Иммуноглобулин М
| Иммуноглобулин М | |||||||
|---|---|---|---|---|---|---|---|
| (пентамер) | |||||||
 | |||||||
| Тип белка | антитело | ||||||
| |||||||
Иммуноглобулин М ( IgM ) — самый крупный из нескольких изотипов антител иммуноглобулин (также известных как ) , вырабатываемых позвоночными . IgM — первое антитело, появляющееся в ответ на первоначальное воздействие антигена ; [1] [2] в результате чего его также называют антителом острой фазы. У людей и других млекопитающих, которые были изучены, плазмобласты в селезенке являются основным источником продукции специфических IgM. [3] [4]
История
[ редактировать ]В 1937 году было обнаружено антитело у лошадей, гипериммунизированных полисахаридом пневмококка, , которое было намного больше по размеру, чем типичный кроличий γ-глобулин. [5] с молекулярной массой 990 000 дальтон . [6] В связи с большим размером новое антитело первоначально называлось γ-макроглобулин, а впоследствии было названо IgM-M, что означает «макро». Домены V нормального иммуноглобулина очень гетерогенны, что отражает их роль в защите от большого количества инфекционных микробов, и эта гетерогенность затрудняет детальный структурный анализ IgM. Впоследствии были обнаружены два источника гомогенных IgM. Во-первых, белок с высокой молекулярной массой, продуцируемый некоторыми пациентами с множественной миеломой , был признан продуцируемым опухолью γ-макроглобулином, а поскольку опухоль представляет собой клон , вырабатываемые ею IgM гомогенны. [7] В 1960-е годы были разработаны методы индуцирования у мышей иммуноглобулинпродуцирующих опухолей (плазмоцитом), обеспечивающих источник гомогенных иммуноглобулинов различных изотипов, в том числе IgM (обзор опубликован в 1960-е годы). [8] ). Совсем недавно экспрессию сконструированных генов иммуноглобулинов в тканевой культуре можно было использовать для получения IgM со специфическими изменениями и, таким образом, для определения молекулярных требований к интересующим признакам. [ нужна ссылка ]
Структура
[ редактировать ]Иммуноглобулины состоят из легких и тяжелых цепей. Легкая цепь (λ или κ) представляет собой белок, состоящий из ~220 аминокислот, состоящий из вариабельного домена VL (сегмент примерно из 110 аминокислот) и константного домена CL (также длиной примерно 110 аминокислот). Тяжелая цепь μ IgM представляет собой белок, содержащий ~576 аминокислот, включает вариабельный домен (VH ~110 аминокислот), четыре отдельных домена константной области (Cμ1, Cμ2, Cμ3, Cμ4, каждый ~110 аминокислот) и « хвостовая часть» из ~20 аминокислот. Тяжелая цепь μ содержит олигосахариды по пяти остаткам аспарагина. Олигосахариды на IgM мыши и человека были частично охарактеризованы с помощью различных методов, включая ЯМР, связывание лектина, различные хроматографические системы и ферментативную чувствительность (см. [9] ). Структура олигосахаридов в каждом сайте различается в деталях, и преобладающие олигосахариды - двухантенные, трехантенные и с высоким содержанием маннозы - различаются в зависимости от сайта. [ нужна ссылка ]

А) Гетеродимер μL, иногда называемый полумером, с вариабельными (VH, VL) и константными областями (Cμ1, Cμ2, Cμ3, Cμ4tp; CL) доменами. Цистеины, которые обеспечивают дисульфидные связи между μ-цепями, показаны красными наконечниками стрелок, так что дисульфидная связь цистеина отображается как красный двойной наконечник стрелки (красный ромб). [ нужна ссылка ]
Б) «Мономер» IgM (мкл)2. Дисульфидные связи между доменами Cμ2 представлены двойной красной стрелкой.
C, D) Две модели пентамера IgM, содержащего J-цепь, которые в разное время появлялись в различных публикациях. Как и в (B), дисульфидные связи между доменами Cμ2 и дисульфидные связи между доменами Cμ4tp представлены двойной красной стрелкой; дисульфидные связи Cμ3 показаны (для наглядности) длинными двунаправленными стрелками. Связность, т. е. межцепочечная дисульфидная связь µ-цепей, обозначается как электрическая связность. В (C) дисульфидные связи Cμ3 соединяют μ-цепи параллельно дисульфидным связям Cμ4tp, и эти дисульфидные связи соединяют μ-цепи последовательно с дисульфидными связями Cμ2. В (D) дисульфидные связи Cμ2 и Cμ4tp соединяют μ-цепи параллельно, и оба типа соединяют μ-цепи последовательно с дисульфидными связями Cμ3. (Рисунок воспроизведен с разрешения издателя и авторов. [10] ).
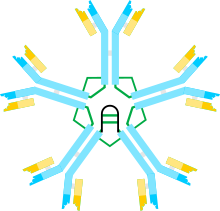
Мультимерная структура IgM схематически показана на фигуре 1. На фигуре 1А показан «гетеродимер», состоящий из одной легкой цепи, обозначенной L, и одной тяжелой цепи, обозначенной μ. Тяжелая и легкая цепи удерживаются вместе как дисульфидными связями (изображенными красными треугольниками), так и нековалентными взаимодействиями.
На рисунке 1B показаны две единицы мкл, связанные дисульфидной связью в доменах Cμ2; эту структуру (мкл)2 часто называют «мономером» IgM, поскольку она в некотором смысле аналогична структуре иммуноглобулина G (IgG) .
На основании скорости седиментации и внешнего вида на электронных микрофотографиях был сделан вывод, что IgM обычно встречается в виде «пентамера», т.е. полимера, состоящего из пяти «мономеров» [(мкл)2]5, и первоначально был описан модели на рисунках 1C и 1D, с дисульфидными связями между доменами Cμ3 и между хвостовыми частями. [11] [12] Также показано, что пентамерный IgM включает третий белок, J-цепь. J-цепь (J означает соединение) была обнаружена как ковалентно связанный компонент полимерных IgA и IgM. [13] [14] J-цепь представляет собой небольшой (~ 137 аминокислот) кислый белок. Как показано, J-цепь соединяет две ц-цепи посредством дисульфидных связей с участием цистеинов в хвостовых частях. [15]
Молекулярные требования для образования полимерных IgM
[ редактировать ]Первоначально ожидалось, что J-цепь будет важна для образования полимерных иммуноглобулинов, и действительно, полимеризация IgA сильно (но не абсолютно) зависит от J-цепи. [16] [17] Напротив, полимерный IgM эффективно образуется в отсутствие J-цепи. [18] [19]
Преобладающей формой человеческого и мышиного IgM является пентамер. Для сравнения: структура IgM лягушки (Xenopus) преимущественно гексамерная. [20] [21] IgM костистых рыб преимущественно тетрамерный, а IgM хрящевых рыб (главным образом акул) преимущественно пентамерный. [22] [23] Хотя пентамерная форма преобладает у мышей и людей, также наблюдалась гексамерная форма. [24] [25] Последующие исследования с использованием систем экспрессии рекомбинантной ДНК показали, что гексамер является основной формой мышиного IgM, когда IgM продуцируется в условиях, когда включение J-цепи предотвращается, либо путем производства IgM в клетках, у которых отсутствует J-цепь. [18] или путем производства IgM с тяжелой цепью μ, в которой отсутствует цистеин в хвостовой части. [26] [27] Таким образом, гексамерный IgM никогда не содержит J-цепи; пентамерный IgM может быть образован таким образом, чтобы он включал или не включал J-цепь. [28]
Важным различием между тяжелыми цепями μ и γ является наличие цистеинов для образования дисульфидных связей между тяжелыми цепями. В случае тяжелой γ-цепи единственные связи между γ образуются цистеинами в шарнире, и, соответственно, каждая γ-цепь связывается только с одной другой γ-цепью. Напротив, каждый из доменов Cμ2 и Cμ3 и хвостовой части включает цистеин, который образует дисульфидную связь с другой μ-цепью. Цистеины в доменах Cμ2 опосредуют образование мономерного IgM (мкл)2. Хвостик вместе с включенным в него цистеином необходим и достаточен для образования полимерных иммуноглобулинов. То есть удаление хвостовой части тяжелой цепи μ предотвращает образование полимерного IgM. [29] И наоборот, клетки, экспрессирующие тяжелую цепь γ, которая была модифицирована для включения хвостовой части, продуцируют полимерный IgG. [30] [31] [32]
Роль цистеина в домене Cμ3 более тонкая. На рисунках 1C и 1D представлены возможные модели пентамерного IgM. В этих моделях предполагается, что каждая µ-цепь связывает две другие µ-цепи. Однако ни одна из моделей сама по себе не может полностью объяснить структуру полимерного IgM. Например, модель на рисунке 1C предсказывает, что дисульфидная связь между доменами Cμ2 необходима для создания полимерного IgM с дисульфидной связью. Модель на рисунке 1D предсказывает, что дисульфидная связь между доменами Cμ3 важна. Полимерные IgM с дисульфидной связью все еще могут образовываться, если какой-либо из трех цистеинов отсутствует. В контексте моделей, в которых каждая µ-цепь взаимодействует только с двумя другими µ-цепями, эти результаты позволяют предположить, что некоторые молекулы похожи на рисунок 1C, а некоторые — на рисунок 1D. Однако наличие трех цистеинов для связывания между µ-цепями предполагает, что каждая из µ-цепей может связывать три другие µ-цепи, как показано на рисунке 2. В том же духе на рисунке 2C представлена модель пентамера, содержащего J-цепь, который отражает доказательства того, что J-цепь присоединяется к µ-цепям, которые не соединены с другими µ-цепями цистеинами в доменах Cμ3. Эти и другие модели, как регулярные, так и нерегулярные, обсуждаются в другом месте. [27] [33]
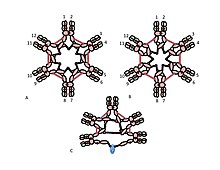
А, Б) На этих рисунках изображены две из многих возможных моделей дисульфидной связи между мю-цепями в гексамерном IgM. Как и на рисунке 1, дисульфидные связи Cμ2 и дисульфидные связи Cμ4tp представлены красными двойными стрелками, а дисульфидные связи Cμ3 представлены длинными двунаправленными стрелками. В обеих моделях А и Б каждый тип дисульфидной связи (Cμ2-Cμ2; Cμ3-Cμ3; Cμ4tp-Cμ4tp) соединяет ряды μ-цепей друг с другом. Методы различения этих и других моделей обсуждаются в ссылке [28].
C) Это изображение пентамерного IgM иллюстрирует, как J-цепь может быть связана с μ-цепями, которые не связаны дисульфидными связями Cμ3.
Пентамерный IgM обычно представляют как содержащий одну J-цепь на полимер, но на самом деле измерения стехиометрии J-цепи варьируются от одной молекулы J на полимер до трех молекул J на полимер. [34] [35] [36] [37] Широкий диапазон может быть обусловлен техническими проблемами, такими как неполная радиоактивная маркировка или неточное количественное определение линии Оухтерлони. Однако вариации могут быть также обусловлены гетерогенностью препаратов IgM, т.е. различные препараты могли существенно различаться по содержанию J-содержащих и J-дефицитных полимеров.
Третичная и четверичная структура постоянной области μ
[ редактировать ]Отдельные домены C2, C3 и C4tp были созданы независимо в E. coli , а затем изучены с использованием ряда подходов, включая скорость седиментации, рентгеновскую кристаллографию и ЯМР-спектроскопию , чтобы получить представление о детальной трехмерной структуре цепи. . Домены тяжелой цепи, как и домены других иммуноглобулинов, имеют характерные перекрывающиеся слои из семи нитей, которые стабилизированы внутридоменными дисульфидными связями. В целом константная область IgM имеет «грибовидную» форму: домены C2-C3 образуют диск, похожий на головку гриба, а домены C4tp выступают, как короткий стебель. [38]
Функция
[ редактировать ]IgM взаимодействует с рядом других физиологических молекул:
- может связывать комплемента компонент C1 и активировать классический путь , приводя к опсонизации антигенов IgM и цитолизу .
- IgM связывается с рецептором полииммуноглобулина (pIgR) в процессе, в результате которого IgM попадает на поверхности слизистых оболочек , например, в просвет кишечника , и в грудное молоко. Это связывание зависит от J-цепи. [39]
- Были обнаружены два других рецептора Fc, связывающих IgM, — Fcα/μ-R и Fcμ-R. Fcα/μ-R, как и pIgR, связывает полимерные IgM и IgA. Fcα/μ-R может опосредовать эндоцитоз , и его экспрессия в кишечнике предполагает участие в иммунитете слизистой оболочки. Fcμ-R (ранее известный как Toso/Faim3) связывает исключительно IgM и может опосредовать клеточное поглощение антигена, конъюгированного с IgM. [40] Инактивация соответствующих генов у нокаутных мышей приводит к появлению фенотипа , но физиологические функции этих рецепторов до сих пор неясны. [41]
Регуляция иммунного ответа
[ редактировать ]Специфические иммуноглобулины , вводимые животным вместе с их антигеном, могут влиять на реакцию антител к этому же антигену. [42] Эндогенные антитела, вырабатываемые после первичной иммунизации, также могут влиять на реакцию антител на повторную иммунизацию, что позволяет предположить, что аналогичные эффекты возникают и при физиологических условиях. «Регуляторные» эффекты могут быть как положительными, так и отрицательными. То есть, в зависимости от типа антигена и изотипа антитела, эффект может заключаться в подавлении или усилении гуморального ответа. Подобные эффекты хорошо иллюстрируются экспериментами по иммунизации ксеногенными (чужеродными) эритроцитами (эритроцитами). Например, при введении IgG вместе с ксеногенными эритроцитами такая комбинация вызывает практически полное подавление эритроцит-специфического гуморального ответа. Этот эффект используется в клинике для предотвращения иммунизации резус-отрицательных матерей против резус-положительных эритроцитов плода, а его использование резко снизило частоту гемолитической болезни у новорожденных. [43] В отличие от эффекта IgG, антигенспецифические IgM могут значительно усиливать гуморальный ответ, особенно в случае крупных антигенов. [44] Таким образом, когда IgM, специфичный для эритроцитов, вводят животным (включая человека) вместе с эритроцитами, индуцируется гораздо более сильный гуморальный ответ на эритроциты, чем при введении эритроцитов отдельно.Несколько доказательств указывают на то, что способность IgM активировать комплемент необходима для его усиления эффекта. То есть IgM-опосредованное усиление не происходит ни у животных, у которых истощен компонент комплемента C3, ни у мутантных животных, у которых отсутствуют рецепторы комплемента 1 и 2. Аналогичным образом, мутантный IgM, который не может активировать комплемент, не усиливает иммунный ответ.Возможным объяснением IgM-опосредованного усиления является то, что B-лимфоциты захватывают комплексы IgM-антиген-комплемент и транспортируют эти комплексы в области селезенки, где генерируются эффективные иммунные ответы. Поскольку IgM вырабатывается на ранних стадиях иммунного ответа, это может быть важно для инициации иммунного ответа. [ нужна ссылка ]
Синтез
[ редактировать ]В клетках зародышевой линии (сперматозоидах и яйцеклетках) гены, которые в конечном итоге будут кодировать иммуноглобулины, не находятся в функциональной форме (см. Рекомбинация V(D)J ). В случае тяжелой цепи три сегмента зародышевой линии, обозначенные V, D и J, лигируются вместе и присоединяются к ДНК, кодирующей константную область тяжелой цепи μ. В начале онтогенеза В-клетки экспрессируют как тяжелые цепи μ, так и δ; совместная экспрессия этих двух тяжелых цепей, каждая из которых несет один и тот же домен V, зависит от альтернативных сайтов сплайсинга и альтернативных сайтов добавления поли-А. На экспрессию других изотипов (γ, ε и α) влияет другой тип перестройки ДНК, процесс, называемый переключением класса иммуноглобулина . [45]
Клиническое значение
[ редактировать ]IgM — это первый иммуноглобулин, экспрессируемый у плода человека (около 20 недель). [46] и филогенетически самое раннее из появившихся антител. [47]
Антитела IgM появляются на ранних стадиях инфекции и обычно появляются вновь, в меньшей степени, после дальнейшего заражения. Антитела IgM не проходят через плаценту человека (только изотип IgG ). [48]
Эти два биологических свойства IgM делают его полезным при диагностике инфекционных заболеваний. Наличие антител IgM в сыворотке пациента указывает на недавнюю инфекцию, а в сыворотке новорожденного указывает на внутриутробную инфекцию (например, синдром врожденной краснухи ).
Развитие антидонорских IgM после трансплантации органов не связано с отторжением трансплантата, но может оказывать защитное действие. [49]
IgM в нормальной сыворотке часто связывается со специфическими антигенами даже при отсутствии предварительной иммунизации. [50] По этой причине IgM иногда называют «естественными антителами». Этот феномен, вероятно, обусловлен высокой авидностью IgM, которая позволяет ему заметно связываться даже со слабо перекрестно реагирующими антигенами , встречающимися в природе. эритроцитов, Например, антитела IgM, которые связываются с антигенами A и B могут образовываться в раннем возрасте в результате воздействия A- и B-подобных веществ, которые присутствуют в бактериях или, возможно, также в растительных материалах.
Антитела IgM главным образом ответственны за слипание ( агглютинацию ) эритроцитов, если реципиенту переливания крови вводят кровь, несовместимую с его группой крови .
Мутация мю -цепи внутри IgM вызывает аутосомно-рецессивную агаммаглобулинемию . [51]
Наличие IgM или, реже, IgG — один из облигатных критериев диагностики синдрома Шницлера . [52] [53]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Иммуноглобулин М». Словарь английского языка американского наследия (Четвертое изд.). Компания Хоутон Миффлин. 2004. ISBN 978-0618082308 .
- ^ Альбертс, Б.; Джонсон, А.; Льюис, Дж.; Уолтер, П.; Рафф, М.; Робертс, К. (2002). «Глава 24» . Молекулярная биология клетки (4-е изд.). Рутледж. ISBN 978-0-8153-3288-6 .
- ^ Каполунги, Ф.; Росадо, ММ; Синибальди, М.; Аранбуру, А.; Карсетти, Р. (2013). «Зачем нам нужны B-клетки памяти IgM?». Письма по иммунологии . 152 (2): 114–20. дои : 10.1016/j.imlet.2013.04.007 . ПМИД 23660557 .
- ^ Уильямс, Н.; О'Коннелл, PR (2008). «Глава 62». Краткая хирургическая практика Бейли и Лав (25-е изд.). ЦРК Пресс. п. 1102. ИСБН 9780340939321 .
- ^ Гейдельбергер, М.; Педерсен, нокаут (1937). «Молекулярная масса антител» . Журнал экспериментальной медицины . 65 (3): 393–414. дои : 10.1084/jem.65.3.393 . ПМК 2133497 . ПМИД 19870608 .
- ^ Кабат, Э.А. (1939). «Молекулярная масса антител» . Журнал экспериментальной медицины . 69 (1): 103–118. дои : 10.1084/jem.69.1.103 . ПМЦ 2133729 . ПМИД 19870830 .
- ^ Вальденстрем, Дж. (1943). «Начинающийся миеломатоз или «эссенциальная» гиперглобулинемия с фибриногенопенией - новый синдром?». Акта Медика Скандинавия . 142 (3–4): 216–247. дои : 10.1111/j.0954-6820.1944.tb03955.x .
- ^ Поттер, М. (2007). Ранняя история плазмоклеточных опухолей у мышей, 1954–1976 гг . Том. 98. стр. 17–51. дои : 10.1016/S0065-230X(06)98002-6 . ISBN 9780123738967 . ПМИД 17433907 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Моника, Ти Джей; Уильямс, С.Б.; Гучи, CF; Майорелла, Б.Л. (1995). «Характеристика гликозилирования человеческого IgM, продуцируемого гибридомой человека и мыши». Гликобиология . 5 (2): 175–185. дои : 10.1093/гликоб/5.2.175 . ПМИД 7780192 .
- ^ Хейман, Б.; Шульман, МЮ (2016). «Структура, функция и продукция иммуноглобулина М (IgM)». В Рэтклиффе, М. (ред.). Энциклопедия иммунобиологии . Том. 2. Эльзевир. стр. 1–14. дои : 10.1016/B978-0-12-374279-7.05001-3 . ISBN 978-0-12-374279-7 .
- ^ Бил, Д.; Файнштейн, А. (1969). «Исследования по снижению человеческого 19S иммуноглобулина М» . Биохимический журнал . 112 (2): 187–194. дои : 10.1042/bj1120187 . ПМЦ 1187691 . ПМИД 4979347 .
- ^ Мильштейн, КП; и др. (1975). «Межцепочечные дисульфидные мостики мышиного иммуноглобулина М» . Биохимический журнал . 151 (3): 615–624. дои : 10.1042/bj1510615 . ПМЦ 1172409 . ПМИД 766753 .
- ^ Халперн, М.С.; Кошланд, Мэн (1970). «Новая субъединица секреторного IgA». Природа . 228 (5278): 1276–1278. Бибкод : 1970Natur.228.1276H . дои : 10.1038/2281276a0 . ПМИД 5530654 . S2CID 36189290 .
- ^ Местецкий, Дж.; Зикин Дж.; Батлер, WT (1971). «Иммуноглобулин М, секреторный иммуноглобулин А: наличие общей полипептидной цепи, отличной от легких цепей». Наука . 171 (3976): 1163–1165. Бибкод : 1971Sci...171.1163M . дои : 10.1126/science.171.3976.1163 . ПМИД 5544873 . S2CID 6834561 .
- ^ Фрутигер, С.; и др. (1992). «Назначение дисульфидной связи в J-цепи человека и ее ковалентное спаривание с иммуноглобулином М». Биохимия . 31 (50): 12643–12647. дои : 10.1021/bi00165a014 . ПМИД 1472500 .
- ^ Йохансен, FE; Браатен, Р.; Брандцаег, П. (2000). «Роль J-цепи в образовании секреторного иммуноглобулина» . Скандинавский журнал иммунологии . 52 (3): 240–8. дои : 10.1046/j.1365-3083.2000.00790.x . ПМИД 10972899 .
- ^ Соренсен, В.; и др. (2000). «Структурные требования для включения J-цепи в человеческие IgM и IgA» . Международная иммунология . 12 (1): 19–27. дои : 10.1093/интимм/12.1.19 . ПМИД 10607746 .
- ^ Перейти обратно: а б Каттанео, А.; Нойбергер, М.С. (1987). «Полимерный иммуноглобулин М секретируется трансфектантами нелимфоидных клеток в отсутствие J-цепи иммуноглобулина» . Журнал ЭМБО . 6 (9): 2753–2758. дои : 10.1002/j.1460-2075.1987.tb02569.x . ПМЦ 553699 . ПМИД 3119328 .
- ^ Фазель, С.; Виерсма, Э.Дж.; Шульман, МД (1997). «Взаимодействие J-цепи и дисульфидной связи при сборке полимерного IgM» . Международная иммунология . 9 (8): 1149–1158. дои : 10.1093/интимм/9.8.1149 . hdl : 1807/11898 . ПМИД 9263012 .
- ^ Паркхаус, Р.; Асконас, бакалавр наук; Дурмашкин Р.Р. (1970). «Электронно-микроскопические исследования мышиного иммуноглобулина М; структура и восстановление после восстановления» . Иммунология . 18 (4): 575–584. ПМЦ 1455497 . ПМИД 5421036 .
- ^ Швагер, Дж.; Хаджи-Азлми, И. (1984). «Митоген-индуцированная дифференцировка В-клеток Xenopus laevis». Дифференциация . 27 (3): 182–188. дои : 10.1111/j.1432-0436.1984.tb01426.x . ПМИД 6334001 .
- ^ Филлатро, С.; и др. (2013). «Удивительное разнообразие классов Ig и репертуара В-клеток у костистых рыб» . Границы в иммунологии . 4 : 1–14. дои : 10.3389/fimmu.2013.00028 . ПМЦ 3570791 . ПМИД 23408183 .
- ^ Гетахун, А.; и др. (1999). «Влияние С-концевой последовательности μ-цепи на полимеризацию иммуноглобулина М» . Иммунология . 97 (3): 408–413. дои : 10.1046/j.1365-2567.1999.00797.x . ПМК 2326861 . ПМИД 10447761 .
- ^ Долдер, Ф. (1971). «Возникновение, выделение и межцепные мостики природного 7-S иммуноглобулина М в сыворотке человека». Биохимика и биофизика Acta . 236 (3): 675–685. ПМИД 4997811 .
- ^ Эскеланд, Т.; Кристенсен, ТБ (1975). «Молекулы IgM с J-цепью и без нее в сыворотке и после очистки, изученные с помощью ультрацентрифугирования, электрофореза и электронной микроскопии». Скандинавский журнал иммунологии . 4 (3): 217–228. дои : 10.1111/j.1365-3083.1975.tb02620.x . ПМИД 807966 . S2CID 8246173 .
- ^ Дэвис, AC; Ру, К.Х.; Шульман, МД (1988). «О строении полимерных IgM». Европейский журнал иммунологии . 18 (7): 1001–1008. дои : 10.1002/eji.1830180705 . ПМИД 3136022 . S2CID 34679165 .
- ^ Перейти обратно: а б Дэвис, AC; и др. (1989). «Межмолекулярная дисульфидная связь в IgM: эффекты замены остатков цистеина в тяжелой цепи μ» . Журнал ЭМБО . 8 (9): 2519–2526. дои : 10.1002/j.1460-2075.1989.tb08389.x . ПМК 401247 . ПМИД 2511005 .
- ^ Коллинз, К.; Цуй, ФРВ; Шульман, MJ (2002). «Дифференциальная активация комплемента человека и морской свинки пентамерными и гексамерными IgM» . Европейский журнал иммунологии . 32 (6): 1802–1810. doi : 10.1002/1521-4141(200206)32:6<1802::AID-IMMU1802>3.0.CO;2-C . ПМИД 12115664 .
- ^ Дэвис, AC; и др. (1989). «Мутации m H-цепи мыши, которые препятствуют сборке полимера» . Журнал иммунологии . 43 (4): 1352–1357. дои : 10.4049/jimmunol.143.4.1352 . ПМИД 2501393 . S2CID 40558731 .
- ^ Смит, РИФ; Колома, MJ; Моррисон, СЛ (1995). «Добавление мю-хвоста к IgG приводит к образованию полимерных антител с усиленными эффекторными функциями, включая опосредованный комплементом цитолиз с помощью IgG4» . Журнал иммунологии . 154 (5): 2226–2236. дои : 10.4049/jimmunol.154.5.2226 . PMID 7868896 . S2CID 10506582 .
- ^ Соренсен, В.; и др. (1996). «Влияние секреторных частей IgM и IgA на полимеризацию и секрецию IgM и IgG» . Журнал иммунологии . 156 (8): 2858–2865. дои : 10.4049/jimmunol.156.8.2858 . ПМИД 8609405 . S2CID 23601980 .
- ^ Смит, Р.; Моррисон, СЛ (1994). «Рекомбинантный полимерный IgG: подход к созданию более мощных антител». Природная биотехнология . 12 (7): 683–688. дои : 10.1038/nbt0794-683 . ПМИД 7764912 . S2CID 19055205 .
- ^ Виерсма, Э.Дж.; Шульман, МД (1995). «Сборка IgM: роль дисульфидной связи и нековалентных взаимодействий» . Журнал иммунологии . 154 (10): 5265–5272. дои : 10.4049/jimmunol.154.10.5265 . ПМИД 7730630 . S2CID 22148157 .
- ^ Шапюи, РМ; Кошланд, Мэн (1974). «Механизм полимеризации IgM» . Учеб. Натл. акад. наук. США 71 (3): 657–661. Бибкод : 1974PNAS...71..657C . дои : 10.1073/pnas.71.3.657 . ПМК 388071 . ПМИД 4207070 .
- ^ Михаеско, К.; Михаеско, Э.; Мецгер, Х. (1973). «Вариабельное содержание J-цепи в IgM человека» . Письма ФЭБС . 37 (2): 303–306. Бибкод : 1973FEBSL..37..303M . дои : 10.1016/0014-5793(73)80483-1 . ПМИД 4202824 . S2CID 41601548 .
- ^ Брандцаег, П. (1976). «Комплексное образование между секреторным компонентом и иммуноглобулином человека, связанное с содержанием в них J-цепи». Скандинавский журнал иммунологии . 5 (4): 411–419. дои : 10.1111/j.1365-3083.1976.tb00295.x . ПМИД 821140 . S2CID 39847718 .
- ^ Грабб, АО (1978). «Количественное определение J-цепи в биологических жидкостях человека с помощью простой иммунохимической процедуры». Акта Медика Скандинавия . 204 (1–6): 453–465. дои : 10.1111/j.0954-6820.1978.tb08473.x . ПМИД 104551 .
- ^ Мюллер, Р.; и др. (2013). «Структуры Fc-доменов IgM высокого разрешения раскрывают принципы образования его гексамеров» . Учеб. Натл. акад. наук. США 110 (25): 10183–10188. Бибкод : 2013PNAS..11010183M . дои : 10.1073/pnas.1300547110 . ПМЦ 3690842 . ПМИД 23733956 .
- ^ Йохансен, FE; Браатен, Р.; Брандцаег, П. (2000). «Роль J-цепи в образовании секреторного иммуноглобулина» . Скандинавский журнал иммунологии . 52 (3): 240–8. дои : 10.1046/j.1365-3083.2000.00790.x . ПМИД 10972899 .
- ^ Шима, Х.; и др. (2010). «Идентификация TOSO/FAIM3 как Fc-рецептора для IgM» . Межд. Иммунол. 22 (3): 149–56. дои : 10.1093/intimm/dxp121 . ПМИД 20042454 .
- ^ Ушида, Р.; и др. (2012). «Критическая роль рецептора Fc IgM в гомеостазе IgM, выживании B-клеток и гуморальных иммунных реакциях» . Учеб. Натл. акад. наук. США 109 (40): E2699–706. Бибкод : 2012PNAS..109E2699O . дои : 10.1073/pnas.1210706109 . ПМЦ 3479561 . ПМИД 22988094 .
- ^ Хейман, Б. (2013). Антитела-опосредованная регуляция гуморального иммунитета . в Ниммерджане, Ф. (ред.) Молекулярные и клеточные механизмы активности антител: Springer.
- ^ Урбаниак, С. Дж. и Грейсс, Массачусетс (2000). «RhD-гемолитическая болезнь плода и новорожденного». Кровавый преподобный . 14 (1): 33–61. дои : 10.1054/blre.1999.0123 . ПМИД 10805260 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сёрман А., Чжан Л., Дин З., Хейман Б. (2014). «Как антитела используют комплемент для регулирования реакции антител» . Мол. Иммунол . 61 (2): 79–88. дои : 10.1016/j.molimm.2014.06.010 . ПМИД 25001046 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мерфи, К.; Уивер, К. (2016). Иммунобиология Джейнвей . Нью-Йорк, штат Нью-Йорк: Garland Science / Тейлор и Фрэнсис. п. 195. ИСБН 9780815345053 .
- ^ ван Фюрт, Р.; Шуит, HR; Хиджманс, В. (1965). «Иммунологическое развитие плода человека» . Журнал экспериментальной медицины . 122 (6): 1173–88. дои : 10.1084/jem.122.6.1173 . ПМК 2138097 . ПМИД 4159036 .
- ^ Обзор медицинской физиологии Уильяма Фрэнсиса Ганонга
- ^ Мэтьюз, Т.Г.; О'Херлихи, К. (ноябрь 1978 г.). «Значение повышенного уровня иммуноглобулина М в пуповинной крови у младенцев, маленьких для гестационного возраста» . Архив болезней в детстве . 53 (11): 895–898. дои : 10.1136/adc.53.11.895 . ISSN 1468-2044 . ПМЦ 1545275 . ПМИД 727813 .
- ^ Макалистер, CC; и др. (2004). «Продукция защитных антидонорских IgM после трансплантации печени и почек с положительной перекрестной совместимостью» . Трансплантация печени . 10 (2): 315–9. дои : 10.1002/lt.20062 . ПМИД 14762873 .
- ^ Джаясекера, JP; Моузман, Э.А.; Кэрролл, MC (2007). «Природные антитела и комплемент опосредуют нейтрализацию вируса гриппа при отсутствии предварительного иммунитета» . Журнал вирусологии . 81 (7): 3487–94. дои : 10.1128/JVI.02128-06 . ПМК 1866020 . ПМИД 17202212 .
- ^ «Запись OMIM - № 601495 - АГАММАГЛОБУЛИНЕМИЯ 1, АУТОСОМНО-РЕЦЕССИВНАЯ; AGM1» . omim.org . Проверено 25 марта 2021 г.
- ^ Гусдорф, Л.; Липскер, Д. (17 июля 2017 г.). «Синдром Шницлера: обзор». Текущие отчеты по ревматологии . 19 (8): 46. дои : 10.1007/s11926-017-0673-5 . ПМИД 28718061 . S2CID 13780498 .
- ^ Саймон, А.; Асли, Б.; Браун-Фалько, М.; Конинг, Х. Де; Ферман, Ж.-П.; Граттан, К.; Краузе, К.; Лахманн, Х.; Ленорман, К.; Мартинес-Табоада, В.; Маурер, М.; Питерс, М.; Рицци, Р.; Ронджолетти, Ф.; Ружичка, Т.; Шницлер, Л.; Шуберт, Б.; Сибилия, Дж.; Липскер, Д. (2013). «Синдром Шницлера: диагностика, лечение и наблюдение» . Аллергия . 68 (5): 562–568. дои : 10.1111/all.12129 . ПМИД 23480774 . S2CID 12831354 .
Внешние ссылки
[ редактировать ]- Иммуноглобулин + М в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Справочник по дефициту иммуноглобулина М с сайта Medscape.com