В-клетка
| В-лимфоцит клетка | |
|---|---|
 Анимация B-клетки | |
| Подробности | |
| Предшественник | Гемопоэтические стволовые клетки |
| Система | Иммунная система |
| Идентификаторы | |
| латинский | лимфоцит Б |
| МеШ | D001402 |
| ФМА | 62869 |
| Анатомические термины микроанатомии | |
клетки , также известные как В-лимфоциты , представляют собой разновидность лейкоцитов подтипа лимфоцитов В - . [1] Они функционируют в иммунитета гуморальном компоненте адаптивной иммунной системы . [1] В-клетки производят молекулы антител , которые могут либо секретироваться, либо встраиваться в плазматическую мембрану, где они служат частью рецепторов В-клеток . [2] Когда наивная В-клетка или В-клетка памяти активируется антигеном, она пролиферирует и дифференцируется в эффекторную клетку, секретирующую антитела, известную как плазмобласт или плазматическая клетка. [2] Кроме того, В-клетки презентируют антигены (их также классифицируют как профессиональные антигенпрезентирующие клетки, АПК ) и секретируют цитокины . [1] У млекопитающих , в том числе сумчатых [3] В-клетки созревают в костном мозге , который является основой большинства костей . [4] У птиц В-клетки созревают в сумке Фабрициуса , лимфоидном органе, где они были впервые обнаружены Чангом и Гликом. [5] вот почему буква B означает бурсу , а не костный мозг , как принято считать.
В-клетки, в отличие от двух других классов лимфоцитов, Т-клеток и естественных клеток-киллеров , экспрессируют рецепторы В-клеток (BCR) на своей клеточной мембране . [1] BCR позволяют В-клетке связываться с чужеродным антигеном , против которого она инициирует ответ антител. [1] Рецепторы В-клеток чрезвычайно специфичны: все BCR на В-клетке распознают один и тот же эпитоп . [6]
Разработка
[ редактировать ]
В-клетки развиваются из гемопоэтических стволовых клеток (ГСК), происходящих из костного мозга . [7] [8] ЗКП сначала дифференцируются в мультипотентные клетки-предшественники (MPP), а затем в общие лимфоидные клетки-предшественники (CLP). [8] Отсюда их развитие в B-клетки происходит в несколько стадий (показано на изображении справа), каждая из которых отмечена различными паттернами экспрессии генов и иммуноглобулина H-цепи и L-цепи расположением генных локусов , последнее связано с тем, что B-клетки подвергаются V (D) J-рекомбинации по мере их развития. [9]
В-клетки подвергаются двум типам отбора во время развития в костном мозге, чтобы обеспечить правильное развитие, оба с участием рецепторов В-клеток (BCR) на поверхности клетки. Положительный отбор происходит посредством антиген-независимой передачи сигналов с участием как pre-BCR, так и BCR. [10] [11] Если эти рецепторы не связываются со своим лигандом , В-клетки не получают нужных сигналов и перестают развиваться. [10] [11] Негативная селекция происходит посредством связывания аутоантигена с BCR; если BCR может прочно связываться с аутоантигеном, то B-клетка претерпевает одну из четырех судеб: клональную делецию , редактирование рецептора , анергию или игнорирование (B-клетка игнорирует сигнал и продолжает развитие). [11] Этот процесс негативного отбора приводит к состоянию центральной толерантности , при котором зрелые В-клетки не связывают аутоантигены, присутствующие в костном мозге. [9]
Для завершения развития незрелые В-клетки мигрируют из костного мозга в селезенку в виде переходных В-клеток , проходя две переходные стадии: Т1 и Т2. [12] На протяжении всей миграции в селезенку и после попадания в селезенку они считаются В-клетками Т1. [13] В селезенке В-клетки Т1 переходят в В-клетки Т2. [13] В-клетки Т2 дифференцируются либо в В-клетки фолликула (FO), либо в В-клетки маргинальной зоны (MZ) в зависимости от сигналов, полученных через BCR и другие рецепторы. [14] После дифференцировки они теперь считаются зрелыми В-клетками или наивными В-клетками. [13]
Активация
[ редактировать ]
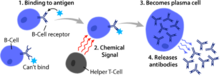
Активация В-клеток происходит во вторичных лимфоидных органах (SLO), таких как селезенка и лимфатические узлы . [1] После того, как В-клетки созревают в костном мозге, они мигрируют через кровь в SLO, которые получают постоянный запас антигена через циркулирующую лимфу . [15] В SLO активация В-клеток начинается, когда В-клетка связывается с антигеном через свой BCR. [16] Хотя события, происходящие сразу после активации, еще полностью не определены, считается, что В-клетки активируются в соответствии с моделью кинетической сегрегации. [ нужна ссылка ] , первоначально определяемый в Т-лимфоцитах. Эта модель означает, что перед стимуляцией антигеном рецепторы диффундируют через мембрану, вступая в контакт с Lck и CD45 с одинаковой частотой, обеспечивая чистое равновесие фосфорилирования и нефосфорилирования. Только когда клетка вступает в контакт с антигенпрезентирующей клеткой, более крупный CD45 смещается из-за близкого расстояния между двумя мембранами. Это позволяет осуществить чистое фосфорилирование BCR и инициировать путь передачи сигнала. [ нужна ссылка ] . Из трех подпопуляций В-клеток В-клетки FO преимущественно подвергаются активации, зависящей от Т-клеток, тогда как В-клетки MZ и В-клетки B1 преимущественно подвергаются активации, независимой от Т-клеток. [17]
Активация В-клеток усиливается за счет активности CD21 , поверхностного рецептора в комплексе с поверхностными белками CD19 и CD81 (все три вместе известны как корецепторный комплекс В-клеток). [18] Когда BCR связывает антиген, меченный фрагментом белка комплемента C3, CD21 связывает фрагмент C3, соединяется со связанным BCR, и сигналы передаются через CD19 и CD81, чтобы снизить порог активации клетки. [19]
Т-клеточно-зависимая активация
[ редактировать ]Антигены, которые активируют В-клетки с помощью Т-клеток, известны как Т-клеточно-зависимые (TD) антигены и включают чужеродные белки. [1] Они названы так потому, что не способны вызывать гуморальный ответ у организмов, лишенных Т-клеток. [1] Реакция В-клеток на эти антигены занимает несколько дней, хотя образующиеся антитела обладают более высоким сродством и более функционально универсальны, чем антитела, образующиеся в результате независимой от Т-клеток активации. [1]
Как только BCR связывает антиген TD, антиген попадает в В-клетку посредством рецептор-опосредованного эндоцитоза , разлагается и презентируется Т-клеткам в виде пептидных фрагментов в комплексе с молекулами MHC-II на клеточной мембране. [20] Т-хелперы ( TH ) , обычно фолликулярные Т-хелперы (TFH ) , распознают и связывают эти комплексы MHC-II-пептид через свой Т-клеточный рецептор (TCR) . [21] После связывания пептида TCR-MHC-II Т-клетки экспрессируют поверхностный белок CD40L , а также цитокины, такие как IL-4 и IL-21 . [21] CD40L служит необходимым костимулирующим фактором для активации В-клеток путем связывания поверхностного рецептора В-клеток CD40 , который способствует пролиферации В-клеток , переключению классов иммуноглобулинов и соматической гипермутации , а также поддерживает рост и дифференцировку Т-клеток. [1] Цитокины Т-клеточного происхождения, связанные с рецепторами цитокинов В-клеток , также способствуют пролиферации В-клеток, переключению классов иммуноглобулинов и соматической гипермутации, а также направляют дифференцировку. [21] После того как В-клетки получают эти сигналы, они считаются активированными. [21]

После активации В-клетки участвуют в двухэтапном процессе дифференцировки, в результате которого образуются как короткоживущие плазмобласты для немедленной защиты, так и долгоживущие плазматические клетки и В-клетки памяти для постоянной защиты. [17] Первый этап, известный как экстрафолликулярный ответ, происходит за пределами лимфоидных фолликулов, но все еще в SLO. [17] На этом этапе активированные В-клетки пролиферируют, могут подвергаться переключению класса иммуноглобулинов и дифференцироваться в плазмобласты, которые продуцируют ранние слабые антитела, в основном класса IgM. [22]

Второй этап состоит из активированных В-клеток, проникающих в лимфоидный фолликул и образующих зародышевый центр (ГЦ) , который представляет собой специализированную микросреду, где В-клетки подвергаются обширной пролиферации, переключению класса иммуноглобулина и созреванию аффинности, направляемому соматической гипермутацией. [23] Этим процессам способствуют Т -ФГ- клетки внутри ГК и генерируют как В-клетки памяти с высоким сродством, так и долгоживущие плазматические клетки. [17] Образующиеся плазматические клетки секретируют большое количество антител и либо остаются в пределах SLO, либо, что более предпочтительно, мигрируют в костный мозг. [23]
Независимая от Т-клеток активация
[ редактировать ]Антигены, которые активируют В-клетки без помощи Т-клеток, известны как Т-клеточно-независимые (TI) антигены. [1] и включают чужеродные полисахариды и неметилированную CpG-ДНК. [17] Они названы так потому, что способны вызывать гуморальный ответ у организмов, лишенных Т-клеток. [1] Ответ В-клеток на эти антигены быстрый, хотя образующиеся антитела имеют тенденцию иметь более низкое сродство и менее функционально универсальны, чем антитела, образующиеся в результате Т-клеточно-зависимой активации. [1]
Как и в случае с TD-антигенами, В-клетки, активированные TI-антигенами, нуждаются в дополнительных сигналах для завершения активации, но вместо получения их от Т-клеток они обеспечиваются либо распознаванием и связыванием общего микробного компонента с toll-подобными рецепторами (TLR), либо путем обширное сшивание BCR с повторяющимися эпитопами бактериальной клетки. [1] В-клетки, активированные антигенами TI, продолжают пролиферировать за пределами лимфоидных фолликулов, но все еще находятся в SLO (GC не образуются), возможно, подвергаются переключению класса иммуноглобулинов и дифференцируются в короткоживущие плазмобласты, которые продуцируют ранние, слабые антитела, в основном класса IgM, но также некоторые популяции долгоживущих плазматических клеток. [24]
Активация B-клеток памяти
[ редактировать ]Активация В-клеток памяти начинается с обнаружения и связывания их целевого антигена, который является общим для их родительской В-клетки. [25] Некоторые В-клетки памяти могут быть активированы без помощи Т-клеток, например, некоторые вирус-специфические В-клетки памяти, но другим требуется помощь Т-клеток. [26] При связывании антигена В-клетка памяти поглощает антиген посредством рецептор-опосредованного эндоцитоза, расщепляет его и представляет Т-клеткам в виде фрагментов пептида в комплексе с молекулами MHC-II на клеточной мембране. [25] Т-хелперы памяти ( TH ), обычно фолликулярные Т-хелперы памяти (TFH ) , которые были получены из Т-клеток, активированных тем же антигеном, распознают и связывают эти комплексы MHC-II-пептид через их TCR. [25] После связывания пептида TCR-MHC-II и передачи других сигналов от клетки памяти Т- ФГ В-клетка памяти активируется и дифференцируется либо в плазмобласты и плазматические клетки посредством внефолликулярного ответа, либо вступает в реакцию зародышевого центра, где они генерируют плазму. клеток и большего количества В-клеток памяти. [25] [26] Неясно, подвергаются ли В-клетки памяти дальнейшему созреванию аффинности внутри этих вторичных GC. [25] in vitro Активации B-клеток памяти против CD40 можно добиться путем стимуляции различными активаторами, такими как митоген рябины или моноклональные антитела , однако исследование показало, что комбинация R-848 и рекомбинантного человеческого IL-2 является наиболее эффективным активатором. . [27]
Типы В-клеток
[ редактировать ]
- Плазмобласт
- Короткоживущая пролиферирующая клетка, секретирующая антитела, возникающая в результате дифференцировки В-клеток. [1] Плазмобласты образуются на ранних стадиях инфекции, и их антитела, как правило, имеют более слабое сродство к целевому антигену по сравнению с плазматическими клетками. [17] Плазмобласты могут возникать в результате независимой от Т-клеток активации В-клеток или экстрафолликулярного ответа в результате Т-зависимой активации В-клеток. [1]
- Плазматическая клетка
- Долгоживущая, непролиферирующая клетка, секретирующая антитела, возникающая в результате дифференцировки В-клеток. [1] Имеются данные о том, что В-клетки сначала дифференцируются в плазмобластоподобные клетки, а затем дифференцируются в плазматические клетки. [17] Плазматические клетки образуются позже при инфекции и, по сравнению с плазмобластами, имеют антитела с более высоким сродством к целевому антигену из-за созревания сродства в зародышевом центре (ГЦ) и производят больше антител. [17] Плазматические клетки обычно возникают в результате реакции зародышевого центра в результате Т-клеточно-зависимой активации В-клеток, хотя они также могут возникать в результате независимой от Т-клеток активации В-клеток. [24]
- Лимфоплазмоцитоидная клетка
- Клетка со смесью морфологических особенностей В-лимфоцитов и плазматических клеток, которая считается близкородственной или подтипом плазматических клеток. Этот тип клеток обнаруживается при предраковых и злокачественных дискразиях плазматических клеток , которые связаны с секрецией IgM моноклональных белков ; эти дискразии включают моноклональную гаммапатию IgM неопределенного значения и макроглобулинемию Вальденстрема . [28]
- Ячейка памяти B
- Спящие В-клетки, возникающие в результате дифференцировки В-клеток. [1] Их функция состоит в том, чтобы циркулировать по организму и инициировать более сильный и быстрый ответ антител (известный как анамнестический вторичный ответ антител), если они обнаруживают антиген, который активировал их родительскую В-клетку (В-клетки памяти и их родительские В-клетки имеют одинаковые BCR, таким образом, они обнаруживают один и тот же антиген). [26] В-клетки памяти могут генерироваться в результате Т-клеточно-зависимой активации посредством как экстрафолликулярного ответа, так и реакции зародышевого центра, а также в результате независимой от Т-клеток активации В1-клеток. [26]
- Ячейка Б-2
- FO B-клетки и MZ B-клетки. [29]
- Фолликулярная (FO) B-клетка (также известная как B-2-клетка)
- Наиболее распространенный тип В-клеток, когда они не циркулируют в крови, обнаруживается в основном в лимфоидных фолликулах вторичных лимфоидных органов (SLO). [17] Они ответственны за выработку большинства высокоаффинных антител во время инфекции. [1]
- В-клетка маргинальной зоны (MZ)
- Встречается главным образом в маргинальной зоне селезенки и служит первой линией защиты от передающихся через кровь патогенов, поскольку маргинальная зона получает большое количество крови из общего кровообращения. [30] Они могут подвергаться как независимой от Т-клеток, так и зависимой от Т-клеток активации, но предпочтительно подвергаться независимой от Т-клеток активации. [17]
- В-1 ячейка
- Возникает на пути развития, отличном от В-клеток FO и B-клеток MZ. [29] У мышей они преимущественно заселяют брюшную полость и плевральную полость , генерируют естественные антитела (антитела, вырабатываемые без инфекции), защищают от патогенов слизистой оболочки и в первую очередь проявляют независимую от Т-клеток активацию. [29] Истинный гомолог мышиных клеток B-1 у человека не обнаружен, хотя описаны различные популяции клеток, сходные с клетками B-1. [29]
- Регуляторная B-клетка (Breg)
- Иммуносупрессивный тип В - клеток, который останавливает размножение патогенных провоспалительных лимфоцитов посредством секреции IL-10, IL-35 и TGF-β. [31] Кроме того, он способствует образованию регуляторных Т-клеток (Treg) , напрямую взаимодействуя с Т-клетками, искажая их дифференцировку в сторону Treg. [31] Никакой общей идентичности клеток Breg не описано, и как у мышей, так и у людей было обнаружено множество подпопуляций клеток Breg, разделяющих регуляторные функции. [31] В настоящее время неизвестно, связаны ли подмножества клеток Breg в процессе развития и как именно происходит дифференцировка в клетки Breg. [31] Имеются данные, показывающие, что почти все типы В-клеток могут дифференцироваться в клетки Breg посредством механизмов, включающих воспалительные сигналы и распознавание BCR. [31]
Патология, связанная с B-клетками
[ редактировать ]Аутоиммунное заболевание может возникнуть в результате аномального распознавания аутоантигенов В-клетками с последующей выработкой аутоантител. [32] Аутоиммунные заболевания, при которых активность заболевания коррелирует с активностью В-клеток, включают склеродермию , рассеянный склероз , системную красную волчанку , диабет 1 типа , постинфекционный синдром раздраженного кишечника и ревматоидный артрит . [32]
Злокачественная трансформация В-клеток и их предшественников может вызывать множество видов рака , включая хронический лимфоцитарный лейкоз (ХЛЛ) , острый лимфобластный лейкоз (ОЛЛ) , волосатоклеточный лейкоз , фолликулярную лимфому , неходжкинскую лимфому , лимфому Ходжкина и злокачественные плазмоклеточные опухоли , такие как как множественная миелома , макроглобулинемия Вальденстрема и некоторые формы амилоидоза . [33] [34]
Аномальные В-клетки могут быть относительно большими, и некоторые заболевания включают это в свои названия, например, диффузная крупноклеточная В-клеточная лимфома (DLBCL) и внутрисосудистая крупноклеточная В-клеточная лимфома .
Пациенты с В-клеточным лимфоцитозом предрасположены к инфекциям. [35]
Эпигенетика
[ редактировать ]Исследование, в котором изучали метилом В-клеток на протяжении их цикла дифференцировки с использованием полногеномного бисульфитного секвенирования (WGBS), показало, что существует гипометилирование от самых ранних стадий до наиболее дифференцированных стадий. Наибольшая разница в метилировании наблюдается между стадиями В-клеток зародышевого центра и В-клеток памяти. Кроме того, это исследование показало, что существует сходство между опухолями В-клеток и долгоживущими В-клетками в признаках метилирования их ДНК . [36]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Мерфи К. (2012). Иммунобиология Джейнвей (8-е изд.). Нью-Йорк: Garland Science. ISBN 9780815342434 .
- ^ Перейти обратно: а б Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «В-клетки и антитела» . Молекулярная биология клетки (4-е изд.). Гирляндная наука.
- ^ Шравен, Андреа Л.; Хансен, Виктория Л.; Моррисси, Калифорния; Стэннард, Хейли Дж.; Онг, Оселин OTW; Дуек, округ Колумбия; Миллер, Роберт Д.; Олд, Джули М. (2021). «Анализ одноклеточного транскриптома репертуара B-клеток показывает использование иммуноглобулинов у серого короткохвостого опоссума ( Monodelphis Domestica )». Развивающая и сравнительная иммунология . 123 : 104141. doi : 10.1016/j.dci.2021.104141 .
- ^ Купер, доктор медицинских наук (март 2015 г.). «Ранняя история В-клеток» . Обзоры природы. Иммунология . 15 (3): 191–197. дои : 10.1038/nri3801 . ПМИД 25656707 .
- ^ Глик, Брюс; Чанг, Тимоти С.; Яап, Р. Джордж (1 января 1956 г.). «Бурса Фабрициуса и производство антител» . Птицеводство . 35 (1): 224–225. дои : 10.3382/ps.0350224 . ISSN 0032-5791 .
- ^ Джесперсен, Мартин Клостер; Махаджан, Свапнил; Петерс, Бьорн; Нильсен, Мортен; Маркатили, Паоло (2019). «Прогнозирование антитело-специфического эпитопа B-клеток: использование информации из белковых комплексов антитело-антиген» . Границы в иммунологии . 10 :298. дои : 10.3389/fimmu.2019.00298 . ПМК 6399414 . ПМИД 30863406 .
- ^ Фишер Ю, Ян Дж. Дж., Икава Т., Хейн Д., Висенте-Дуэньяс К., Боркхардт А., Санчес-Гарсия I (ноябрь 2020 г.). «Решения клеточной судьбы: роль факторов транскрипции в раннем развитии B-клеток и лейкемии» . Открытие рака крови . 1 (3): 224–233. дои : 10.1158/2643-3230.BCD-20-0011 . ПМЦ 7774874 . ПМИД 33392513 .
- ^ Перейти обратно: а б Кондо М (ноябрь 2010 г.). «Приверженность лимфоидной и миелоидной линии у мультипотентных гемопоэтических предшественников» . Иммунологические обзоры . 238 (1): 37–46. дои : 10.1111/j.1600-065X.2010.00963.x . ПМЦ 2975965 . ПМИД 20969583 .
- ^ Перейти обратно: а б Пеланда Р., Торрес Р.М. (апрель 2012 г.). «Центральная толерантность B-клеток: где начинается отбор» . Перспективы Колд-Спринг-Харбор в биологии . 4 (4): а007146. doi : 10.1101/cshperspect.a007146 . ПМК 3312675 . ПМИД 22378602 .
- ^ Перейти обратно: а б Мортенссон И.Л., Альмквист Н., Гримсхольм О., Бернарди А.И. (июнь 2010 г.). «Контрольная точка рецептора пре-В-клеток» . Письма ФЭБС . 584 (12): 2572–2579. дои : 10.1016/j.febslet.2010.04.057 . ПМИД 20420836 . S2CID 43158480 .
- ^ Перейти обратно: а б с ЛеБьен Т.В., Теддер Т.Ф. (сентябрь 2008 г.). «В-лимфоциты: как они развиваются и функционируют» . Кровь . 112 (5): 1570–1580. doi : 10.1182/blood-2008-02-078071 . ПМК 2518873 . ПМИД 18725575 .
- ^ Лодер Ф., Мучлер Б., Рэй Р.Дж., Пейдж С.Дж., Сидерас П., Торрес Р. и др. (июль 1999 г.). «Развитие В-клеток в селезенке происходит дискретными этапами и определяется качеством сигналов, поступающих от рецепторов В-клеток» . Журнал экспериментальной медицины . 190 (1): 75–89. дои : 10.1084/jem.190.1.75 . ПМК 2195560 . ПМИД 10429672 .
- ^ Перейти обратно: а б с Чунг Дж.Б., Сильверман М., Монро Дж.Г. (июнь 2003 г.). «Переходные B-клетки: шаг за шагом к иммунной компетентности». Тенденции в иммунологии . 24 (6): 343–349. дои : 10.1016/S1471-4906(03)00119-4 . ПМИД 12810111 .
- ^ Черутти А, Колс М, Пуга I (февраль 2013 г.). «В-клетки маргинальной зоны: достоинства лимфоцитов, продуцирующих врожденные антитела» . Обзоры природы. Иммунология . 13 (2): 118–132. дои : 10.1038/nri3383 . ПМЦ 3652659 . ПМИД 23348416 .
- ^ Харвуд, штат Небраска, Батиста, федеральный округ (01 января 2010 г.). «Ранние события активации B-клеток». Ежегодный обзор иммунологии . 28 (1): 185–210. doi : 10.1146/annurev-immunol-030409-101216 . ПМИД 20192804 .
- ^ Юсефф М.И., Пьеробон П., Реверсат А., Леннон-Дюмениль А.М. (июль 2013 г.). «Как В-клетки захватывают, обрабатывают и представляют антигены: решающая роль в полярности клеток». Обзоры природы. Иммунология . 13 (7): 475–486. дои : 10.1038/nri3469 . ПМИД 23797063 . S2CID 24791216 .
- ^ Перейти обратно: а б с д и ж г час я дж Натт С.Л., Ходжкин П.Д., Тарлинтон Д.М., Коркоран Л.М. (март 2015 г.). «Поколение плазматических клеток, секретирующих антитела». Обзоры природы. Иммунология . 15 (3): 160–171. дои : 10.1038/nri3795 . ПМИД 25698678 . S2CID 9769697 .
- ^ Асокан Р., Банда Н.К., Сакони Г., Чен X.С., Холерс В.М. (январь 2013 г.). «Рецептор 2 человеческого комплемента (CR2/CD21) как рецептор ДНК: значение его роли в иммунном ответе и патогенезе системной красной волчанки (СКВ)» . Молекулярная иммунология . 53 (1–2): 99–110. дои : 10.1016/j.molimm.2012.07.002 . ПМЦ 3439536 . ПМИД 22885687 .
- ^ Забель, доктор медицинских наук, Вейс Дж. Х. (март 2001 г.). «Клеточно-специфическая регуляция гена CD21». Международная иммунофармакология . Раскрытие механизмов и обнаружение новых ролей дополнения. 1 (3): 483–493. дои : 10.1016/S1567-5769(00)00046-1 . ПМИД 11367532 .
- ^ Блюм Дж.С., Уирш П.А., Крессвелл П. (1 января 2013 г.). «Пути процессинга антигена» . Ежегодный обзор иммунологии . 31 (1): 443–473. doi : 10.1146/annurev-immunol-032712-095910 . ПМК 4026165 . ПМИД 23298205 .
- ^ Перейти обратно: а б с д Кротти С. (март 2015 г.). «Краткая история помощи Т-клеток В-клеткам» . Обзоры природы. Иммунология . 15 (3): 185–189. дои : 10.1038/nri3803 . ПМЦ 4414089 . ПМИД 25677493 .
- ^ МакЛеннан И.С., Теллнер К.М., Каннингем А.Ф., Серр К., Сзе Д.М., Суньига Э. и др. (август 2003 г.). «Экстрафолликулярные антитела». Иммунологические обзоры . 194 : 8–18. дои : 10.1034/j.1600-065x.2003.00058.x . ПМИД 12846803 . S2CID 2455541 .
- ^ Перейти обратно: а б Шломчик М.Ю., Вайзель Ф. (май 2012 г.). «Выбор зародышевого центра и развитие памяти B и плазматических клеток» . Иммунологические обзоры . 247 (1): 52–63. дои : 10.1111/j.1600-065X.2012.01124.x . ПМИД 22500831 . S2CID 5362003 .
- ^ Перейти обратно: а б Бортник А., Чернова И., Куинн В.Дж., Мюнье М., Канкро М.П., Оллман Д. (июнь 2012 г.). «Долгоживущие плазматические клетки костного мозга индуцируются на ранней стадии в ответ на независимые или зависимые от Т-клеток антигены» . Журнал иммунологии . 188 (11): 5389–5396. doi : 10.4049/jimmunol.1102808 . ПМК 4341991 . ПМИД 22529295 .
- ^ Перейти обратно: а б с д и МакХейзер-Уильямс М., Окицу С., Ван Н., МакХейзер-Уильямс Л. (декабрь 2011 г.). «Молекулярное программирование памяти В-клеток» . Обзоры природы. Иммунология . 12 (1): 24–34. дои : 10.1038/nri3128 . ПМЦ 3947622 . ПМИД 22158414 .
- ^ Перейти обратно: а б с д Куросаки Т., Кометани К., Исэ В. (март 2015 г.). «В-клетки памяти». Обзоры природы. Иммунология . 15 (3): 149–159. дои : 10.1038/nri3802 . ПМИД 25677494 . S2CID 20825732 .
- ^ Янмац, Майя; Кеса, Гун; Неттерлид, Ева; Бюисман, Анн-Мари; Торстенссон, Ригмор; Альборг, Никлас (31 мая 2013 г.). «Оптимизация анализа ELISpot IgG B-клеток человека для анализа ответов B-клеток, индуцированных вакциной» . Журнал иммунологических методов . 391 (1): 50–59. дои : 10.1016/j.jim.2013.02.009 . ISSN 0022-1759 . ПМИД 23454005 .
- ^ Рибурту Б., Зандеки М. (июнь 2015 г.). «Морфология плазматических клеток при множественной миеломе и связанных с ней заболеваниях». Морфология . 99 (325): 38–62. дои : 10.1016/j.morpho.2015.02.001 . ПМИД 25899140 . S2CID 1508656 .
- ^ Перейти обратно: а б с д Баумгарт Н. (январь 2011 г.). «Двойная жизнь клетки B-1: самореактивность выбирает защитные эффекторные функции». Обзоры природы. Иммунология . 11 (1): 34–46. дои : 10.1038/nri2901 . ПМИД 21151033 . S2CID 23355423 .
- ^ Пиллаи С., Кариаппа А., Моран С.Т. (1 января 2005 г.). «Б-клетки маргинальной зоны». Ежегодный обзор иммунологии . 23 (1): 161–196. doi : 10.1146/annurev.immunol.23.021704.115728 . ПМИД 15771569 .
- ^ Перейти обратно: а б с д и Россер ЕС, Маури С (апрель 2015 г.). «Регуляторные В-клетки: происхождение, фенотип и функции» . Иммунитет . 42 (4): 607–612. doi : 10.1016/j.immuni.2015.04.005 . ПМИД 25902480 .
- ^ Перейти обратно: а б Янаба К., Буазиз Дж.Д., Мацусита Т., Магро СМ, Сент-Клер Э.В., Теддер Т.Ф. (июнь 2008 г.). «Вклад B-лимфоцитов в аутоиммунные заболевания человека». Иммунологические обзоры . 223 (1): 284–299. дои : 10.1111/j.1600-065X.2008.00646.x . ПМИД 18613843 . S2CID 11593298 .
- ^ Шаффер А.Л., Янг Р.М., Штаудт Л.М. (1 января 2012 г.). «Патогенез В-клеточных лимфом человека» . Ежегодный обзор иммунологии . 30 (1): 565–610. doi : 10.1146/annurev-immunol-020711-075027 . ПМЦ 7478144 . ПМИД 22224767 .
- ^ Кастильо Джей-Джей (декабрь 2016 г.). «Заболевания плазматических клеток». Первичный уход . 43 (4): 677–691. дои : 10.1016/j.pop.2016.07.002 . ПМИД 27866585 .
- ^ Грамматикос Александрос, Донати Мэтью, Джонстон Сара Л., Гомпельс Марк М. Дефицит периферических B-клеток и предрасположенность к вирусным инфекциям: парадигма иммунодефицита. Frontiers in Immunology (12) 2021 https://www.frontiersin.org/articles/10.3389/fimmu.2021.731643 DOI=10.3389/fimmu.2021.731643
- ^ Кулис М., Меркель А., Хит С., Кейрос А.С., Шайлер Р.П., Кастеллано Г. и др. (июль 2015 г.). «Полногеномный отпечаток метилома ДНК во время дифференцировки B-клеток человека» . Природная генетика . 47 (7): 746–756. дои : 10.1038/ng.3291 . ПМЦ 5444519 . ПМИД 26053498 .