Фолликулярные В-хелперные Т-клетки
Фолликулярные Т-хелперы (также известные как Т-фолликулярные хелперные клетки и сокращенно TFH ) представляют собой антиген -экспериментированные CD4. + Т-клетки обнаруживаются на периферии внутри В-клеточных фолликулов вторичных лимфоидных органов, таких как лимфатические узлы , селезенка и пейеровы бляшки , и идентифицируются по их конститутивной экспрессии В-клеточных рецептора самонаведения фолликулов CXCR5 . [1] При клеточном взаимодействии и передаче перекрестных сигналов со своими родственными фолликулярными (Fo B) B-клетками T FH- клетки запускают образование и поддержание зародышевых центров посредством экспрессии лиганда CD40 (CD40L) и секреции IL-21. [2] и Ил-4 . [3] Клетки Т -СГ также мигрируют из зон Т-клеток в эти засеянные зародышевые центры , преимущественно состоящие из быстро делящихся В-клеток, мутирующих в своих генах Ig. Внутри зародышевых центров Т -ФГ- клетки играют решающую роль в обеспечении отбора и выживания В-клеток, которые затем дифференцируются либо в долгоживущие плазматические клетки, способные продуцировать высокоаффинные антитела против чужеродного антигена, либо в зародышевые центрозависимые В-клетки памяти. способны к быстрой реактивации иммунитета в будущем, если тот же антиген когда-либо встретится повторно. [4] Т- ФГ- Считается также, что клетки способствуют негативному отбору потенциально аутоиммунных мутировавших В-клеток в зародышевом центре . Однако биомеханизмы, с помощью которых клетки Т -ФГ опосредуют толерантность зародышевого центра , еще полностью не изучены.
Вполне возможно, что клетки T FH могут возникнуть как ветви путей дифференцировки Th1 и Th2, но их точное родство с другим эффекторным CD4 + Подмножества Т-клеток все еще не определены. Однако исследования показали, что TFH имеют разные профили экспрессии генов, что подтверждает теорию о том, что TFH являются подмножеством CD4. + Т-клетки отличаются от Th-1 , Th-2 , Th-17 или Treg . [5] [6]
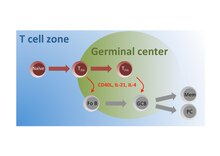
Подмножество наивных Т-клеток в зоне Т-клеток активируется антигеном и мигрирует в фолликулы, где они дифференцируются в Т- клетки FH , которые взаимодействуют с фолликулярными B-клетками (Fo B) и инструктируют их о переключении изотипа, соматической гипермутации и быстром клеточном разрушении. деление на зародышевые центры семян (ГЦ). Внутри этих зародышевых центров клетки T FH продолжают оказывать помощь GC B-клеткам, способствуя выработке ими высокоаффинных антител, продуцирующих плазматические клетки (PC) и B-клетки долгоживущей памяти (Mem).
Биомолекулярная характеристика
[ редактировать ]- ФГ , поскольку экспериментальные мыши Доказано, что индуцируемый костимулятор Т-клеток (CD278 или ICOS) обеспечивает особенно критический сигнал для клеток Т с дефицитом ICOS не способны развивать какой-либо Т -ФГ . [7] Кроме того, было показано, что ICOS индуцирует секрецию цитокина IL-21 активированным CD4. + Т-клетки и что IL-21 играет решающую роль в развитии Т- ФГ- клеток и зародышевых центров . [8] [9] Кроме того, Bcl-6 является фактором транскрипции, идентифицированным в клетках T FH , но его роль может выходить за рамки этого подмножества, поскольку он также участвует в CD8 памяти. + Развитие Т-клеток. [10]
В зародышевых центрах клетки Т - ФГ , подвергшиеся воздействию антигена , быстро повышают экспрессию CD40L, который связывает и стимулирует поверхностный рецептор В-клеток CD40. [11] Зависимая от Т -ФГ -клеток паракринная активация CD40 В-клеток приводит к выживанию и дифференцировке В-клеток, включая индукцию AID ( индуцированной активацией (цитидин) деаминазы ). [12] Экспрессия AID (кодируемая геном AICDA ) вызывает переключение класса В-клеточных антител с IgM/IgD на другие изотипы антител и вызывает соматическую гипермутацию во время клональной пролиферации. Переключенные антитела приобретают лучшие эффекторные функции, а гипермутированные антитела проявляют большее сродство к антигену.
Классы Т- ФГ- клеток
[ редактировать ]Клетки Т -СГ, образующиеся на ранних стадиях реакции зародышевого центра, формально называются клетками пре-Т -СГ . Они обнаруживаются преимущественно на границе Т-клеточной зоны, которая сливается с В-клеточными фолликулами и зародышевыми центрами. Клетки Pre-T FH функционально очень похожи на другие клетки T FH в облегчении реакций В-клеточных центров зародышевого центра; однако они также способны стимулировать развитие фолликулярных В-клеток, прилегающих к зародышевым центрам и за их пределами, вызывая быстро реагирующие, но недолговечные реакции антител, управляемые плазматическими клетками (известные как экстрафолликулярный ответ).
Эти клетки T FH, специфически находящиеся в зрелом зародышевом центре, иногда называют клетками GC T FH зародышевого центра (для клеток T FH ), чтобы отличить их от клеток пре-T FH . [13] [14] В этой популяции клеток GC Tfh имеется также второстепенный подкласс, который экспрессирует ген Foxp3 , кодирующий фактор транскрипции. Эта небольшая дискретная субпопуляция клеток, называемая Т- FR- клетками (т.е. Т-фолликулярными регуляторными клетками), играет важную роль в контроле и ограничении величины нормальных реакций зародышевого центра, чтобы избежать возможности образования аномально мутированных или аутореактивных клеток. аутоиммунно -ассоциированные антитела. [15] Следовательно, клетки T FR оказывают уникальное ингибирующее влияние на реакцию зародышевого центра.
Хотя Т -ФГ -клетки обнаруживаются главным образом во вторичных лимфоидных органах, небольшая их часть циркулирует в крови и называется «периферическими» Т-фолликулярными хелперными клетками (pT FH ). Эти клетки можно идентифицировать по экспрессии ими IL-21 при стимуляции. [16]
Медицинская значимость
[ редактировать ]Создание прочной иммунной памяти
[ редактировать ]Т- СГ- клетки считаются незаменимой субпопуляцией Т-клеток в генерации и поддержании ответов зародышевого центра . Следовательно, в отсутствие Т -ФГ- клеток, подобно активации В-клеток независимыми антигенами Т-клеток, формируется быстрый всплеск продукции низкоаффинных плазматических клеток, но это не приводит к индукции зародышевого центра и не способствует созреванию аффинности антител или дифференцировке. эффективных В-клеток памяти , которые необходимы для защиты организма от последующих инфекций. В частности, В-клетки памяти, зависимые от зародышевого центра, являются движущей силой продукции отзывных антител во время вторичного иммунного ответа. Следовательно, правильная активация и развитие клеток Т -ФГ имеет решающее значение для эффективности иммунизации и разработки вакцин для индукции долговременного иммунитета . В бангладешском популяционном исследовании пациентов, инфицированных холерным вибрионом , и здоровых добровольцев, которым вводили существующую вакцину против холеры, [17] памяти, ответ Т- ФГ специфичный против холерного антигена, коррелировал с дальнейшей продукцией антител В-клетками.
Контроль возрастного снижения иммунитета
[ редактировать ]При нормальном старении происходит постепенное ослабление иммунной системы организма. Этот феномен, называемый иммуностарением , во многом обусловлен снижением функции Т-клеток, включая способность Т- ФГ- клеток должным образом поддерживать реакции зародышевого центра . [18] Частично это может быть связано с более низкими уровнями CD40L на поверхности клеток T FH у пожилых людей. [19]
Как избежать аутоиммунитета
[ редактировать ]Неконтролируемые или сверхактивные иммунные реакции Т -ФГ- клеток могут привести к образованию необоснованных зародышевых центров, состоящих из аберрантно мутировавших В-клеток опосредованные антителами , которые могут вызывать аутоиммунные заболевания, . Повышенные уровни Т- ФГ- подобных клеток могут быть обнаружены в крови пациентов с системной красной волчанкой (СКВ) и синдромом Шегрена . [20] Однако научные данные, свидетельствующие о том, что Т- ФГ- клетки могут окончательно вызывать аутоиммунитет у людей, остаются неполными.
Ссылки
[ редактировать ]- ^ Фазильо; Марк, Л; Макхейзер-Уильямс, LJ; Макхейзер-Уильямс, МГ; и др. (март 2009 г.). «Фолликулярные Т-хелперы: происхождение и расположение» . Иммунитет . 30 (3): 324–35. doi : 10.1016/j.immuni.2009.03.003 . ПМК 2731675 . ПМИД 19303387 .
- ^ Сео; Юн, Дж; Ким, PH; и др. (май 2009 г.). «IL-21 обеспечивает индуцированную TGF-бета 1 экспрессию изотипа IgA в пейеровых пятнах мыши». J Лейкок Биол . 85 (5): 744–50. дои : 10.1189/jlb.0708450 . ПМИД 19168593 . S2CID 22358429 .
- ^ Зарецкий; Тейлор, Джей-Джей; Кинг, Иллинойс; Маршалл, ФА; Морс, М; Пирс, Э.Дж.; и др. (май 2009 г.). «Т-фолликулярные хелперные клетки дифференцируются из клеток Th2 в ответ на антигены гельминтов» . Джей Эксп Мед . 206 (5): 991–9. дои : 10.1084/jem.20090303 . ПМК 2715032 . ПМИД 19380637 .
- ^ Куросаки; Кометани, К; Исе, W (март 2015 г.). «В-клетки памяти». Нат Рев Иммунол . 15 (3): 149–59. дои : 10.1038/nri3802 . ПМИД 25677494 . S2CID 20825732 .
- ^ Чтанова; Танье, Южная Дакота; Ньютон, Р; Франк, Н; Ходж, MR; Рольф, MS; Маккей, ЧР; и др. (июль 2004 г.). «Т-фолликулярные хелперные клетки имеют характерный транскрипционный профиль, отражающий их роль в качестве эффекторных клеток, не относящихся к Th1/Th2, которые оказывают помощь В-клеткам» . Дж Иммунол . 173 (1): 68–78. дои : 10.4049/jimmunol.173.1.68 . ПМИД 15210760 .
- ^ Нуриева Р.И., Чунг Ю., Хван Д., Ян Ксо, Кан Х.С., Ма Л., Ван Ю.Х., Ватович С.С. и др. (2008). «Поколение Т-фолликулярных хелперных клеток опосредовано интерлейкином-21, но независимо от клеточных линий Т-хелперов 1, 2 или 17» . Иммунитет . 29 (1): 138–49. doi : 10.1016/j.immuni.2008.05.009 . ПМЦ 2556461 . ПМИД 18599325 .
- ^ Акиба; Такеда, К.; Кодзима, Ю; Усуи, Ю; Харада, Н.; Ямадзаки, Т; Ма, Дж; Тезука, К; и др. (август 2005 г.). «Роль ICOS в поддержании фолликулярных В-хелперных Т-клеток CXCR5+ in vivo» . Дж Иммунол . 175 (4): 2340–8. дои : 10.4049/гиммунол.175.4.2340 . ПМИД 16081804 .
- ^ Буке; Джин, Х; Патерсон, AM; Митсдорффер, М; Хо, IC; Шарп, АХ; Качроо, В.К. (февраль 2009 г.). «Костимулирующая молекула ICOS регулирует экспрессию c-Maf и IL-21 при развитии фолликулярных Т-хелперных клеток и клеток TH-17» . Нат Иммунол . 10 (2): 167–75. дои : 10.1038/ni.1690 . ПМЦ 2742982 . ПМИД 19098919 .
- ^ Фогельзанг; Макгуайр, HM; Ю, Д; Срент, Дж; Маккей, ЧР; Кинг, К. (июль 2008 г.). «Фундаментальная роль интерлейкина-21 в образовании Т-фолликулярных хелперных клеток» . Иммунитет . 29 (1): 127–37. doi : 10.1016/j.immuni.2008.06.001 . ПМИД 18602282 .
- ^ Ичии; Сакамото, А; Хатано, М; Окада, С; Тояма, Х; Таки, С; Арима, М; Курода, Ю; Токухиса, Т; и др. (июнь 2002 г.). «Роль Bcl-6 в создании и поддержании CD8+ Т-клеток памяти». Нат Иммунол . 3 (6): 558–63. дои : 10.1038/ni802 . hdl : 20.500.14094/D1002645 . ПМИД 12021781 . S2CID 2552376 .
- ^ Ким, младший; Лим, Х.В.; Канг, СГ; Хиллсамер, П; Ким, Швейцария (4 февраля 2005 г.). «Т-клетки зародышевого центра CD57+ человека являются основными помощниками для клеток GC-B и вызывают рекомбинацию переключения класса» . БМК Иммунология . 6 :3. дои : 10.1186/1471-2172-6-3 . ПМЦ 548684 . ПМИД 15694005 .
- ^ Макхейзер-Уильямс, М; Окицу, С; Ван, Н.; Макхейзер-Уильямс, L (9 декабря 2011 г.). «Молекулярное программирование памяти В-клеток» . Обзоры природы. Иммунология . 12 (1): 24–34. дои : 10.1038/nri3128 . ПМЦ 3947622 . ПМИД 22158414 .
- ^ Юсуф И., Кагеяма Р., Монтичелли Л., Джонстон Р.Дж., Диторо Д., Хансен К., Барнетт Б., Кротти С. (июль 2010 г.). «Выработка IL-4 фолликулярными хелперными клетками зародышевого центра зависит от рецептора сигнальной молекулы активации лимфоцитов (CD150)» . Дж. Иммунол . 185 (1): 190–202. doi : 10.4049/jimmunol.0903505 . ПМЦ 2913439 . ПМИД 20525889 .
- ^ Рамискаль Р.Р., Винуэса К.Г. (март 2013 г.). «Субпопуляции Т-клеток в зародышевом центре». Иммунол. Преподобный . 252 (1): 146–55. дои : 10.1111/imr.12031 . ПМИД 23405902 . S2CID 21787025 .
- ^ Волленберг И., Агуа-Досе А., Эрнандес А., Алмейда С., Оливейра В.Г., Фару Х., Граса Л. (ноябрь 2011 г.). «Регуляция реакции зародышевого центра с помощью фолликулярных регуляторных Т-клеток Foxp3» . Дж. Иммунол . 187 (9): 4553–60. doi : 10.4049/jimmunol.1101328 . ПМИД 21984700 .
- ^ Шульц, Брюс Т.; Тейглер, Джеффри Э.; Писсани, Франко; Остер, Александр Ф.; Краниас, Грегори; Альтер, Галит ; Марович, Мария; Эллер, Майкл А.; Диттмер, Ульф (19 января 2016 г.). «Циркулирующие ВИЧ-специфические Т-клетки интерлейкина-21(+)CD4(+) представляют собой периферические клетки Tfh с антиген-зависимыми хелперными функциями» . Иммунитет . 44 (1): 167–178. doi : 10.1016/j.immuni.2015.12.011 . ISSN 1097-4180 . ПМИД 26795249 .
- ^ Арифуззаман М., Рашу Р., Люнг Д.Т., Хосен М.И., Бхуян Т.Р., Бхуян М.С., Рахман М.А., Ханам Ф., Саха А., Чарльз Р.К., ЛаРок Р.К., Вейл А.А., Клементс Дж.Д., Холмс Р.К., Колдервуд С.Б., Харрис Дж.Б., Райан Э.Т. , Кадри Ф (август 2012 г.). «Антигенспецифические реакции Т-клеток памяти после вакцинации пероральной убитой холерной вакциной у детей Бангали и сравнение с реакциями у пациентов с холерой, приобретенной естественным путем» . Клиническая и вакциноиммунология . 19 (8): 1304–11. дои : 10.1128/CVI.00196-12 . ПМК 3416086 . ПМИД 22739692 .
- ^ Лефевр Дж.С., Мауэ А.С., Итон С.М., Лантье П.А., Тайг М., Хейнс Л. (2012). «Старое микроокружение способствует возрастным функциональным дефектам CD4 Т-клеток у мышей» . Стареющая клетка . 11 (5): 732–40. дои : 10.1111/j.1474-9726.2012.00836.x . ПМЦ 3444657 . ПМИД 22607653 .
- ^ Перки Э., Миллер Р.А., Гарсия Г.Г. (2012). «Ферментативная обработка старых CD4 Т-клеток ex vivo восстанавливает хелперную функцию родственных Т-клеток и усиливает выработку антител у мышей» . Дж Иммунол . 189 (12): 5582–9. doi : 10.4049/jimmunol.1200487 . ПМЦ 3518580 . ПМИД 23136198 .
- ^ Крафт JE. (2012). «Фолликулярные Т-хелперы в иммунитете и системном аутоиммунитете» . Нат Рев Ревматол . 8 (6): 337–47. дои : 10.1038/nrheum.2012.58 . ПМК 3604997 . ПМИД 22549246 .