный классы не были отдельными, а скорее двумя частично переплетенными генеалогическими древами. [ нужна ссылка ]
Функция
[ редактировать ]Зрелые лимфоциты являются важной частью иммунной системы, которые, за исключением В- и Т-клеток памяти , имеют короткую жизнь, измеряемую днями или неделями, и должны непрерывно генерироваться на протяжении всей жизни путем клеточного деления и дифференциации из таких клеток, как общие лимфоидные предшественники . CLP) у мышей. [ нужна ссылка ]
Набор, включающий клетки CLP и подобные предшественники, сами являются потомками плюрипотентных гемопоэтических стволовых клеток (pHSC), которые способны генерировать все типы клеток полной системы клеток крови. [3] Несмотря на свою способность генерировать полный набор лимфоцитов , большинство предшественников не являются настоящими стволовыми клетками и должны постоянно обновляться путем дифференциации из стволовых клеток pHSC. [4]
Многие клетки-предшественники также называются транзитными клетками . [5] иногда их также называют транзитными амплифицирующими клетками . Значение этого термина заключается в том, что транзитная клетка может найти новую сублинию, но количество образующихся клеток строго ограничено (хотя, возможно, очень велико, даже триллионы, но конечно), и линия прекращается клетки, которые отмирают (путем апоптоза ) или остаются клетками, которые больше не могут делиться . Примерами таких клеток являются КОЕ (колониеобразующие единицы, называемые таковыми из-за их способности образовывать колонии in vitro в искусственных средах), такие как КОЕ-Т. [6]
Трансплантация одной клетки pHSC может восстановить сублетально облученного хозяина (т.е. мышь, которая была облучена так, что все лейкоциты погибли) со всеми этими линиями клеток, включая все типы лимфоцитов через CLP.
Лимфопоэз продолжается на протяжении всей жизни, поэтому клетки-предшественники и их родительские стволовые клетки должны присутствовать всегда. [7]
Обзор
[ редактировать ]У млекопитающих, таких как человек ( Homo sapiens ), лимфопоэз начинается с ограниченного пассивного обеспечения со стороны матери. Сюда входят лимфоциты и иммуноглобулин G , которые проникают через плаценту и попадают в плод, обеспечивая некоторую защиту от патогенов , а также лейкоциты, которые поступают из грудного молока и попадают в кровоток через пищеварительный тракт. [8] Часто это неэффективно для предотвращения инфекций у новорожденного. [9]
Однако на ранних этапах беременности развивающийся эмбрион начинает собственный лимфопоэз из печени плода . Лимфопоэз возникает также из желточного мешка . [10] В этом отличие от взрослого человека, у которого все лимфоциты происходят из костного мозга. [11]
Существует четыре основных типа лимфоцитов, а также множество подтипов. Ученые определили сотни или тысячи типов клеток лимфоцитов , все из которых образуются в результате нормального или аномального лимфопоэза, за исключением некоторых искусственных штаммов, созданных в лабораториях путем развития существующих штаммов. Хотя лимфоциты обычно считаются зрелыми, как показывают анализы крови, они определенно не инертны. Лимфоциты могут путешествовать по организму везде, где в этом есть необходимость. Когда возникают такие потребности, могут произойти новые раунды нижестоящего лимфопоэза, такие как размножение и дифференцировка клеток, сопровождающиеся интенсивной митотической и метаболической активностью .
Это далеко не простая тема. В своей книге 1976 года «Иммунология, старение и рак» иммунолог и лауреат Нобелевской премии сэр Фрэнк Макфарлейн Бёрнет предположил, что однажды иммунная система может оказаться такой же сложной, как и нервная система . Поскольку производство лимфоцитов настолько близко к центральной роли иммунного ответа, разумно подойти к его изучению с некоторой смирением перед лицом этой задачи. Однако есть общие принципы, которые помогают в понимании.
Процесс
[ редактировать ]| Сокращения лимфопоэза | |
|---|---|
|
Лимфопоэз можно рассматривать в математическом смысле как рекурсивный процесс деления клеток, а также как процесс дифференцировки, измеряемый изменениями свойств клеток.
- Учитывая, что лимфоциты возникают из определенных типов ограниченных стволовых клеток, которые мы можем назвать P-клетками (от «прародитель»), такие клетки могут делиться несколькими способами. Это общие принципы ограниченного использования стволовых клеток. [12]
Считая P «материнской» клеткой, а не настоящей стволовой клеткой, она может разделиться на две новые клетки, которые сами по себе идентичны, но в некоторой степени отличаются от материнской. Или материнская клетка P может неравномерно делиться на две новые дочерние клетки, каждая из которых отличается друг от друга, а также от материнской.
Любая дочерняя клетка обычно обладает новыми специализированными способностями, и если она способна делиться, она образует новую подлинию. Отличие дочерней клетки от материнской может быть большим, но оно может быть и гораздо меньшим, даже едва заметным. делает материнская P-клетка, Чего не так это деления на две новые материнские P-клетки или на мать и дочь; это вопрос наблюдения, поскольку известно, что такие ограниченные клетки-предшественники не самообновляются.
- Существует своего рода исключение, когда дочерние клетки на каком-то уровне линии могут делиться несколько раз, образуя более внешне идентичные клетки, но тогда неизбежно происходит дальнейшая дифференциация и деление, пока не будет достигнута финальная стадия, на которой дальнейшее деление не может происходить. и линия клеточного типа наконец созрела. Примером зрелости являются плазматические клетки линии В-клеток, которые производят большое количество антител, но не могут делиться и в конечном итоге погибают через несколько дней или недель.
- Предшественник CLP мыши или предшественник MLP человека дифференцируется в лимфоциты, сначала становясь лимфобластами (Медицинская иммунология, стр. 10). Затем он делится еще несколько раз, образуя пролимфоцит, который имеет специфические маркеры клеточной поверхности, уникальные либо для (1) Т-клетки , либо (2) для В-клетки . Предшественник также может дифференцироваться в (3) естественные клетки-киллеры (NK) и (4) дендритные клетки (DC).
- Т-клетки, В-клетки и NK-клетки (и все другие врожденные лимфоидные клетки ) уникальны для семейства лимфоцитов, а дендритные клетки — нет. ДК, которые имеют одинаковый внешний вид, но имеют разные маркеры, распространены по всему телу и происходят как из лимфоидной, так и из миелоидной линии. Тем не менее, эти ячейки могут иметь несколько разные задачи и могут размещаться преимущественно в разных местах. [13] (Пересмотр с учетом новых исследований) Сейчас это открытый вопрос; кроме того, разные линии дендритных клеток могут иметь разные «задачи» или функции и находиться в разных «местах». [14]
Т- и В-лимфоциты неразличимы под микроскопом . Неактивные В- и Т-клетки настолько безлики, с небольшим количеством цитоплазматических органелл и в основном неактивным хроматином , что до 1960-х годов в учебниках можно было описать эти клетки, которые сейчас находятся в центре внимания иммунологии, как не имеющие известных функций! [15]
Однако Т- и В-лимфоциты представляют собой совершенно разные клеточные линии и «растут» в разных местах тела. Они выполняют в организме совершенно разные (хотя и совместные) функции. Никаких доказательств того, что Т- и В-клетки могут когда-либо взаимопревращаться, обнаружено не было. Т- и В-клетки биохимически различны, и это отражается в различных маркерах и рецепторах, которыми они обладают на поверхности клеток. Похоже, это справедливо для всех позвоночных, хотя между видами существует множество различий в деталях.
- Независимо от того, берут ли на себя ответственность за создание большого количества лимфоцитов CLP (мышь), MLP или небольшой близкородственный набор клеток-предшественников, одни и те же лимфоидные предшественники все же могут генерировать некоторые клетки, которые четко идентифицируются как миелоидные.
Лимфопоэз Т-клеток
[ редактировать ]Т-клетки образуются в костном мозге , а затем мигрируют в кору тимуса, где подвергаются созреванию в среде, свободной от антигенов, в течение примерно одной недели, где успех достигают лишь 2–4% Т-клеток. Остальные 96–98% Т-клеток погибают в результате и фагоцитируются макрофагами тимуса апоптоза . Так много тимоцитов (Т-клеток) погибают в процессе созревания, потому что проводится интенсивный скрининг, чтобы убедиться, что каждый тимоцит может распознавать собственный пептид: комплекс собственного MHC. [16] и за самотолерантность . Испытав апоптоз, тимоцит погибает и быстро перерабатывается.
По мере созревания существует несколько форм тимоцитов, в том числе [17]
- Т-хелпер (необходим для активации других клеток, таких как В-клетки и макрофаги),
- Т-цитотоксический (убивает инфицированные вирусом клетки),
- Т-память (Т-клетки, которые запоминают ранее встречавшиеся антигены) и
- Т-супрессорные клетки (которые смягчают иммунный ответ других лейкоцитов). Также называются Т-регуляторными клетками (Трег ) .
Когда Т-клетки активируются, они претерпевают дальнейшую серию изменений. Маленький покоящийся Т-лимфоцит быстро подвергается бластогенной трансформации в большой лимфоцит (13–15 мкм ). Этот крупный лимфоцит (известный в данном контексте как лимфобласт ) затем несколько раз делится, образуя расширенную популяцию средних (9–12 мкм) и малых лимфоцитов (5–8 мкм) с одинаковой антигенной специфичностью . [18] Окончательно активированные и дифференцированные Т-лимфоциты снова морфологически неотличимы от небольших покоящихся лимфоцитов. Таким образом, в анализах крови можно последовательно обнаружить следующие состояния развития:
- Пролимфоцит
- Большой лимфоцит
- Малый лимфоцит
Базовая карта Т-клеточного лимфопоэза
Эта базовая карта последовательного формирования Т-клеток упрощена и похожа на описания в учебниках и может не отражать последние исследования. (Медицинская иммунология, стр. 119)
В тимусе
- МЛП
- ЭТП
- DN1
- (Б; Мφ)
- Ду2
- (округ Колумбия; НК)
- DN3
- (Б-г)
- DN4
- ДП
- (ТНК; CD4; CD8; Т рег )
На периферии
- (Т ч 1; Т ч 2)
Развитие Т-клеток
[ редактировать ]В отличие от других лимфоидных линий, развитие Т-клеток происходит почти исключительно в тимусе. Т-лимфопоэз не происходит автоматически, а требует сигналов, генерируемых стромальными клетками тимуса. Определено несколько стадий, на которых для развития Т-клеток необходимы специфические регуляторы и факторы роста. На более позднем этапе развития и созревания Т-клеток те же самые регуляторные факторы снова используются для влияния на специализацию Т-клеток. [ нужна ссылка ]
Т-клетки уникальны среди популяций лимфоцитов своей способностью далее специализироваться как зрелые клетки и становиться еще более зрелыми. Т-клетки бывают разных видов, например: обычные Т-клетки TcRαβ; так называемые нетрадиционные Т-клетки TcRγδ; NKT-клетки; и Т-регуляторные клетки (Treg ) . Детали, касающиеся развития и жизненного цикла нетрадиционных Т-клеток, менее подробно описаны по сравнению с обычными Т-клетками. [ нужна ссылка ]
Стадии созревания Т-клеток
Этап первый: миграция тимуса
Мультипотентные лимфоидные предшественники (MLP) входят в путь Т-клеток по мере их миграции в тимус. Наиболее примитивными клетками тимуса являются ранние предшественники тимоцитов (ETP), которые сохраняют весь лимфоидный и миелоидный потенциал, но существуют лишь временно, быстро дифференцируясь в линии T и NK. (Медицинская иммунология, стр. 118)
Этап второй: пролиферативная экспансия и приверженность T-линии
Окончательная приверженность линии Т-клеток происходит в микроокружении тимуса, микроскопических структурах тимуса, где питаются Т-клетки. Наиболее примитивные Т-клетки сохраняют мультипотентную способность и могут дифференцироваться в клетки миелоидного или лимфоидного ряда (В-клетки, ДК, Т-клетки или NK-клетки).
Более дифференцированные двойные негативные Т-клетки (клетки DN2) имеют более ограниченный потенциал, но еще не полностью ограничены линией Т-клеток (они все еще могут развиваться в DC, Т-клетки или NK-клетки). Позже они полностью связываются с линией Т-клеток - когда тимоциты, экспрессирующие рецепторы Notch1, взаимодействуют с стромальными клетками тимуса, экспрессирующими лиганды Notch1, тимоциты окончательно становятся приверженными линии Т-клеток. См. изображение галереи «Двойные негативы».
С принятием линии Т-клеток начинается очень сложный процесс, известный как реаранжировка гена TCR . Это создает огромное разнообразие Т-клеток, несущих антигенные рецепторы. После этого некоторые Т-клетки покидают тимус и мигрируют в кожу и слизистые оболочки .
Третий этап: β-отбор
Этап четвертый: выбор рецепторов Т-клеток
Только 2–3% дифференцирующихся тимоцитов, экспрессирующих TcR, способных взаимодействовать с молекулами MHC, но толерантных к аутопептидам, [ нужны дальнейшие объяснения ] выжить в процессе отбора на четвертом этапе.
Этап пятый: продолжение дифференциации на периферии
Ранее считалось, что тимус человека остается активным местом дифференцировки Т-клеток только до раннего взросления и что позже во взрослой жизни тимус атрофируется , возможно, даже исчезает. Недавние сообщения показывают, что тимус человека активен на протяжении всей взрослой жизни. Таким образом, несколько факторов могут способствовать поставке Т-клеток во взрослой жизни: генерация в тимусе, внетимическая дифференцировка и тот факт, что Т-клетки памяти долгоживущие и выживают в течение десятилетий.
Типы Т-клеток
[ редактировать ]- Нетрадиционные Т-клетки
Тимус также дает начало так называемым «нетрадиционным Т-клеткам», таким как γδ Т-клетки, естественные Т-клетки-киллеры (NKT) и регуляторные Т-клетки (Treg ) . [ нужна ссылка ]
- γδ Т-клетки
γδT-клетки составляют лишь от 1% до 5% циркулирующих Т-клеток, но их много в иммунной системе слизистых оболочек и коже, где они представляют собой доминирующую популяцию Т-клеток. Эти «не-MHC-рестриктированные Т-клетки» участвуют в специфических первичных иммунных реакциях, надзоре за опухолями, иммунной регуляции и заживлении ран. [19]
Описано несколько различий между развитием αβ и γδ Т-клеток. Они эмигрируют из тимуса «волнами» клональных популяций, в которых обитают отдельные ткани. Например, один вид обнаруживается в периферической крови, а другой преобладает в кишечном тракте.
- Естественные Т-киллеры
NKT-клетки человека представляют собой уникальную популяцию и, как полагают, играют важную роль в опухолевом иммунитете. [20] и иммунорегуляция .
- Т-регуляторные клетки
Т- рег- клетки считаются встречающимися в природе регуляторными Т-клетками. Трег составляют около 5% циркулирующего CD4. + Т-клетки. Считается, что эти клетки обладают важным аутоиммунным свойством, регулируя «аутореактивные» Т-клетки на периферии . (Медицинская иммунология, стр. 117-122)
Лимфопоэз В-клеток
[ редактировать ]В-клетки формируются и созревают в костном мозге (и селезенке).
То, что В-клетки образуются в костном мозге, является хорошим подспорьем, но это простое совпадение, поскольку В-клетки впервые были изучены в куриной сумке Фабрициуса , и именно от этой бурсы В-клетки получили свое название. [ нужна ссылка ]
Эти В-клетки затем покидают костный мозг и мигрируют через кровоток и лимфу в периферические лимфоидные ткани, такие как селезенка, лимфатические узлы, миндалины и ткани слизистых оболочек. Попав во вторичный лимфоидный орган, В-клетка может быть введена в контакт с антигеном, который она способна распознать. [ нужна ссылка ]
Благодаря распознаванию антигена и другим клеточным взаимодействиям В-клетка активируется, а затем делится и дифференцируется, образуя плазматическую клетку. Плазматическая клетка, конечный продукт В-клеток, представляет собой очень активную клетку, секретирующую антитела , которая помогает защитить организм, атакуя антиген и связываясь с ним.
Даже после многих десятилетий исследований остаются некоторые разногласия относительно того, где В-клетки созревают и «завершают свое образование», при этом сохраняется вероятность того, что этим местом также могут частично быть перикишечные лимфоидные ткани . [21]
В-лимфопоэз происходит исключительно в костном мозге, и В-лимфоциты производятся там непрерывно на протяжении всей жизни в «микроокружении», состоящем из стромальных клеток , внеклеточного матрикса , цитокинов и факторов роста, которые имеют решающее значение для пролиферации, дифференциации и выживания ранних лимфоцитов и В-лимфоцитов. -предшественники рода.
Относительная доля В-клеток-предшественников в костном мозге остается довольно постоянной на протяжении всей жизни организма. Существуют такие стадии, как клетки Pre-BI (от 5% до 10% от общего числа); Клетки Pre-B-II (60–70%), а остальные 20–25% представляют собой незрелые В-клетки. В большинстве учебников говорится, что В-клетки созревают в костном мозге, но, как правило, незрелые В-клетки мигрируют в селезенку для получения своего рода «высшего образования», где они проходят переходные стадии перед окончательным созреванием. (Медицинская иммунология, стр. 136)
В-лимфоциты идентифицируются по наличию растворимого иммуноглобулина G (IgG). Это самый распространенный защитный иммуноглобулин в организме взрослого человека. После антигенной стимуляции В-клетки дифференцируются в плазматические клетки, секретирующие большие количества растворимого IgG. Это заключительная стадия B-лимфопоэза, но она является решающей, поскольку плазматические клетки должны либо вырабатывать антитела вблизи источника инфекции, либо распространять их в крови для борьбы с инфекцией на расстоянии или в недоступной части тела.
Базовая карта В-клеточного лимфопоэза
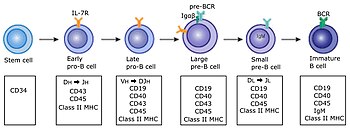
Общепризнанная действительная карта В-клеточного лимфопоэза состоит из двух частей, первая из которых находится в костном мозге, а вторая - в селезенке: [22] Процесс развития в костном мозге происходит в зародышевых центрах
В костном мозге
- Про-Б
- Предварительный BI
- Pre-B-II большой
- Pre-B-II маленький
- Незрелый)
В селезенке
- Т1
- Т2/Т3
- (Маргинальная зона (МЗ); Б-1; Б-2)
- B-2 далее подразделяется на:
- (Герминальный центр (ГЦ); Память; Плазма)
Лимфопоэз NK-клеток
[ редактировать ]NK-клетки, у которых отсутствуют антигенспецифические рецепторы, развиваются в костном мозге. После созревания и выхода из костного мозга они всю жизнь циркулируют в крови в поисках новых возможностей. Возможность, которую они ищут, состоит в том, чтобы встретиться, распознать, а затем уничтожить аномальные клетки, такие как раковые или инфицированные вирусом клетки. Хорошо известно, что лимфоциты никогда не имеют гранул или, по крайней мере, гранул, которые хорошо видны даже при окрашивании . Однако NK-клетки являются исключением. У них действительно есть многочисленные гранулы, которые обеспечивают их способность убивать клетки , и именно благодаря этим гранулам NK-клетки имеют альтернативное название — большие гранулярные лимфоциты (LGL). [ нужна ссылка ]
NK-клетки — единственные лимфоциты, которые считаются частью врожденной иммунной системы (в отличие от адаптивной иммунной системы ). Однако они гораздо более тесно связаны с Т-клетками (частью адаптивной иммунной системы ), чем с другими клетками врожденной иммунной системы. NK-клетки не только имеют много общих с Т-клетками поверхностных маркеров, функций и активностей, они также возникают из общего предшественника T/NK. Считается, что предшественник T/NK также является источником субпопуляции лимфоидных DC (Medical. Иммунология, стр. 121)
NK-клетки имеют определение «штрих-код» как CD3, CD16+, CD56t-лимфоциты. (См. раздел «Штрих-код» в этой статье). Предшественники NK можно обнаружить главным образом в тимусе (мыши), но тимус не является абсолютно необходимым для развития NK. Вероятно, NK-клетки могут развиваться в различных органах, но основной сайт развития NK-клеток неизвестен. [ нужна ссылка ]
У человека большинство (85–90%) NK-клеток обладают высокой цитолитической способностью (способностью лизировать клетки). Меньшая подгруппа (10–15%), называемая NK «CD56 Bright», в основном отвечает за выработку цитокинов и повышает выживаемость . Двигаясь к лимфатическим узлам, «яркие CD56» NK-клетки снова дифференцируются в зрелые NK-клетки, которые экспрессируют иммуноглобулиноподобные рецепторы киллеров (KIR), естественные рецепторы цитотоксичности (NCR) и критические молекулы адгезии . (Медицинская иммунология, стр. 122)
Лимфопоэз дендритных клеток
[ редактировать ]Процесс, посредством которого клетки CLP могут дифференцироваться с образованием дендритных клеток лимфоидного происхождения, еще четко не определен. [23]
ДК представляют собой высокоспециализированные и эффективные антигенпрезентирующие клетки . Клетки, идентичные по внешнему виду, происходят как из миелоидной линии (называемой миелоидными дендритными клетками), так и из лимфоидной линии (называемой плазмоцитоидными дендритными клетками ). [ нужна ссылка ]
Развитие и регуляция ДК изучены недостаточно. Хотя предшественники ДК были идентифицированы в печени, тимусе и костном мозге плода человека, считается, что во время взрослой жизни ДК производятся только из костного мозга и высвобождаются в кровь, чтобы перемещаться и оседать. В целом, большое количество ДК различных типов распределяется по всему телу, особенно в эпителиях, таких как кожа, для отслеживания захватчиков и уничтожения их антигенов. (Медицинская иммунология, стр. 122)
Сравнение киллеров лимфопоэза
[ редактировать ]Лимфоциты обладают рядом тревожных свойств, таких как способность перемещаться по телу и занимать ночлег практически где угодно, и по пути отдавать команды в виде цитокинов , хемокинов и лимфокинов , команды, которые влияют на многие типы клеток в организме и который также может рекурсивно индуцировать дальнейший лимфопоэз. Одна из сильных моделей поведения, которая очаровывает как исследователей, так и общественность, — это способность лимфоцитов действовать как полицейский, судья и палач, убивая другие клетки или требуя от них самоубийства — команда, которой обычно подчиняются. Другого варианта приговора, похоже, не существует. [ нужна ссылка ]
Киллеры отличаются от таких клеток, как макрофаги, которые поедают другие клетки или пережевывают остатки, с помощью метода, называемого фагоцитозом . Киллеры не используют фагоцитоз; они просто убивают и оставляют зачистку другим клеткам.
Известно, что убийцы атакуют инфицированные вирусом клетки и клетки, ставшие раковыми. Благодаря этим способностям было проведено много исследований по использованию этих качеств в медицинской терапии, но прогресс был медленным.
Вот парад убийц и как они работают:
- Цитотоксические Т-клетки
(также называемые Tc или антигенспецифическими цитолитическими или -токсичными Т-лимфоцитами ( CTL) ). [24] Tc убивает путем апоптоза и либо обрызгивает цель перфорином или гранзимами , либо использует взаимодействие Fas-Fasl для управления уничтожением цели. Это убивает клетки, которые инфицированы и содержат антиген.
- NK-клетки (также называемые LGL (большие гранулярные лимфоциты))
Они убивают точно так же, как и Tc, но не взаимодействуют ни с каким антигеном. Они выбирают свои цели на основе типичных молекул, отображаемых клетками, испытывающими стресс из-за вирусной инфекции. NK-клетки в основном находятся в кровообращении (5-15% циркулирующих лимфоцитов), но также повсеместно распределены в тканях. [25]
- Клетки LAK (киллеры, активируемые лимфокинами) представляют собой лабораторную/клиническую подгруппу NK-клеток, которым IL-2 способствует атаке опухолевых клеток. [26]
- NKT-клетки см. Т-клетки натуральных киллеров» . в основной статье «
Естественные Т-клетки-киллеры. NK-T-клетки человека представляют собой уникальную популяцию (которая экспрессирует маркеры NK-клеток, такие как CD56 и KIR). Считается, что NKT-клетки играют важную роль в опухолевом иммунитете и иммунорегуляции (Медицинская иммунология, стр. 135), однако об этом мало что известно. Недавние данные свидетельствуют о том, что звездчатые клетки печени играют роль резидентных антигенпрезентирующих клеток, которые представляют липидные антигены и стимулируют пролиферацию NKT-клеток.
- Естественные Т-клетки, подобные киллерам
Гетерогенная группа с плохо определенными свойствами.
Однако, вкратце, не существует известной клетки или набора клеток , способных убивать раковые клетки в целом.
Маркировка лимфопоэза
[ редактировать ]Поскольку все лейкоциты микроскопичны, бесцветны и часто кажутся идентичными по внешнему виду, их индивидуально идентифицируют по природным химическим маркерам, многие из которых были проанализированы и названы. Когда две клетки имеют одинаковые маркеры, делается разумное предположение, что в этот момент клетки идентичны. Набор маркеров в просторечии описывается как штрих-код для этой клетки или этой клеточной линии.
- важнейшей гемопоэтической стволовой клетки (HSC). Вот пример того, как может возникнуть штрих-код, на примере [ нужна ссылка ]
Технически HSC описываются как: лишенные FMS-подобной тирозинкиназы 3 (Flt3) и маркеров, специфичных для отдельных лимфоидных линий (Lin), но экспрессирующих высокие уровни Sca1 и c-kit; HSC также экспрессируют CD44, низкие уровни Thy1.1 (CD90), но не экспрессируют IL-7Ra или CD27. [ нужна ссылка ]
Это называется (поверхностным) фенотипом HSC. Он может быть выражен в виде набора (Lin2, Sca1high, c-kit high, CD44+, Thy1.1low, CD272 и IL-7Ra2). Этот набор представляет собой «штрих-код» для HSC, аналог этикетки со штрих-кодом, прикрепленной к пластиковому пакету с куриными крылышками для кассы в супермаркете! Ученые используют эти штрих-коды для проверки, классификации и накопления клеток для многих целей, часто используя лабораторные методы, такие как проточная цитометрия клеток . коды частично определяют современное значение фенотипа лейкоцитов Эти штрих - .
Прогресс дифференцировки HSC и детерминации клонов указывается изменениями в этом фенотипе . То есть по мере изменения ячейки будут меняться и маркеры, и штрих-код.
- Типичные штрих-коды для некоторых типов клеток, представленных в этой статье.
| Тип ячейки | Штрих-код |
|---|---|
| ЭТП | C-Kit+, CD44+, CD25- |
| DN1 | CD44+, CD25- |
| Ду2 | CD44+, CD25+ |
- Примечание, поясняющее детали параметров штрих-кода: Flt3 представляет собой рецептор цитокина тирозинкиназы, который считается важным на раннем этапе развития лимфоидных клеток. Кроме того, Flt3 играет важную роль в поддержании B-лимфоидных предшественников. CD27 играет роль в лимфоидной пролиферации, дифференцировке и апоптозе. Приобретение CD27 и Flt3 HSC совпадает с потерей долгосрочного потенциала репопуляции. На этой стадии клетки сохраняют как лимфоидный, так и миелоидный потенциал и называются мультипотентными предшественниками. (Медицинская иммунология, стр. 114)
Развитие знаний о лимфопоэзе
[ редактировать ]В иммунологии постоянно возникают новые вопросы, как если бы для вопросов существовала стволовая клетка . Например, считалось, что процесс лимфопоэза представляет собой прямую, упорядоченную однонаправленную последовательность. Но неясно, происходят ли лимфоциты конечной стадии из предшественников, которые представляют собой однородные популяции или перекрывающиеся популяции. Неясно также, развиваются ли линии лимфоцитов посредством континуума дифференцировки с прогрессирующей утратой вариантов линии или же резкие события приводят к приобретению определенных свойств. [27]
Изменения цитоплазмы, морфологии ядра клетки, гранул, внутренней биохимии клетки, сигнальных молекул и маркеров клеточной поверхности трудно коррелировать с определенными этапами лимфопоэза. Морфологические различия не просто соответствуют этапам митоза (деление соматической клетки), но являются результатом непрерывных «процессов созревания» ядра клетки, а также цитоплазмы, поэтому не следует быть слишком жесткими в отношении морфологических различий между определенными клетками. этапы. [28]
- Модели и обновления генеалогического древа лимфопоэза
До недавнего времени модель CMP, генерирующая все миелоидные клетки, и CLP, генерирующая все лимфоидные клетки, считалась необходимой и достаточной для объяснения известных фактов, наблюдаемых при образовании лейкоцитов, и она до сих пор встречается в большинстве основных учебников. Однако начиная примерно с 2000 г. и набирая обороты после 2005 г. [29] в обоих исследованиях на людях и мышах были отмечены и опубликованы в статьях новые сложности. [ нужна ссылка ] Эти исследования сейчас важны в основном для исследователей-иммунологов, но, вероятно, в конечном итоге приведут к изменениям в методах лечения.
Изменения были вызваны наблюдениями о том, что лимфопоэз не всегда распадается на две линии на уровне CLP. Хуже того, некоторые макрофаги (долгое время считавшиеся миелоидной линией) могут генерироваться предшественниками лимфоидной линии. По сути, фокус был смещен с CLP на MLP. [ нужна ссылка ] (лимфоидные предшественники), которые явно являются лимфоидными предшественниками, но сохраняют некоторый миелоидный потенциал, в частности, способность как у людей, так и у мышей вырабатывать макрофаги – одни из наиболее универсальных защитников иммунных клеток – а также множество дендритных клеток, лучших «сторожевых псов» антигенных захватчиков.
Однако какими бы ни были подробности, процесс лимфопоэза всегда, по-видимому, неустанно порождает потомство с особыми качествами и способностями – так сказать, «сверхспособностями», – но со все более ограниченным лимфоидным потенциалом развития.
Этапы развития
[ редактировать ]Старая модель: лимфоидная или миелоидная.
[ редактировать ]Эта модель лимфопоэза отличалась относительной простотой, соответствием номенклатуре и терминологии и до сих пор по существу актуальна для лабораторных мышей.
- pHSC Плюрипотентные самообновляющиеся гемопоэтические стволовые клетки [30] которые порождают
- Мультипотентные предшественники MPP , дающие начало
- ELP (или PRO) Пролимфоциты , ранние лимфоидные предшественники и, наконец,
- CLP Общий лимфоидный предшественник, тип клеток, полностью принадлежащий к лимфоидной линии.
Клетки pHSC, MPP и ELP не полностью связаны с лимфоидной линией, поскольку, если их удалить в другое место, они могут дифференцироваться в нелимфоидное потомство. Однако CLP придерживаются лимфоидной линии. CLP — это транзитная ячейка, ответственная за эти (как правило, параллельные) стадии развития, представленные ниже:
- NK-клетки
- Дендритные клетки (лимфоидная линия; DC2 [31] )
- В-клетки-предшественники
- Про-В-клетки => Ранние Про-(или пре-пре)-В-клетки => Поздние Про-(или пре-пре)-В-клетки
- Большие клетки Pre-B => маленькие клетки Pre-B
- Незрелые В-клетки
- B-клетки => (клетки B1; клетки B2)
- Плазматические клетки
- Про-Т-клетки
- Т-клетки
Исследование новых моделей (не мышей)
[ редактировать ]К 2008 году было обнаружено, что «большинство ранних клеток-предшественников тимуса [ETP] не превращаются в Т-клетки к тому времени, когда они попадают в вилочковую железу. ETP-клетки сохраняют способность становиться либо T-клетками, либо миелоидными клетками». [32] [33]
См. также: [34] [35] [36] [37] [38] [39]
Графическое представление старой модели по сравнению со смешанной лимфоидной и миелоидной моделью
[ редактировать ] Бок о бок. Сравнение новых и старых моделей линейки.
Бок о бок. Сравнение новых и старых моделей линейки. Пересмотренная блок-схема лимфоидных и миелоидных линий.
Пересмотренная блок-схема лимфоидных и миелоидных линий.


Справочные материалы по общей иммунологии
[ редактировать ]Тексты, выделенные жирным шрифтом , наиболее часто цитируются в этой статье.
- Клеточная коммуникация в нервной и иммунной системе ; Гундельфингер, Зайденбехер, Шравен; Спрингер Берлин Гейдельберг Нью-Йорк; 2006 г.; ISBN 3-540-36828-0
- Цветной атлас гематологии ; Темл и др.; Тиме; 2004 г.; ISBN 1-58890-193-9
- Динамика рака ; Стивен А. Франк; Издательство Принстонского университета, Принстон, Нью-Джерси; 2007 г.; ISBN 978-0-691-13366-9 , общественная лицензия Creative Commons
- Фундаментальная иммунология , 5-е издание; Уильям Э. Пол (редактор); Издательство Липпинкотт Уильямс и Уилкинс; 2003 г.; ISBN 0-7817-3514-9
- Иммунобиология: Иммунная система в здравоохранении и науке , 6-е издание; Джейнвей, Трэверс; 2005 г.; Издательство Garland Science, Нью-Йорк; ISBN 0-8153-4101-6
- Вводный учебник по иммунологии (электронная книга; переработанное 2-е издание); Нандини Шетти; New Age International (P) Limited, Паблишерс, Индия; 2005 г.; ISBN 81-224-2335-3
- Мгновенные заметки по иммунологии , 2-е изд.; Лидьярд, Уилан, Фэнджер; Группа Тейлор и Фрэнсис; 2004 г.; Китайская версия ISBN 978-7-03-025225-8 ; 46 юаней Книжный магазин Ванфуцзин
- Медицинская иммунология —6-е изд.; Г. Вирелла, редактор; Информа Хелскейр США, Инк; 2007 г.; ISBN 978-0-8493-9696-0
- Биология стволовых клеток ; Маршак, Гарднер, Готлиб; Лабораторный пресс Колд-Спринг-Харбор; 2001 г.; ISBN 0-87969-575-7 /01
- Учебник развития человека и гистологии ; Чжун Цуйпин и др.; Шанхайское научно-техническое издательство; 2006 г.; ISBN 7-5323-8230-3
- Учебник медицинской иммунологии (Иммунология, 7-е издание); ЛИМ Пак Леонг; «Элзевир (Сингапур) Пте Лтд.»; 2006 г.; ISBN 0-323-03399-7
Ссылки
[ редактировать ]- ^ Бирбрайр, Александр; Френетт, Пол С. (01 марта 2016 г.). «Неоднородность ниш в костном мозге» . Анналы Нью-Йоркской академии наук . 1370 (1): 82–96. Бибкод : 2016NYASA1370...82B . дои : 10.1111/nyas.13016 . ISSN 1749-6632 . ПМЦ 4938003 . ПМИД 27015419 .
- ^ Бирбрайр, Александр; Френетт, Пол С. (01 марта 2016 г.). «Неоднородность ниш в костном мозге» . Анналы Нью-Йоркской академии наук . 1370 (1): 82–96. Бибкод : 2016NYASA1370...82B . дои : 10.1111/nyas.13016 . ISSN 1749-6632 . ПМЦ 4938003 . ПМИД 27015419 .
- ^ Биология стволовых клеток , стр. 307.
- ^ Бирбрайр, Александр; Френетт, Пол С. (01 марта 2016 г.). «Неоднородность ниш в костном мозге» . Анналы Нью-Йоркской академии наук . 1370 (1): 82–96. Бибкод : 2016NYASA1370...82B . дои : 10.1111/nyas.13016 . ISSN 1749-6632 . ПМЦ 4938003 . ПМИД 27015419 .
- ^ Динамика рака , стр. 251.
- ^ «КФУ-Т» . Архивировано из оригинала 26 мая 2023 г. Проверено 24 августа 2010 г.
- ^ Бирбрайр, Александр; Френетт, Пол С. (01 марта 2016 г.). «Неоднородность ниш в костном мозге» . Анналы Нью-Йоркской академии наук . 1370 (1): 82–96. Бибкод : 2016NYASA1370...82B . дои : 10.1111/nyas.13016 . ISSN 1749-6632 . ПМЦ 4938003 . ПМИД 27015419 .
- ^ Ребер А.Дж., Донован, округ Колумбия, Габбард Дж., Галланд К., Асевес-Авила М., Холберт К.А., Маршалл Л., Херли DJ (2008). «Перенос материнских колостральных лейкоцитов способствует развитию иммунной системы новорожденного. Часть II. Влияние на неонатальные лимфоциты». Вет Иммунол Иммунопатол . 123 (3–4): 305–13. дои : 10.1016/j.vetimm.2008.02.009 . ПМИД 18394715 .
- ^ Прихубер, Глория С. (2015). «Послеродовые инфекции и иммунология, влияющие на хронические заболевания легких недоношенных» . Клиники перинатологии . 42 (4): 697–718. дои : 10.1016/j.clp.2015.08.002 . ISSN 0095-5108 . ПМК 4660246 . ПМИД 26593074 ; Доступ предоставлен Питтсбургским университетом.
{{cite journal}}: CS1 maint: постскриптум ( ссылка ) - ^ Предшественники взрослых Т-клеток сохраняют миелоидный потенциал; Природа ; 2008 год
- ^ Учебник человеческого развития и гистологии , стр.176.
- ^ Бирбрайр, Александр; Френетт, Пол С. (01 марта 2016 г.). «Неоднородность ниш в костном мозге» . Анналы Нью-Йоркской академии наук . 1370 (1): 82–96. Бибкод : 2016NYASA1370...82B . дои : 10.1111/nyas.13016 . ISSN 1749-6632 . ПМЦ 4938003 . ПМИД 27015419 .
- ^ Иммунобиология, Иммунная система в здравоохранении и науке .
- ^ Фундаментальная иммунология, 5-е издание.
- ^ Иммунобиология, Иммунная система в здравоохранении и науке . Гарленд Научное Издательство
- ^ Чиофани, Мария; Суньига-Пфлюкер, Хуан Карлос (1 января 2007 г.). «Тимус как индуктивный участок Т-лимфопоэза» . Ежегодный обзор клеточной биологии и биологии развития . 23 : 463–493. doi : 10.1146/annurev.cellbio.23.090506.123547 . ISSN 1081-0706 . ПМИД 17506693 . Архивировано из оригинала 3 сентября 2021 г. Проверено 3 июля 2019 г.
- ^ Учебник медицинской иммунологии , стр. 5.
- ^ Медицинская иммунология , стр. 23.
- ^ «Гамма-дельта (γδ) Т-клетки | Британское общество иммунологии» . www.immunology.org . Архивировано из оригинала 2 января 2020 г. Проверено 14 июня 2019 г.
- ^ «Опухолевый иммунитет и иммунотерапия рака» . Архивировано из оригинала 27 августа 2010 г. Проверено 25 августа 2010 г.
- ^ Медицинская иммунология , стр. 22.
- ^ Медицинская иммунология , с. 123
- ^ Фундаментальная иммунология ; Пол; Ч. 15 «РАСПРЕДЕЛЕНИЕ ДЕНДРИТИЧЕСКИХ КЛЕТОК IN VIVO: МНОГОЧЛЕННОЕ СЕМЬЕ»
- ^ Иммунология ; Лидьярд и др.; п. 22, 132-137
- ^ Иммунология ; Лидьярд и др.; п. 15, 18-20,41
- ^ Иммунология ; Лидьярд и др.; п. 20, 259-260
- ^ Медицинская иммунология ; Литвин, с. 122
- ^ Цветной атлас гематологии 2004 г.
- ^ «Самые ранние тимические предшественники Т-клеток обладают потенциалом миелоидного происхождения»; Белл, Бхандула; Том 452, 10 апреля 2008 г., дои : 10.1038/nature06840
- ^ Медицинская иммунология , Литвин, стр.115.
- ^ Учебник медицинской иммунологии , стр. 31.
- ^ Результаты исследований могут пролить свет на Т-клеточные лейкозы и иммунодефициты. Архивировано 13 февраля 2010 г. в Wayback Machine Bhandoola. 9 апреля 2008 г.;
- ^ Перерисованы линии крови; Томас Граф; Nature Vol 452 10 апреля 2008 г. стр.702-703
- ^ Общий миелолимфоидный предшественник: ключевая промежуточная стадия гемопоэза, генерирующая Т- и В-клетки; Мин Лу, Хироши Кавамото, Ёсихиро Кацубе, Томокацу Икава и Ёсимото Кацура; Дж. Иммунол. 2002;169;3519-3525
- ^ Идентификация Flt3 + лимфо-миелоидных стволовых клеток, лишенных эритро-мегакариоцитарного потенциала: пересмотренная дорожная карта для приверженности линии крови взрослых; Лундский центр стратегических исследований биологии стволовых клеток; Клетка; Том. 121, 295–306, 22 апреля 2005 г.
- ^ Взрослые предшественники Т-клеток сохраняют миелоидный потенциал Харука Вада, Кёко Масуда, Руми Сато, Киёкадзу Какугава, Томокацу Икава, Ёсимото Кацура и Хироши Кавамото, том 452, 10 апреля 2008 г.;
- ^ Самые ранние предшественники Т-клеток тимуса обладают потенциалом миелоидного происхождения; Дж. Джеремия Белл, Авинаш Бхандула; Природа ; Том 452, 10 апреля 2008 г., с. 764-768
- ^ Пересмотренная карта иерархии предшественников человека показывает происхождение макрофагов и дендритных клеток на раннем этапе развития лимфоидных клеток; Дик и др.; Природная иммунология ; Том 11 Номер 7 Июль 2010 г. с. 585-595
- ^ Не раздельное решение для кроветворения человека; Кеннет Доршкинд; Природная иммунология, том 11, номер 7, июль 2010 г. с. 569-570
Дополнительные изображения
[ редактировать ]Альтернативные взгляды на родословную
 Линия клеток крови. Для масштаба обратите внимание, что мегакариоциты (50–100 мкм) в 10–15 раз больше типичных эритроцитов.
Линия клеток крови. Для масштаба обратите внимание, что мегакариоциты (50–100 мкм) в 10–15 раз больше типичных эритроцитов. Линия клеток крови. На основе способности к самообновлению.
Линия клеток крови. На основе способности к самообновлению. Схематический вид. Четко определенные родословные.
Схематический вид. Четко определенные родословные.- Бок о бок. Сравнение новых и старых моделей линейки.



Внешние ссылки
[ редактировать ]- Мини-портал www.copewithcytokines.de, посвященный терминологии лимфопоэза.
- Лимфопоэз в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- «Лимфопоэз» в Медицинском словаре Дорланда.
- Обзор на hematologica.pl