Тяжелая цепь иммуноглобулина

( Тяжелая цепь иммуноглобулина IgH ) представляет собой большую полипептидную субъединицу антитела ( иммуноглобулина). В геноме человека локусы гена IgH находятся на хромосоме 14.
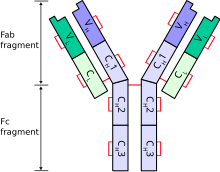
Типичное антитело состоит из двух тяжелых цепей иммуноглобулина (Ig) и двух легких цепей Ig . Существует несколько различных типов тяжелых цепей, которые определяют класс или изотип антитела. Эти типы тяжелых цепей различаются у разных животных. Все тяжелые цепи содержат ряд доменов иммуноглобулина , обычно с одним вариабельным доменом ( VH ), важным для связывания антигена , и несколькими константными доменами (CH1 , CH2 и т. д.). Производство жизнеспособной тяжелой цепи является ключевым этапом созревания В-клеток. Если тяжелая цепь способна связываться с суррогатной легкой цепью и перемещаться к плазматической мембране, тогда развивающаяся В-клетка может начать производить свою легкую цепь. [2]
Тяжелая цепь не всегда обязательно связывается с легкой цепью. Пре-В-лимфоциты могут синтезировать тяжелую цепь в отсутствие легкой цепи, что затем позволяет тяжелой цепи связываться с белком, связывающим тяжелую цепь. [3]
У млекопитающих
[ редактировать ]Классы
[ редактировать ]Существует пять типов тяжелой цепи иммуноглобулина млекопитающих: γ, δ, α, μ и ε. [4] Они определяют классы иммуноглобулинов: IgG , IgD , IgA , IgM и IgE соответственно.
- Тяжелые цепи α и γ содержат примерно 450 аминокислот.
- Тяжелые цепи μ и ε содержат примерно 550 аминокислот. [4]
Регионы
[ редактировать ]Каждая тяжелая цепь имеет две области:
- константная область (которая одинакова для всех иммуноглобулинов одного класса, но различается между классами).
- , вариабельная область которая различается у разных В-клеток , но одинакова для всех иммуноглобулинов, продуцируемых одной и той же В-клеткой или клоном В-клетки . Вариабельный домен любой тяжелой цепи состоит из одного домена иммуноглобулина. Эти домены имеют длину около 110 аминокислот. [6]
Коровы
[ редактировать ]Коровы, в частности Bos taurus , демонстрируют вариацию общей темы млекопитающих, в которой область CDR H3 тяжелой цепи адаптировалась для производства дивергентного репертуара антител, которые представляют собой поверхность взаимодействия антигена «стебель и выступ» вместо более знакомой двухвалентной поверхности кончика. . [7] CDR крупного рогатого скота необычно длинный и содержит уникальные атрибуты последовательности, которые поддерживают продукцию парных цистеина остатков во время соматической гипермутации . [7] Таким образом, если у людей этап соматической гипермутации нацелен на процесс рекомбинации V(D)J , то у коров целью является создание разнообразных дисульфидных связей и генерация уникальных наборов петель, которые взаимодействуют с антигеном. [7] Предполагаемой эволюционной движущей силой этой вариации является наличие гораздо более разнообразной микробной среды в пищеварительной системе коров вследствие того, что они являются жвачными животными . [7]
В рыбе
[ редактировать ]Челюстные рыбы , по-видимому, являются наиболее примитивными животными, способными вырабатывать антитела, подобные тем, которые описаны для млекопитающих. [8] Однако у рыб нет того набора антител, который есть у млекопитающих. [9] к настоящему времени идентифицированы три различные тяжелые цепи Ig У костистых рыб .
- Первой идентифицированной была тяжелая цепь ц (или мю ), которая присутствует во всех челюстных рыбах и является тяжелой цепью того, что считается первичным иммуноглобулином. Полученное антитело IgM секретируется рыб в виде тетрамера у костистых вместо типичного пентамера, обнаруженного у млекопитающих и акул. [ нужна ссылка ]
- Тяжелая цепь (δ) IgD была первоначально идентифицирована у канального сома и атлантического лосося и в настоящее время хорошо документирована для многих костистых рыб. [10]
- Третий ген тяжелой цепи костистых Ig был идентифицирован совсем недавно и не похож ни на одну из тяжелых цепей, описанных до сих пор для млекопитающих. Эта тяжелая цепь обнаружена как у радужной форели (τ), так и у радужной форели (τ). [11] и рыбка данио (ζ), [12] потенциально может образовать отдельный изотип антитела (IgT или IgZ), который может предшествовать IgM в эволюционном плане.
Подобно ситуации, наблюдаемой у костистых рыб, у хрящевых рыб были идентифицированы три различных изотипа тяжелой цепи Ig . За исключением μ, эти изотипы тяжелой цепи Ig, по-видимому, уникальны для хрящевых рыб. Полученные антитела обозначаются IgW (также называемые IgX или IgNARC) и IgNAR ( рецептор нового антигена иммуноглобулина ). [13] [14] Последний тип представляет собой антитело с тяжелой цепью , антитело, лишенное легких цепей, и может использоваться для получения однодоменных антител , которые по существу представляют собой вариабельный домен (V NAR ) IgNAR. [15] [16] [17] Однодоменные антитела акул (V NAR ) к опухолевым или вирусным антигенам можно выделить из большой наивной V NAR библиотеки акулы-няньки с использованием технологии фагового дисплея . [16] [18]
IgW в настоящее время также обнаружен у группы лопастных рыб, включая латимерию и двоякодышащую рыбу. IgW1 и IgW2 латимерии имеют обычную структуру (VD)n-Jn-C, а также большое количество константных доменов. [19] [20]
У амфибий
[ редактировать ]Лягушки могут синтезировать IgX и IgY. [21]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Архивная копия» . Архивировано из оригинала 19 апреля 2007 года . Проверено 20 апреля 2007 г.
{{cite web}}: CS1 maint: архивная копия в заголовке ( ссылка ) [ нужна полная цитата ] - ^ Мортенссон, Иллинойс; Кередиг, Р. (23 января 2017 г.). «Роль суррогатной легкой цепи и рецептора пре-В-клеток в развитии В-клеток мыши» . Иммунология . 101 (4): 435–441. дои : 10.1046/j.1365-2567.2000.00151.x . ISSN 0019-2805 . ПМК 2327112 . ПМИД 11122446 .
- ^ Хаас, Ингрид Г.; Вабль, Матиас (1983). «Белок, связывающий тяжелую цепь иммуноглобулина». Природа . 306 (5941): 387–9. Бибкод : 1983Natur.306..387H . дои : 10.1038/306387a0 . ПМИД 6417546 . S2CID 4247626 .
- ^ Jump up to: а б с Джейнвей, Калифорния, младший ; и др. (2001). Иммунобиология (5-е изд.). Издательство «Гирлянда». ISBN 0-8153-3642-Х . (полный текст в электронной форме через NCBI Bookshelf) . [ нужна страница ]
- ^ Гав, Дженни М.; Бертон, Деннис Р. (2004). «Взаимодействие человеческого антитела и Fc-рецептора, освещенное кристаллическими структурами». Обзоры природы Иммунология . 4 (2): 89–99. дои : 10.1038/nri1266 . ПМИД 15040582 . S2CID 30584218 .
- ^ «Биологический проект» . Структура антител . Университет Аризоны . Проверено 27 мая 2020 г.
- ^ Jump up to: а б с д Ван, Фэн; Экерт, Дамиан К.; Ахмад, Инша; Ю, Вэньли; Чжан, Юн; Базирган, Омар; Торкамани, Али; Раудсепп, Терье; Мванги, Вайтака; Кришителло, Майкл Ф.; Уилсон, Ян А.; Шульц, Питер Г.; Смайдер, Вон В. (2013). «Изменение разнообразия антител» . Клетка . 153 (6): 1379–93. дои : 10.1016/j.cell.2013.04.049 . ПМК 4007204 . ПМИД 23746848 .
- ^ Гены тяжелой и легкой цепи рыбы . [ нужна полная цитата ] Архивировано 23 марта 2007 г. в Wayback Machine.
- ^ Бенгтен, Ева; Клем, Л. Уильям; Миллер, Норман В.; Уорр, Грегори В.; Уилсон, Мелани (2006). «Канальные иммуноглобулины сома: репертуар и экспрессия». Развивающая и сравнительная иммунология . 30 (1–2): 77–92. дои : 10.1016/j.dci.2005.06.016 . ПМИД 16153707 .
- ^ Солем, Стейн Торе; Стенвик, Йорген (2006). «Развитие репертуара антител у костистых рыб - обзор с акцентом на лососевых и Gadus morhua L». Развивающая и сравнительная иммунология . 30 (1–2): 57–76. дои : 10.1016/j.dci.2005.06.007 . ПМИД 16084588 .
- ^ Хансен, доктор юридических наук; Лэндис, Эд; Филлипс, РБ (2005). «Открытие уникального изотипа тяжелой цепи Ig (IgT) у радужной форели: значение для особого пути развития B-клеток у костистых рыб» . Труды Национальной академии наук . 102 (19): 6919–6924. Бибкод : 2005PNAS..102.6919H . дои : 10.1073/pnas.0500027102 . JSTOR 3375456 . ПМЦ 1100771 . ПМИД 15863615 .
- ^ Данилова, Надя; Буссманн, Йерун; Джекош, Керстин; Штайнер, Лиза А. (2005). «Локус тяжелой цепи иммуноглобулина у рыбок данио: идентификация и экспрессия ранее неизвестного изотипа, иммуноглобулина Z». Природная иммунология . 6 (3): 295–302. дои : 10.1038/ni1166 . ПМИД 15685175 . S2CID 5543330 .
- ^ Дули, Х.; Флайник, М.Ф. (2006). «Развитие репертуара антител у хрящевых рыб». Развивающая и сравнительная иммунология . 30 (1–2): 43–56. дои : 10.1016/j.dci.2005.06.022 . ПМИД 16146649 .
- ^ Симмонс, Дэвид П.; Абрегу, Фиона А.; Кришнан, Уша В.; Пролл, Дэвид Ф.; Стрельцов Виктор А.; Даути, Лариса; Хаттарки, Меган К.; Наттолл, Стюарт Д. (2006). «Стратегии димеризации фрагментов однодоменных антител акулы IgNAR». Журнал иммунологических методов . 315 (1–2): 171–84. дои : 10.1016/j.jim.2006.07.019 . ПМИД 16962608 .
- ^ Весоловский, Януш; Алзогарай, Ванина; Рейелт, Ян; Унгер, Мэнди; Хуарес, Карла; Уррутия, Мариэла; Кауэрфф, Ана; Данкуа, Уэлбек; Риссик, Бьорн; Шойплейн, Феликс; Шварц, Николь; Адриуш, Сахиль; Бойер, Оливье; Семан, Мишель; Лицея, Алексей; Серрез, Дэвид В.; Гольдбаум, Фернандо А.; Хааг, Фридрих; Кох-Нольте, Фридрих (2009). «Однодоменные антитела: многообещающие экспериментальные и терапевтические инструменты в области инфекций и иммунитета» . Медицинская микробиология и иммунология . 198 (3): 157–74. дои : 10.1007/s00430-009-0116-7 . ПМК 2714450 . ПМИД 19529959 .
- ^ Jump up to: а б Фэн, Минцянь; Бянь, Хэцзяо; У, Сяолинь; Фу, Тяньюнь; Фу, Ин; Хонг, Джессика; Флеминг, Брайан Д.; Флайник, Мартин Ф.; Хо, Митчелл (январь 2019 г.). «Создание и анализ секвенирования следующего поколения большой библиотеки однодоменных антител VNAR, отображаемой на фагах, от шести наивных акул-нянек» . Терапия антителами . 2 (1): 1–11. дои : 10.1093/abt/tby011 . ISSN 2516-4236 . ПМК 6312525 . ПМИД 30627698 .
- ^ английский, Хэцзяо; Хонг, Джессика; Хо, Митчелл (2020). «Древние виды предлагают современные методы лечения: обновленная информация о последовательностях однодоменных антител акулы VNAR, фаговых библиотеках и потенциальных клинических применениях» . Терапия антителами . 3 (1): 1–9. дои : 10.1093/abt/tbaa001 . ПМК 7034638 . ПМИД 32118195 .
- ^ Ли, Дэн; английский, Хэцзяо; Хонг, Джессика; Лян, Тяньючжоу; Мерлино, Гленн; Дэй, Чи-Пин; Хо, Митчелл (21 июля 2021 г.). «Новая стратегия CAR-T на основе одного домена VNAR акулы, нацеленная на PD-L1, для лечения рака молочной железы и рака печени» : 2021.07.20.453144. дои : 10.1101/2021.07.20.453144 . S2CID 236203365 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Чжан, Тяньи; Такки, Лука; Вэй, Чжиго; Чжао, Яофэн; Салинас, Ирен (2014). «Внутриклассовая диверсификация генов тяжелой цепи иммуноглобулина у африканских двоякодышащих рыб» . Иммуногенетика . 66 (5): 335–51. дои : 10.1007/s00251-014-0769-2 . ПМК 4348116 . ПМИД 24676685 .
- ^ Ота, Т.; Раст, Дж.П.; Литман, Г.В.; Амемия, Коннектикут (2003). «Ограниченное по происхождению сохранение примитивного изотипа тяжелой цепи иммуноглобулина у дипноев раскрывает эволюционный парадокс» . Труды Национальной академии наук . 100 (5): 2501–6. Бибкод : 2003PNAS..100.2501O . дои : 10.1073/pnas.0538029100 . ПМК 151370 . ПМИД 12606718 .
- ^ Ду, Кристина С.; Машуф, Сара М.; Кришителло, Майкл Ф. (2012). «Поральная иммунизация африканской шпорцевой лягушки (Xenopus laevis) повышает уровень иммуноглобулина слизистой оболочки IgX» . Ветеринарная иммунология и иммунопатология . 145 (1–2): 493–8. дои : 10.1016/j.vetimm.2011.10.019 . ПМЦ 3273591 . ПМИД 22100190 .
Внешние ссылки
[ редактировать ]- Иммуноглобулин + тяжелые + цепи Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- Образовательный ресурс по анализу тяжелых цепей