Метилирование ДНК


Метилирование ДНК — это биологический процесс, при котором метильные группы добавляются к молекуле ДНК . Метилирование может изменить активность сегмента ДНК без изменения последовательности. Метилирование ДНК , локализованное в промоторе гена , обычно подавляет транскрипцию гена . У млекопитающих метилирование ДНК необходимо для нормального развития и связано с рядом ключевых процессов, включая геномный импринтинг , инактивацию Х-хромосомы , репрессию мобильных элементов , старение и канцерогенез .
По состоянию на 2016 год были обнаружены два азотистых основания, на которых происходит естественное ферментативное метилирование ДНК: аденин и цитозин . Модифицированные основания - N 6 -метиладенин, [1] 5-метилцитозин [2] и Н 4 -метилцитозин. [3]
| Немодифицированная база |  | 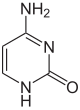 | ||||||
| Аденин, А | Цитозин, С | |||||||
| Модифицированные формы |  | 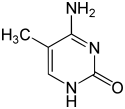 | 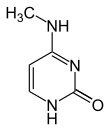 | |||||
| Н 6 -Метиладенин, 6 мА | 5-Метилцитозин, 5мС | Н 4 -Метилцитозин, 4мС | ||||||
Метилирование цитозина широко распространено как у эукариот , так и у прокариот , хотя скорость метилирования ДНК цитозина может сильно различаться у разных видов: 14% цитозинов метилированы у Arabidopsis thaliana , от 4% до 8% у Physarum , [4] 7,6% у Mus musculus , 2,3% у Escherichia coli , 0,03% у дрозофилы ; метилирование практически не обнаруживается у Dictyostelium ; [5] [6] и практически отсутствует (от 0,0002 до 0,0003%) при Caenorhabditis. [7] или грибы, такие как Saccharomyces cerevisiae и S. pombe (но не N. crassa ). [8] [9] : 3699 Метилирование аденина наблюдалось в ДНК бактерий, растений, а недавно и в ДНК млекопитающих. [10] [11] но уделялось значительно меньше внимания.
Метилирование цитозина с образованием 5-метилцитозина происходит в том же положении 5 пиримидинового кольца основания ДНК тимина , где расположена метильная группа ; это же положение отличает тимин от аналогичного основания РНК урацила , не имеющего метильной группы. Спонтанное дезаминирование превращает 5-метилцитозина его в тимин. Это приводит к несоответствию T:G. Механизмы восстановления затем восстанавливают исходную пару C:G; в качестве альтернативы они могут заменить G на A, превратив исходную пару C:G в пару T:A, эффективно изменив основание и введя мутацию. Это неправильно включенное основание не будет исправлено во время репликации ДНК, поскольку основанием ДНК является тимин. Если несоответствие не устранено и клетка вступает в клеточный цикл, цепь, несущая Т, будет дополнена А в одной из дочерних клеток, так что мутация станет постоянной. Почти повсеместное использование тимина исключительно в ДНК и урацила исключительно в РНК, возможно, возникло как механизм контроля ошибок, облегчающий удаление урацилов, образующихся в результате спонтанного дезаминирования цитозина. [12] Считалось, что метилирование ДНК, а также многие из его современных ДНК-метилтрансфераз произошли от примитивной активности метилирования РНК раннего мира, и это подтверждается несколькими доказательствами. [13]
У растений и других организмов метилирование ДНК обнаруживается в трех различных контекстах последовательностей: CG (или CpG ), CHG или CHH (где H соответствуют A, T или C). Однако у млекопитающих метилирование ДНК почти исключительно обнаруживается в динуклеотидах CpG, при этом цитозины на обеих цепях обычно метилируются. Однако не-CpG-метилирование может наблюдаться в эмбриональных стволовых клетках . [14] [15] [16] и также был показан в развитии нервной системы . [17] Более того, не-CpG-метилирование также наблюдалось в гемопоэтических клетках-предшественниках, и оно происходило главным образом в контексте последовательности CpApC. [18]
метилирования функция Консервативная ДНК

Ландшафт метилирования ДНК позвоночных очень специфичен по сравнению с другими организмами. У млекопитающих около 75% динуклеотидов CpG метилированы в соматических клетках . [19] а метилирование ДНК появляется как состояние по умолчанию, которое необходимо специально исключить из определенных мест. [16] [20] Напротив, геном большинства растений, беспозвоночных, грибов или протистов демонстрирует «мозаичный» характер метилирования, когда нацелены только на определенные геномные элементы, и они характеризуются чередованием метилированных и неметилированных доменов. [21] [22]

Высокое метилирование CpG в геномах млекопитающих имеет эволюционную цену, поскольку увеличивает частоту спонтанных мутаций. У цитозинов часто происходит потеря аминогрупп с различными последствиями в зависимости от их метилирования. Метилированные остатки C со временем самопроизвольно дезаминируются с образованием остатков T; следовательно, динуклеотиды CpG постепенно дезаминируются до динуклеотидов TpG, о чем свидетельствует недостаточная представленность динуклеотидов CpG в геноме человека (они встречаются только с 21% ожидаемой частоты). [23] (С другой стороны, спонтанное дезаминирование неметилированных остатков C приводит к образованию остатков U, и это изменение быстро распознается и восстанавливается клеткой.)
CpG острова [ править ]
У млекопитающих единственным исключением из этого глобального истощения CpG является определенная категория последовательностей, богатых GC и CpG, называемых CpG-островками, которые обычно неметилированы и, следовательно, сохраняют ожидаемое содержание CpG. [24] Островки CpG обычно определяются как области с: 1) длиной более 200 п.н., 2) содержанием G+C более 50%, 3) соотношением наблюдаемого и ожидаемого CpG более 0,6, хотя иногда используются и другие определения. [25] Если исключить повторяющиеся последовательности, в геноме человека насчитывается около 25 000 CpG-островков, 75% из которых имеют длину менее 850 пар оснований. [23] Они являются основными регуляторными единицами, и около 50% CpG-островков расположены в областях промоторов генов, а еще 25% лежат в телах генов, часто служа альтернативными промоторами. И наоборот, около 60-70% генов человека имеют островок CpG в своей промоторной области. [26] [27] Большинство CpG-островков конститутивно неметилированы и обогащены пермиссивной модификацией хроматина, такой как метилирование H3K4 . В соматических тканях метилировано лишь 10% CpG-островков, большая часть которых расположена в межгенных и внутригенных регионах. [ нужна ссылка ]
Репрессия CpG плотных - промоторов
Метилирование ДНК, вероятно, в некоторой степени присутствовало у очень ранних предков эукариот. Практически в каждом проанализированном организме метилирование промоторных областей отрицательно коррелирует с экспрессией генов. [21] [28] CpG-плотные промоторы активно транскрибируемых генов никогда не метилируются, но, наоборот, транскрипционно молчащие гены не обязательно несут метилированный промотор. У мышей и человека около 60–70% генов имеют островок CpG в своей промоторной области, и большинство этих островков CpG остаются неметилированными независимо от транскрипционной активности гена как в дифференцированных, так и в недифференцированных типах клеток. [29] [30] Следует отметить, что если метилирование ДНК CpG-островков однозначно связано с репрессией транскрипции, то функция метилирования ДНК в CG-бедных промоторах остается неясной; хотя существует мало доказательств того, что это может быть функционально значимо. [31]
Метилирование ДНК может влиять на транскрипцию генов двумя способами. Во-первых, само метилирование ДНК может физически препятствовать связыванию транскрипционных белков с геном. [32] и, во-вторых, что, вероятно, более важно, метилированная ДНК может быть связана с белками, известными как белки метил-CpG-связывающего домена (MBD). Белки MBD затем рекрутируют в локус дополнительные белки, такие как деацетилазы гистонов и другие белки ремоделирования хроматина , которые могут модифицировать гистоны , тем самым образуя компактный, неактивный хроматин, называемый гетерохроматином . Эта связь между метилированием ДНК и структурой хроматина очень важна. В частности, потеря метил-CpG-связывающего белка 2 (MeCP2) связана с синдромом Ретта ; а белок 2 метил-CpG-связывающего домена (MBD2) опосредует подавление транскрипции гиперметилированных генов при «раке». [ нужна ссылка ]
Подавление мобильных элементов [ править ]
Метилирование ДНК является мощным репрессором транскрипции, по крайней мере, в плотных контекстах CpG. Транскрипционная репрессия генов, кодирующих белки, по существу ограничивается очень специфическими классами генов, которые должны постоянно молчать и почти во всех тканях. Хотя метилирование ДНК не обладает той гибкостью, которая необходима для точной настройки регуляции генов, его стабильность идеальна для обеспечения постоянного молчания мобильных элементов . [33] Контроль транспозонов — одна из древнейших функций метилирования ДНК, которая свойственна животным, растениям и многочисленным протистам. [34] Предполагается даже, что метилирование ДНК возникло именно с этой целью. [35]
Расширение генома [ править ]
Известно, что метилирование ДНК мобильных элементов связано с расширением генома. Однако эволюционный драйвер расширения генома остается неизвестным. Существует четкая корреляция между размером генома и CpG, позволяющая предположить, что метилирование ДНК мобильных элементов привело к заметному увеличению массы ДНК. [36]
генного тела высокотранскрибируемых Метилирование генов
Функция, которая кажется даже более консервативной, чем замалчивание транспозонов, положительно коррелирует с экспрессией генов. Почти у всех видов, у которых присутствует метилирование ДНК, метилирование ДНК особенно богато в организме высоко транскрибируемыми генами. [21] [28] Функция метилирования тела гена недостаточно изучена. Ряд данных свидетельствует о том, что он может регулировать сплайсинг. [37] и подавляют активность внутригенных транскрипционных единиц (криптических промоторов или мобильных элементов). [38] Метилирование гена-тельца, по-видимому, тесно связано с метилированием H3K36. У дрожжей и млекопитающих метилирование H3K36 сильно обогащает организм хорошо транскрибируемыми генами. По крайней мере, у дрожжей H3K36me3 привлекает ферменты, такие как деацетилазы гистонов, для конденсации хроматина и предотвращения активации загадочных стартовых сайтов. [39] У млекопитающих домены PWWP DNMT3a и DNMT3b связываются с H3K36me3, и оба фермента рекрутируются в организм активно транскрибируемых генов. [ нужна ссылка ]
У млекопитающих [ править ]

Во время эмбрионального развития [ править ]
Паттерны метилирования ДНК в значительной степени стираются, а затем восстанавливаются между поколениями у млекопитающих. Почти все метилирования родителей стираются сначала во время гаметогенеза , а затем в раннем эмбриогенезе деметилирование и реметилирование , причем каждый раз происходит . Деметилирование в раннем эмбриогенезе происходит в предимплантационном периоде в две стадии — первоначально в зиготе , затем в течение первых нескольких эмбриональных циклов репликации морулы и бластулы . Затем на стадии имплантации эмбриона происходит волна метилирования, при этом островки CpG защищены от метилирования. Это приводит к глобальной репрессии и позволяет генам домашнего хозяйства экспрессироваться во всех клетках. На постимплантационной стадии характер метилирования зависит от стадии и ткани, причем изменения, которые определяют каждый отдельный тип клеток, сохраняются стабильно в течение длительного периода. [40] Исследования зачатков конечностей крыс во время эмбриогенеза еще раз проиллюстрировали динамическую природу метилирования ДНК в процессе развития. В этом контексте различия в глобальном метилировании ДНК наблюдались на разных стадиях развития и в условиях культивирования, что подчеркивает сложную регуляцию метилирования во время органогенеза и его потенциальные последствия для стратегий регенеративной медицины. [41]
не является необходимым Хотя метилирование ДНК само по себе для подавления транскрипции, тем не менее считается, что оно представляет собой «заблокированное» состояние, которое определенно инактивирует транскрипцию. В частности, метилирование ДНК, по-видимому, имеет решающее значение для поддержания моноаллельного молчания в контексте геномного импринтинга и инактивации Х-хромосомы . [42] [43] В этих случаях экспрессированные и молчащие аллели различаются по статусу метилирования, а потеря метилирования ДНК приводит к потере импринтинга и повторной экспрессии Xist в соматических клетках. Во время эмбрионального развития лишь немногие гены меняют свой статус метилирования, за важным исключением многих генов, специфически экспрессируемых в зародышевой линии. [44] Метилирование ДНК, по-видимому, абсолютно необходимо в дифференцированных клетках , поскольку нокаут любой из трех компетентных ДНК-метилтрансфераз приводит к эмбриональной или послеродовой летальности. Напротив, метилирование ДНК необязательно в недифференцированных типах клеток, таких как внутренняя клеточная масса бластоцисты, первичные зародышевые клетки или эмбриональные стволовые клетки. Поскольку метилирование ДНК, по-видимому, напрямую регулирует лишь ограниченное число генов, вопрос о том, как именно отсутствие метилирования ДНК вызывает гибель дифференцированных клеток, остается открытым вопросом.
Из-за феномена геномного импринтинга материнский и отцовский геномы по-разному маркируются и должны быть правильно перепрограммированы каждый раз, когда они проходят через зародышевую линию. Следовательно, во время гаметогенеза в первичных зародышевых клетках должны быть стерты и восстановлены исходные паттерны метилирования двуродительской ДНК в зависимости от пола передающего родителя. После оплодотворения отцовский и материнский геномы снова деметилируются и реметилируются (за исключением дифференциально метилированных участков, связанных с импринтированными генами). Это перепрограммирование, вероятно, необходимо для тотипотентности новообразованного эмбриона и стирания приобретенных эпигенетических изменений. [45]
При раке [ править ]
При многих болезненных процессах, таких как рак промотора генов , CpG-островки приобретают аномальное гиперметилирование, что приводит к подавлению транскрипции , которое может быть унаследовано дочерними клетками после клеточного деления. [46] Изменения метилирования ДНК признаны важным компонентом развития рака. Гипометилирование, как правило, возникает раньше и связано с хромосомной нестабильностью и потерей импринтинга, тогда как гиперметилирование связано с промоторами и может возникать вторично по отношению к молчанию генов (супрессоров онкогенов), но может быть мишенью для эпигенетической терапии . [47] В контексте развития динамические изменения в паттернах метилирования ДНК также имеют важное значение. Например, в зачатках конечностей крыс изменения статуса метилирования были связаны с разными стадиями хондрогенеза, что указывает на потенциальную связь между метилированием ДНК и прогрессированием определенных процессов развития. [41]
Глобальное гипометилирование также участвует в развитии и прогрессировании рака посредством различных механизмов. [48] Обычно наблюдается гиперметилирование генов-супрессоров опухолей и гипометилирование онкогенов . [49]
Как правило, при прогрессировании рака сотни генов подавляются или активируются . Хотя замалчивание некоторых генов при раке происходит в результате мутации, большая часть замалчивания канцерогенных генов является результатом изменения метилирования ДНК (см. Метилирование ДНК при раке ). Метилирование ДНК, вызывающее молчание при раке, обычно происходит в нескольких сайтах CpG на CpG-островках , которые присутствуют в промоторах генов, кодирующих белок. [ нужна ссылка ]
Измененная экспрессия микроРНК также подавляет или активирует многие гены при прогрессировании рака (см. микроРНК при раке ). Изменение экспрессии микроРНК происходит посредством гипер/гипометилирования сайтов CpG в островках CpG в промоторах, контролирующих транскрипцию микроРНК .
Замалчивание генов репарации ДНК посредством метилирования CpG-островков в их промоторах, по-видимому, особенно важно при прогрессировании рака (см. Метилирование генов репарации ДНК при раке ).
При атеросклерозе [ править ]
Эпигенетические модификации, такие как метилирование ДНК, связаны с сердечно-сосудистыми заболеваниями, включая атеросклероз . В моделях атеросклероза на животных сосудистая ткань, а также клетки крови, такие как мононуклеарные клетки крови, демонстрируют глобальное гипометилирование с геноспецифичными областями гиперметилирования. Полиморфизмы метилирования ДНК могут использоваться в качестве раннего биомаркера атеросклероза, поскольку они присутствуют до того, как наблюдаются поражения, что может стать ранним инструментом для обнаружения и предотвращения риска. [50]
Двумя типами клеток, на которые нацелен полиморфизм метилирования ДНК, являются моноциты и лимфоциты, которые испытывают общее гипометилирование. Одним из предполагаемых механизмов этого глобального гипометилирования является повышенный уровень гомоцистеина, вызывающий гипергомоцистеинемию , известный фактор риска сердечно-сосудистых заболеваний. Высокие уровни гомоцистеина в плазме ингибируют ДНК-метилтрансферазы, что вызывает гипометилирование. Гипометилирование ДНК влияет на гены, которые изменяют пролиферацию гладкомышечных клеток, вызывают дисфункцию эндотелиальных клеток и увеличивают количество медиаторов воспаления, которые имеют решающее значение в формировании атеросклеротических поражений. [51] Высокие уровни гомоцистеина также приводят к гиперметилированию CpG-островков в промоторной области гена рецептора альфа эстрогена (ERα), вызывая его понижающую регуляцию. [52] ERα защищает от атеросклероза благодаря своему действию в качестве супрессора роста, заставляя гладкомышечные клетки оставаться в состоянии покоя. [53] Таким образом, гиперметилирование промотора ERα позволяет клеткам гладкой мускулатуры интимы чрезмерно пролиферировать и способствовать развитию атеросклеротического поражения. [54]
Другим геном, статус метилирования которого изменяется при атеросклерозе, является переносчик монокарбоксилата (MCT3), который продуцирует белок, ответственный за транспорт лактата и других кетоновых тел из многих типов клеток, включая клетки гладких мышц сосудов. У пациентов с атеросклерозом наблюдается усиление метилирования CpG-островков в экзоне 2, что снижает экспрессию белка MCT3. Снижение уровня MCT3 ухудшает транспорт лактата и значительно увеличивает пролиферацию гладкомышечных клеток, что еще больше способствует атеросклеротическому поражению. В эксперименте ex vivo с использованием деметилирующего агента децитабина (5-аза-2-дезоксицитидин) было показано, что он индуцирует экспрессию MCT3 дозозависимым образом, поскольку все гиперметилированные сайты в CpG-островке экзона 2 становятся деметилированными после лечения. Это может служить новым терапевтическим средством для лечения атеросклероза, хотя исследований на людях до сих пор не проводилось. [55]
При сердечной недостаточности [ править ]
Помимо описанного выше атеросклероза , в поврежденном сердце человека были выявлены специфические эпигенетические изменения. Это может варьироваться в зависимости от этиологии заболевания. Например, при ишемической сердечной недостаточности изменения метилирования ДНК связаны с изменениями в экспрессии генов, которые могут направлять экспрессию генов, связанную с известными изменениями в сердечном метаболизме. [56] Необходимо изучить дополнительные формы сердечной недостаточности (например, диабетическую кардиомиопатию) и сопутствующие заболевания (например, ожирение), чтобы увидеть, насколько распространены эти механизмы. Наиболее поразительно то, что при недостаточности человеческого сердца эти изменения в метилировании ДНК связаны с расовым и социально-экономическим статусом, что в дальнейшем влияет на изменение экспрессии генов. [57] и может влиять на то, как следует лечить сердечную недостаточность у человека.
В старости [ править ]
У людей и других млекопитающих уровни метилирования ДНК можно использовать для точной оценки возраста тканей и типов клеток, формируя точные эпигенетические часы . [58]
Лонгитюдное исследование детей - близнецов показало, что в возрасте от 5 до 10 лет наблюдались расхождения в моделях метилирования из-за скорее экологических, чем генетических влияний. [59] С возрастом происходит глобальная потеря метилирования ДНК. [49]
В исследовании, в котором анализировались полные метиломы ДНК CD4 + У Т-клеток у новорожденного, человека в возрасте 26 лет и человека в возрасте 103 лет было обнаружено, что потеря метилирования пропорциональна возрасту. [60] Гипометилированные CpG, обнаруженные в ДНК долгожителей по сравнению с новорожденными, охватывали все компартменты генома (промоторы, межгенные , интронные и экзонные области). [60] Однако с возрастом некоторые гены становятся гиперметилированными, включая гены рецептора эстрогена , p16 и инсулиноподобного фактора роста 2 . [49]
В упражнении [ править ]
Было показано, что упражнения высокой интенсивности приводят к снижению метилирования ДНК в скелетных мышцах. [61] промотора Метилирование PGC-1α и PDK4 сразу снижалось после высокоинтенсивных упражнений, тогда как метилирование PPAR-γ снижалось только через три часа после тренировки. [61] В то же время шесть месяцев тренировок у мужчин среднего возраста, ранее ведущих малоподвижный образ жизни, привели к увеличению метилирования в жировой ткани . [62] Одно исследование показало возможное увеличение глобального метилирования геномной ДНК лейкоцитов при большей физической активности у неиспаноязычных людей. [63]
B- дифференцировке При клеток
Исследование, в котором изучали метилом В-клеток на протяжении их цикла дифференцировки с использованием полногеномного бисульфитного секвенирования (WGBS), показало, что происходит гипометилирование от самых ранних стадий до наиболее дифференцированных стадий. Наибольшая разница в метилировании наблюдается между стадиями В-клеток зародышевого центра и В-клеток памяти. Кроме того, это исследование показало, что существует сходство между опухолями В-клеток и долгоживущими В-клетками в признаках метилирования их ДНК. [18]
В мозгу [ править ]
В двух обзорах обобщены доказательства того, что изменения метилирования ДНК в нейронах головного мозга важны для обучения и памяти. [64] [65] Контекстуальное обусловливание страха (форма ассоциативного обучения) у животных, таких как мыши и крысы, происходит быстро и чрезвычайно надежно в создании воспоминаний. [66] У мышей [67] и у крыс [68] контекстуальное обусловливание страха в течение 1–24 часов связано с изменением метилирования нескольких тысяч ДНК-цитозинов в генах нейронов гиппокампа . Через двадцать четыре часа после контекстуального формирования страха 9,2% генов в нейронах гиппокампа крысы дифференциально метилированы. [68] У мышей, [67] при обследовании через четыре недели после кондиционирования метилирование и деметилирование гиппокампа были сброшены до исходных наивных условий. Гиппокамп . необходим для формирования воспоминаний, но воспоминания там не хранятся У таких мышей через четыре недели после контекстуального формирования страха CpG нейронах во время поддержания памяти происходили существенные дифференциальные метилирования и деметилирования в кортикальных , а в их передней поясной извилине было 1223 дифференциально метилированных гена. [67] В 2022 году были обобщены механизмы, управляющие новым метилированием ДНК и новым деметилированием ДНК в гиппокампе во время установления памяти. [69] Этот обзор также указал на механизмы, с помощью которых новые паттерны метилирования привели к появлению новых паттернов экспрессии информационной РНК . Эти новые информационные РНК затем переносились с помощью информационных частиц РНП (нейрональных гранул) в синапсы нейронов, где они могли транслироваться в белки. [69] Активные изменения в метилировании и деметилировании нейрональной ДНК, по-видимому, действуют как контроллеры синаптического масштабирования и трафика глутаматных рецепторов при обучении и формировании памяти . [64]
у млекопитающих ДНК-метилтрансферазы ( )

В клетках млекопитающих метилирование ДНК происходит главным образом в положении C5 динуклеотидов CpG и осуществляется за счет двух основных классов ферментативной активности – поддерживающего метилирования и метилирования de novo . [70]
Поддерживающая активность метилирования необходима для сохранения метилирования ДНК после каждого цикла репликации клеточной ДНК. Без ДНК-метилтрансферазы (DNMT) сам механизм репликации будет производить неметилированные дочерние цепи, что со временем приведет к пассивному деметилированию. DNMT1 — это предполагаемая поддерживающая метилтрансфераза, которая отвечает за копирование паттернов метилирования ДНК на дочерние цепи во время репликации ДНК. Мышиные модели с удаленными обеими копиями DNMT1 являются эмбрионально летальными примерно на 9-й день из-за необходимости активности DNMT1 для развития в клетках млекопитающих. [ нужна ссылка ]
Считается, что DNMT3a и DNMT3b являются метилтрансферазами de novo , которые устанавливают закономерности метилирования ДНК на ранних стадиях развития. DNMT3L представляет собой белок, гомологичный другим DNMT3, но не обладающий каталитической активностью. Вместо этого DNMT3L помогает метилтрансферазам de novo , увеличивая их способность связываться с ДНК и стимулируя их активность. У мышей и крыс есть третий функциональный фермент метилтрансфераза de novo , названный DNMT3C, который развился как паралог Dnmt3b путем тандемного дупликации у общего предка грызунов Muroidea. DNMT3C катализирует метилирование промоторов мобильных элементов во время раннего сперматогенеза, активность, которая, как было показано, важна для их эпигенетической репрессии и мужской фертильности. [71] [72] Пока неясно, полагаются ли у других млекопитающих, у которых нет DNMT3C (например, у людей), на DNMT3B или DNMT3A для метилирования de novo мобильных элементов в зародышевой линии. Наконец, DNMT2 (TRDMT1) был идентифицирован как гомолог ДНК-метилтрансферазы, содержащий все 10 мотивов последовательности, общих для всех ДНК-метилтрансфераз; однако DNMT2 (TRDMT1) не метилирует ДНК, а вместо этого метилирует цитозин-38 в антикодоновой петле РНК-переносчика аспарагиновой кислоты. [73]
Поскольку многие гены-супрессоры опухолей подавляются метилированием ДНК во время канцерогенеза , предпринимались попытки реэкспрессировать эти гены путем ингибирования DNMT. 5-Аза-2'-дезоксицитидин ( децитабин ) представляет собой аналог нуклеозида , который ингибирует DNMT, захватывая их в ковалентный комплекс на ДНК, предотвращая стадию β-элиминирования катализа, что приводит к деградации ферментов. Однако для того, чтобы децитабин был активным, он должен быть включен в геном клетки, что может вызвать мутации в дочерних клетках, если клетка не погибнет. Кроме того, децитабин токсичен для костного мозга, что ограничивает размер его терапевтического окна. Эти ловушки привели к разработке методов лечения антисмысловыми РНК, которые нацелены на DNMT, разрушая их мРНК и предотвращая их трансляцию . Однако в настоящее время неясно, достаточно ли воздействия только на DNMT1 для реактивации генов-супрессоров опухолей, подавленных метилированием ДНК. [ нужна ссылка ]
У растений [ править ]
Значительный прогресс был достигнут в понимании метилирования ДНК у модельного растения Arabidopsis thaliana . Метилирование ДНК у растений отличается от такового у млекопитающих: в то время как метилирование ДНК у млекопитающих в основном происходит на нуклеотиде цитозина в сайте CpG , у растений цитозин может метилироваться в сайтах CpG, CpHpG и CpHpH, где H представляет собой любой нуклеотид, но не гуанин. . [74] В целом ДНК арабидопсиса сильно метилирована, масс-спектрометрический анализ показал, что 14% цитозинов модифицировано. [9] : абстрактный Позже данные бисульфитного секвенирования показали, что около 25% сайтов CG Arabidopsis метилированы, но эти уровни варьируются в зависимости от географического местоположения образцов Arabidopsis (растения на севере метилированы более сильно, чем южные образцы). [75]
Основными ферментами ДНК-метилтрансфераз арабидопсиса , которые переносят и ковалентно присоединяют метильные группы к ДНК, являются DRM2, MET1 и CMT3. Белки DRM2 и MET1 имеют значительную гомологию с метилтрансферазами млекопитающих DNMT3 и DNMT1 соответственно, тогда как белок CMT3 уникален для царства растений. В настоящее время существует два класса ДНК-метилтрансфераз: 1) класс de novo или ферменты, которые создают новые метки метилирования на ДНК; 2) класс поддержания, который распознает метки метилирования на родительской цепи ДНК и передает новое метилирование на дочерние цепи после репликации ДНК. DRM2 — единственный фермент, который участвует в роли ДНК-метилтрансферазы de novo . Также было показано, что DRM2 наряду с MET1 и CMT3 участвует в поддержании меток метилирования посредством репликации ДНК. [76] Другие ДНК-метилтрансферазы экспрессируются в растениях, но не имеют известных функций (см. Базу данных хроматина ).
Уровни метилирования ДНК в масштабе всего генома широко варьируются между видами растений, а цитозины Arabidopsis, как правило, метилированы менее плотно, чем цитозины других растений. Например, ~92,5% цитозинов CpG метилированы у Beta vulgaris . [77] Паттерны метилирования также различаются в зависимости от контекста последовательности цитозина; повсеместно метилирование CpG выше, чем метилирование CHG и CHH, и метилирование CpG можно обнаружить как в активных генах, так и в мобильных элементах, тогда как CHG и CHH обычно относят к молчащим мобильным элементам. [78] [74]
Неясно, как клетка определяет места метилирования ДНК de novo , но данные свидетельствуют о том, что во многих (хотя и не во всех) местах РНК-направленное метилирование ДНК задействовано (RdDM). При RdDM специфические транскрипты РНК производятся из матрицы геномной ДНК, и эта РНК образует вторичные структуры, называемые двухцепочечными молекулами РНК. [79] Двухцепочечные РНК через пути малых интерферирующих РНК ( миРНК ) или микроРНК ( миРНК ) направляют метилирование ДНК de novo в исходном месте генома, в котором продуцируется РНК. [79] Считается, что механизм такого рода важен для защиты клеток от РНК-вирусов и/или транспозонов , которые часто образуют двухцепочечную РНК, которая может быть мутагенной для генома хозяина. Метилируя свои геномные местоположения посредством пока еще плохо изученного механизма, они отключаются и больше не активны в клетке, защищая геном от их мутагенного эффекта. Недавно было описано, что метилирование ДНК является основной детерминантой формирования эмбриогенных культур из эксплантов древесных растений и считается основным механизмом, объясняющим плохую реакцию зрелых эксплантов на соматический эмбриогенез у растений (Isah 2016). [ нужна ссылка ]
У насекомых [ править ]
Различные отряды насекомых демонстрируют различные закономерности метилирования ДНК: от почти необнаружимых уровней у мух до низких уровней у бабочек и более высоких у настоящих клопов и некоторых тараканов (до 14% всех сайтов CG у Blattella asahinai ). [80]
Функциональное метилирование ДНК обнаружено у медоносных пчел. [81] [82] Метки метилирования ДНК находятся в основном на теле гена, и современные мнения о функции метилирования ДНК заключаются в регуляции генов посредством альтернативного сплайсинга. [83]
Уровни метилирования ДНК у Drosophila melanogaster практически не обнаруживаются. [84] Чувствительные методы, применяемые к ДНК дрозофилы. Предполагают уровни в диапазоне 0,1–0,3% от общего цитозина. [85] Исследование 2014 года показало, что низкий уровень метилирования у плодовых мушек проявляется «в определенных коротких мотивах и не зависит от активности DNMT2». [86] Кроме того, высокочувствительные подходы масс-спектрометрии, [87] теперь продемонстрировали наличие низких (0,07%), но значительных уровней метилирования аденина на самых ранних стадиях эмбриогенеза дрозофилы.
У грибов [ править ]
Многие грибы имеют низкий уровень метилирования цитозина (от 0,1 до 0,5%), тогда как у других грибов метилировано до 5% генома. [88] Это значение, по-видимому, варьирует как среди видов, так и среди изолятов одного и того же вида. [89] Есть также доказательства того, что метилирование ДНК может участвовать в специфичном для каждого состояния контроле экспрессии генов у грибов. [ нужна ссылка ] Однако при пределе обнаружения 250 аттомолей с помощью сверхвысокочувствительной масс-спектрометрии метилирование ДНК не было подтверждено у одноклеточных видов дрожжей, таких как Saccharomyces cerevisiae или Schizosaccharomyces pombe , что указывает на то, что дрожжи не обладают этой модификацией ДНК. [9] : абстрактный
Хотя пивные дрожжи ( Saccharomyces ), делящиеся дрожжи ( Schizosaccharomyces ) и Aspergillus flavus [90] не имеют обнаруживаемого метилирования ДНК, модельный нитчатый гриб Neurospora crassa имеет хорошо охарактеризованную систему метилирования. [91] Несколько генов контролируют метилирование у Neurospora , а мутация ДНК-метилтрансферазы dim-2 устраняет все метилирование ДНК, но не влияет на рост или половое размножение. Хотя в геноме Neurospora очень мало повторяющейся ДНК, половина метилирования происходит в повторяющейся ДНК, включая реликты транспозонов и центромерную ДНК. Способность оценивать другие важные явления на генетическом фоне с дефицитом ДНК-метилазы делает Neurospora важной системой для изучения метилирования ДНК. [ нужна ссылка ]
У других эукариотов [ править ]
Метилирование ДНК практически отсутствует у Dictyostelium diskoidium. [92] где он, по-видимому, встречается примерно в 0,006% цитозинов. [6] Напротив, метилирование ДНК широко распространено у Physarum polycephalum. [93] где 5-метилцитозин составляет до 8% от общего количества цитозина. [4]
У бактерий [ править ]

Метилирование аденина или цитозина опосредуется системами рестрикционной модификации многих бактерий , в которых определенные последовательности ДНК периодически метилируются по всему геному. [95] Метилаза — это фермент, который распознает определенную последовательность и метилирует одно из оснований в этой последовательности или рядом с ней. Чужеродные ДНК (которые не метилированы таким образом), которые вводятся в клетку, разрушаются специфичными для последовательности ферментами рестрикции и расщепляются. Бактериальная геномная ДНК не распознается этими ферментами рестрикции. Метилирование нативной ДНК действует как своего рода примитивная иммунная система, позволяя бактериям защитить себя от заражения бактериофагом . [ нужна ссылка ]
E. coli ДНК-аденин-метилтрансфераза (Dam) представляет собой фермент массой ~32 кДа, не принадлежащий к системе рестрикции/модификации. Целевой последовательностью распознавания для Dam E. coli является GATC, поскольку метилирование происходит в положении N6 аденина в этой последовательности (G meATC). Три пары оснований, фланкирующие каждую сторону этого сайта, также влияют на связывание ДНК-Dam. Dam играет несколько ключевых ролей в бактериальных процессах, включая восстановление несоответствий, время репликации ДНК и экспрессию генов. В результате репликации ДНК статус GATC-сайтов в геноме E. coli меняется с полностью метилированного на гемиметилированный. Это связано с тем, что аденин, введенный в новую цепь ДНК, неметилирован. Повторное метилирование происходит в течение двух-четырех секунд, в течение которых исправляются ошибки репликации в новой цепи. Метилирование или его отсутствие является маркером, который позволяет аппарату репарации клетки различать матрицу и возникающие цепи. Было показано, что изменение активности Dam в бактериях приводит к увеличению частоты спонтанных мутаций. Жизнеспособность бактерий снижается у dam-мутантов, у которых также отсутствуют некоторые другие ферменты репарации ДНК, что является дополнительным доказательством роли Dam в репарации ДНК.
Одной из областей ДНК, которая дольше сохраняет свой полуметилированный статус, является точка начала репликации , которая имеет множество сайтов GATC. Это занимает центральное место в бактериальном механизме синхронизации репликации ДНК. SeqA связывается с началом репликации, изолируя его и тем самым предотвращая метилирование. Поскольку гемиметилированные начала репликации неактивны, этот механизм ограничивает репликацию ДНК до одного раза за клеточный цикл.
Экспрессия некоторых генов, например кодирующих экспрессию пилусов в E. coli , регулируется метилированием сайтов GATC в промоторной области генного оперона. Условия окружающей среды клеток сразу после репликации ДНК определяют, блокируется ли Dam от метилирования области, проксимальной или дистальной от области промотора. Как только паттерн метилирования создан, транскрипция гена пилуса блокируется во включенном или выключенном положении до тех пор, пока ДНК не будет снова реплицирована. В E. coli эти опероны пилей играют важную роль в вирулентности инфекций мочевыводящих путей. Было предложено [ кем? ] что ингибиторы Dam могут действовать как антибиотики.
С другой стороны, ДНК-цитозинметилаза нацелена на сайты CCAGG и CCTGG, чтобы метилировать цитозин в положении C5 (C meC(A/T) GG). Другой фермент метилазы, EcoKI, вызывает метилирование аденинов в последовательностях AAC(N 6 )GTGC и GCAC(N 6 )GTT.
Было показано, что у Clostridioides difficile метилирование ДНК по целевому мотиву CAAAAA влияет на споруляцию , ключевой этап передачи заболевания, а также на длину клеток, образование биопленок и колонизацию хозяина. [96]
Молекулярное клонирование [ править ]
Большинство штаммов, используемых молекулярными биологами, являются производными E. coli K-12 и обладают как Dam, так и Dcm, но существуют коммерчески доступные штаммы, которые являются dam-/dcm- (отсутствие активности какой-либо метилазы). Фактически, можно деметилировать ДНК, выделенную из штаммов dam+/dcm+, трансформируя ее в штаммы dam-/dcm-. Это поможет переварить последовательности, которые не распознаются чувствительными к метилированию ферментами рестрикции. [97] [98]
Фермент рестрикции DpnI может распознавать сайты 5'-GmeATC-3' и переваривать метилированную ДНК. Будучи таким коротким мотивом, он часто случайно встречается в последовательностях, и поэтому его основное использование для исследователей заключается в разрушении матричной ДНК после ПЦР (продукты ПЦР не имеют метилирования, поскольку в реакции нет метилаз). Аналогичным образом, некоторые коммерчески доступные рестриктазы чувствительны к метилированию родственных им сайтов рестрикции и, как упоминалось ранее, должны использоваться на ДНК, проходящей через штамм dam-/dcm-, чтобы обеспечить возможность разрезания. [ нужна ссылка ]
Обнаружение [ править ]
Метилирование ДНК можно обнаружить с помощью следующих анализов, используемых в настоящее время в научных исследованиях: [99]
- Масс-спектрометрия — очень чувствительный и надежный аналитический метод обнаружения метилирования ДНК. Однако MS, как правило, не информативен в отношении контекста последовательности метилирования, что ограничивает изучение функции этой модификации ДНК.
- ПЦР, специфичная для метилирования (MSP) , которая основана на химической реакции бисульфита натрия с ДНК, которая превращает неметилированные цитозины динуклеотидов CpG в урацил или UpG, с последующей традиционной ПЦР . [100] Однако метилированные цитозины не будут конвертироваться в этом процессе, и праймеры предназначены для перекрытия интересующего сайта CpG, что позволяет определить статус метилирования как метилированный или неметилированный.
- Полногеномное бисульфитное секвенирование , также известное как BS-Seq, которое представляет собой высокопроизводительный полногеномный анализ метилирования ДНК. Он основан на вышеупомянутом преобразовании геномной ДНК бисульфитом натрия, которая затем секвенируется на платформе секвенирования нового поколения . Полученные последовательности затем повторно выравниваются с эталонным геномом для определения статуса метилирования динуклеотидов CpG на основе несовпадений, возникающих в результате превращения неметилированных цитозинов в урацил.
- Ферментативный метил-seq (EM-seq) работает аналогично бисульфитному секвенированию, но использует ферменты APOBEC и TET2 для дезаминирования неметилированного цитозина в урацил перед секвенированием. Библиотеки EM-seq менее склонны к повреждению ДНК, чем библиотеки, обработанные бисульфитом. [101]
- Бисульфитное секвенирование с уменьшенным представлением , также известное как RRBS, знает несколько рабочих протоколов. Первый протокол RRBS назывался RRBS и нацелен примерно на 10% метилома, поэтому необходим эталонный геном. Позже появилось больше протоколов, которые смогли секвенировать меньшую часть генома и обеспечить более высокое мультиплексирование образцов. EpiGBS был первым протоколом, позволяющим мультиплексировать 96 образцов на одной дорожке секвенирования Illumina и при котором эталонный геном больше не требовался. Эталонная конструкция de novo, основанная на показаниях Уотсона и Крика, сделала популяционный скрининг SNP и SMP одновременно фактом.
- Анализ HELP , основанный на дифференциальной способности ферментов рестрикции распознавать и расщеплять метилированные и неметилированные участки ДНК CpG.
- Анализ GLAD-PCR , в основе которого лежит новый тип ферментов – сайт-специфические метилнаправленные ДНК-эндонуклеазы, которые гидролизуют только метилированную ДНК.
- Анализы ChIP-на-чипе , основанные на способности коммерческих антител связываться с белками, связанными с метилированием ДНК, такими как MeCP2.
- Геномное сканирование ориентиров рестрикции — сложный и в настоящее время редко используемый анализ, основанный на дифференциальном распознавании рестрикционными ферментами метилированных и неметилированных сайтов CpG; По своей концепции анализ аналогичен анализу HELP.
- Иммунопреципитация метилированной ДНК (MeDIP), аналогичная иммунопреципитации хроматина , иммунопреципитация используется для выделения метилированных фрагментов ДНК для ввода в методы обнаружения ДНК, такие как микрочипы ДНК (MeDIP-чип) или секвенирование ДНК (MeDIP-seq).
- Пиросеквенирование ДНК, обработанной бисульфитом. Это секвенирование ампликона, полученного с использованием обычного прямого праймера, но биотинилированного обратного праймера для ПЦР (гена выбора). Затем пиросеквенатор анализирует образец, денатурируя ДНК и добавляя в смесь по одному нуклеотиду в соответствии с последовательностью, заданной пользователем. Если есть несовпадение, его записывают и отмечают процент ДНК, для которого присутствует несовпадение. Это дает пользователю процент метилирования на островок CpG.
- Молекулярный анализ активности ДНК-аденин-метилтрансферазы - анализ, основанный на специфичности фермента рестрикции DpnI к полностью метилированным (метилирование аденина) сайтам GATC в олигонуклеотиде, меченном флуорофором и гасителем. Аденинметилтрансфераза метилирует олигонуклеотид, делая его субстратом для DpnI. Разрезание олигонуклеотида DpnI приводит к увеличению флуоресценции. [102] [103]
- Саузерн-блоттинг, чувствительный к метилу, аналогичен анализу HELP, но использует методы Саузерн-блоттинга для определения ген-специфичных различий в метилировании с использованием рестрикционных гидролизатов. Этот метод используется для оценки локального метилирования вблизи места связывания зонда.
- МетилCpG-связывающие белки (MBP) и слитые белки, содержащие только метилсвязывающий домен (MBD), используются для разделения нативной ДНК на метилированные и неметилированные фракции. Процентное метилирование отдельных CpG-островков можно определить путем количественного определения количества мишени в каждой фракции. [ нужна ссылка ] Чрезвычайно чувствительное обнаружение может быть достигнуто в тканях FFPE с помощью обнаружения на основе атрибуции.
- Анализ расплава высокого разрешения (HRM или HRMA) представляет собой аналитический метод после ПЦР . Целевую ДНК обрабатывают бисульфитом натрия, который химически превращает неметилированные цитозины в урацилы, при этом метилированные цитозины сохраняются. Затем проводят ПЦР-амплификацию с использованием праймеров, предназначенных для амплификации как метилированных, так и неметилированных матриц. После этой амплификации высокометилированные последовательности ДНК содержат большее количество сайтов CpG по сравнению с неметилированными матрицами, что приводит к другой температуре плавления, которую можно использовать для количественного обнаружения метилирования. [104] [105]
- Реконструкция метилирования древней ДНК - метод реконструкции метилирования ДНК с высоким разрешением по образцам древней ДНК. Метод основан на естественных процессах деградации, происходящих в древней ДНК: со временем метилированные цитозины расщепляются до тиминов, а неметилированные цитозины — до урацилов. Эта асимметрия сигналов деградации была использована для реконструкции полных карт метилирования неандертальцев и денисовцев . [106] В сентябре 2019 года исследователи опубликовали новый метод определения морфологических признаков на основе данных о метилировании ДНК. Авторам удалось показать, что связывание генов с пониженной регуляцией с фенотипами моногенных заболеваний, при которых нарушена одна или две копии гена, позволяет с точностью около 85% реконструировать анатомические признаки непосредственно по картам метилирования ДНК. [107]
- Анализ на удлинение одиночного нуклеотидного праймера, чувствительный к метилированию (msSNuPE), в котором используются внутренние праймеры, отжигающие прямые 5'-концы нуклеотида, подлежащего обнаружению. [108]
- Анализ метилирования Illumina измеряет локус-специфическое метилирование ДНК с использованием матричной гибридизации. ДНК, обработанная бисульфитом, гибридизуется с зондами на «BeadChips». Расширение однооснования с помощью меченых зондов используется для определения статуса метилирования сайтов-мишеней. [109] В 2016 году был выпущен Infinium MmethylationEPIC BeadChip, который исследует более 850 000 сайтов метилирования в геноме человека. [110]
регионы (DMR Дифференциально метилированные )
Дифференциально метилированные области , которые представляют собой области генома с разным статусом метилирования среди нескольких образцов (тканей, клеток, индивидуумов или других), рассматриваются как возможные функциональные области, участвующие в регуляции транскрипции генов. Идентификация DMR среди множества тканей (T-DMR) обеспечивает всестороннее исследование эпигенетических различий между тканями человека. [111] Например, эти метилированные области, уникальные для конкретной ткани, позволяют людям различать типы тканей, таких как сперма и вагинальная жидкость. Текущее исследование, проведенное Ли и др., показало, что DACT1 и USP49 положительно идентифицируют сперму путем изучения T-DMR. [112] Использование T-DMR оказалось полезным при идентификации различных биологических жидкостей, обнаруженных на местах преступлений. Исследователи в области судебной медицины в настоящее время ищут новые T-DMR в генах, которые можно использовать в качестве маркеров в судебно-медицинском анализе ДНК. DMR между раковыми и нормальными образцами (C-DMR) демонстрируют аберрантное метилирование при раке. [113] Хорошо известно, что метилирование ДНК связано с дифференцировкой и пролиферацией клеток. [114] Многие DMR были обнаружены на стадии разработки (D-DMR). [115] и в перепрограммированном прогрессе (R-DMR). [116] Кроме того, существуют внутрииндивидуальные DMR (Intra-DMR) с продольными изменениями глобального метилирования ДНК наряду с увеличением возраста у данного индивидуума. [117] Существуют также межиндивидуальные DMR (Inter-DMR) с разными паттернами метилирования у нескольких людей. [118]
QDMR (количественные дифференциально метилированные области) — это количественный подход для количественной оценки различий в метилировании и идентификации DMR на основе профилей метилирования по всему геному путем адаптации энтропии Шеннона. [119] Неплатформенный и бесвидовой характер QDMR делает его потенциально применимым для различных данных по метилированию. Этот подход обеспечивает эффективный инструмент для высокопроизводительной идентификации функциональных областей, участвующих в эпигенетической регуляции. QDMR можно использовать как эффективный инструмент для количественной оценки различий в метилировании и идентификации DMR в нескольких образцах. [120]
Было показано, что анализ набора генов (также известный как анализ путей; обычно используемые инструменты, такие как DAVID, GoSeq или GSEA) является серьезно необъективным при применении к данным высокопроизводительного метилирования (например, MeDIP-seq, MeDIP-ChIP, HELP-seq и т. д.). ), и поэтому в широком спектре исследований ошибочно сообщалось о гиперметилировании генов, связанных с развитием и дифференцировкой; Было высказано предположение, что это можно исправить с помощью перестановок меток образцов или использования статистической модели для контроля различий в количестве зондов CpG/сайтов CpG, нацеленных на каждый ген. [121]
ДНК метилирования Метки
Метки метилирования ДНК – участки генома с определенными закономерностями метилирования в определенном биологическом состоянии, например, в ткани, типе клеток, индивидууме – рассматриваются как возможные функциональные области, участвующие в регуляции транскрипции генов. Хотя разные типы клеток человека могут иметь один и тот же геном, эти клетки имеют разные метиломы. Систематическая идентификация и характеристика меток метилирования разных типов клеток имеют решающее значение для понимания сложной регуляторной сети, определяющей судьбу клеток. Хунбо Лю и др. предложил основанную на энтропии структуру под названием SMART для интеграции метиломов бисульфитного секвенирования всего генома в 42 тканях/клетках человека и идентифицировал 757 887 сегментов генома. [122] Почти 75% сегментов показали равномерное метилирование для всех типов клеток. Из оставшихся 25% сегментов они идентифицировали специфичные для типа клеток метки гипо/гиперметилирования, которые были специфически гипо/гиперметилированы в меньшинстве типов клеток, используя статистический подход, и представили атлас меток метилирования человека. Дальнейший анализ показал, что метки гипометилирования, специфичные для типа клеток, обогащались за счет сайтов связывания H3K27ac и транскрипционных факторов в зависимости от типа клеток. В частности, они заметили, что метки гипометилирования, специфичные для типа клеток, связаны со специфичными для типа клеток суперэнхансерами, которые управляют экспрессией генов клеточной идентичности. Эта структура обеспечивает дополнительную функциональную аннотацию генома человека и помогает выяснить критические особенности и функции гипометилирования, специфичного для типа клеток. [ нужна ссылка ]
Инструмент анализа и составления отчетов по специфическому метилированию на основе энтропии, получивший название «SMART», который фокусируется на интеграции большого количества метиломов ДНК для идентификации de novo меток метилирования, специфичных для типа клеток. Последняя версия SMART ориентирована на три основные функции, включая идентификацию de novo дифференциально метилированных регионов (DMR) путем сегментации генома, идентификацию DMR из заранее определенных интересующих областей и идентификацию дифференциально метилированных сайтов CpG. [123]
При идентификации и обнаружении жидкостей организма [ править ]
Метилирование ДНК позволяет анализировать несколько тканей за один анализ, а также идентифицировать небольшие количества жидкости организма с использованием выделенной ДНК. Обычно двумя подходами к метилированию ДНК являются либо чувствительные к метилированию ферменты рестрикции, либо обработка бисульфитом натрия. [124] Метилированные чувствительные ферменты рестрикции работают путем расщепления специфических CpG, цитозина и гуанина, разделенных только одной фосфатной группой, сайтов узнавания, когда CpG метилирован. Напротив, неметилированные цитозины трансформируются в урацил, а метилированные цитозины остаются метилированными. В частности, профили метилирования могут дать представление о том, когда и как биологические жидкости были оставлены на месте преступления, определить тип телесных жидкостей и приблизительный возраст, пол и фенотипические характеристики преступников. [125] Исследования указывают на различные маркеры, которые можно использовать для метилирования ДНК. Решение о том, какой маркер использовать для анализа, является одним из первых шагов идентификации жидкостей организма. Как правило, маркеры выбираются на основе анализа предыдущих исследований. Выбранные идентификационные маркеры должны давать положительный результат для одного типа клеток. Одной из частей хромосомы, которая является областью внимания при проведении метилирования ДНК, являются тканеспецифичные дифференциально метилированные области, T-DMR. Степень метилирования T-DMR варьируется в зависимости от жидкости организма. [125] Исследовательская группа разработала двойную систему маркеров. Первый маркер метилируется только в целевой жидкости, а второй — в остальных жидкостях. [108] Например, если маркер А венозной крови не метилирован, а маркер В венозной крови метилирован в жидкости, это указывает на наличие только венозной крови. Напротив, если маркер венозной крови A метилирован, а маркер венозной крови B не метилирован в некоторой жидкости, то это указывает на то, что венозная кровь находится в смеси жидкостей. Некоторыми примерами маркеров метилирования ДНК являются Mens1 (менструальная кровь), Spei1 (слюна) и Sperm2 (семенная жидкость).
Метилирование ДНК обеспечивает относительно хорошую чувствительность при идентификации и обнаружении жидкостей организма. В одном исследовании для подтверждения успешных результатов потребовалось всего десять нанограммов образца. [126] Метилирование ДНК обеспечивает хорошее различение смешанных образцов, поскольку оно включает в себя маркеры, которые подают сигналы «включено» или «выключено». Метилирование ДНК не является невосприимчивым к внешним условиям. Даже в условиях деградации с использованием методов метилирования ДНК маркеры достаточно стабильны, поэтому между деградированными и контрольными образцами все еще остаются заметные различия. В частности, в одном исследовании было обнаружено, что не было каких-либо заметных изменений в характере метилирования в течение длительного периода времени. [125]
Обнаружение метилирования ДНК во внеклеточной ДНК и других жидкостях организма в последнее время стало одним из основных подходов к жидкостной биопсии . [127] В частности, идентификация тканеспецифичных и специфических для заболевания моделей позволяет неинвазивно обнаруживать и контролировать такие заболевания, как рак. [128] По сравнению со строго геномными подходами к жидкой биопсии, профилирование метилирования ДНК предлагает большее количество дифференциально метилированных сайтов CpG и дифференциально метилированных областей (DMRS), что потенциально повышает его чувствительность. Алгоритмы деконволюции сигнала, основанные на метилировании ДНК, были успешно применены к бесклеточной ДНК и могут определять ткань происхождения рака неизвестной первичности, отторжения аллотрансплантата и устойчивости к гормональной терапии. [129]
Вычислительное предсказание [ править ]
Метилирование ДНК также можно обнаружить с помощью компьютерных моделей с помощью сложных алгоритмов и методов. Вычислительные модели могут облегчить глобальное профилирование метилирования ДНК в хромосомах, и зачастую такие модели выполняются быстрее и дешевле, чем биологические анализы. К таким современным вычислительным моделям относятся Bhasin, et al. , [130] Бок и др ., [131] и Чжэн и др . [132] [133] Вместе с биологическим анализом эти методы значительно облегчают анализ метилирования ДНК.
См. также [ править ]
- 5-гидроксиметилцитозин
- 5-метилцитозин
- 7-метилгуанозин
- Снижение метилирования ДНК I (DDM1) , гена метилирования растений.
- Деметилирующий агент
- Дифференциально метилированные области
- деметилирование ДНК
- перепрограммирование метилирования ДНК
- Эпигенетика , в которой метилирование ДНК вносит значительный вклад.
- Эпигенетические часы — метод расчета возраста на основе метилирования ДНК.
- Эпигеном
- Геном
- Геномный импринтинг , наследственная репрессия аллеля, основанная на метилировании ДНК.
- MethBase, База данных метилирования ДНК размещенная в браузере генома UCSC.
- MethDB База данных метилирования ДНК
- Н 6 -Метиладенозин
- Метилирование белка
Ссылки [ править ]
- ^ Д.Б. Данн, Дж.Д. Смит: «Наличие 6-метиламинопурина в дезоксирибонуклеиновых кислотах». В: Biochem J. 68(4), апрель 1958 г., S. 627–636. ПМИД 13522672. ПМК 1200409 .
- ^ B. F. Vanyushin, S. G. Tkacheva, A. N. Belozersky: "Rare bases in animal DNA". In: Nature . 225, 1970, S. 948–949. ПМИД 4391887 .
- ^ Мелани Эрлих, Мигель А. Гама-Соса, Лаура Х. Каррейра, Ларс Г. Юнгдал, Кеннет К. Куо, Чарльз В. Герке: «Метилирование ДНК у термофильных бактерий: N6-метилцитозин, 5-метилцитозин и N6-метиладенин ." В: Исследования нуклеиновых кислот . 13, 1985, с.1399. ПМИД 4000939. ПМК 341080 .
- ^ Jump up to: Перейти обратно: а б Эванс Х.Х., Эванс Т.Э. (декабрь 1970 г.). «Метилирование дезоксирибонуклеиновой кислоты Physarum polycephalum в различные периоды митотического цикла» . Журнал биологической химии . 245 (23): 6436–6441. дои : 10.1016/S0021-9258(18)62627-4 . ПМИД 5530731 .

- ^ Смит С.С., Ратер Д.И. (июль 1991 г.). «Отсутствие 5-метилцитозина в ДНК Dictyostelium discoideum» . Биохимический журнал . 277 (1): 273–275. дои : 10.1042/bj2770273 . ПМЦ 1151219 . ПМИД 1713034 .
- ^ Jump up to: Перейти обратно: а б Древелл Р.А., Кормье Т.С., Стинвик Дж.Л., Сен-Дени Дж., Табима Дж.Ф., Дреш Дж.М., Ларошель Д.А. (апрель 2023 г.). « В геноме Dictyostelium discoideum отсутствует значительное метилирование ДНК, и палиндромные последовательности обнаруживаются как источник ложноположительных результатов при бисульфитном секвенировании» . НАР Геномика Биоинформатика . 5 (2): lqad035. дои : 10.1093/nargab/lqad035 . ПМЦ 10111430 . ПМИД 37081864 .
- ^ Ху CW, Чен JL, Сюй YW, Йен CC, Чао MR (январь 2015 г.). «Анализ следов метилированных и гидроксиметилированных цитозинов в ДНК методом ЖХ-МС/МС с изотопным разбавлением: первые доказательства метилирования ДНК у Caenorhabditis elegans». Биохимический журнал . 465 (1): 39–47. дои : 10.1042/bj20140844 . ПМИД 25299492 .
- ^ Птица А (декабрь 2001 г.). «Молекулярная биология. Разговор о метилировании между гистонами и ДНК». Научный компас. Наука . 294 (5549): 2113–2115. дои : 10.1126/science.1066726 . hdl : 1842/464 . ПМИД 11739943 . S2CID 82947750 .
В результате этого процесса, известного как точечная мутация, индуцированная повторами (RIP), геном Neurospora дикого типа содержит небольшую фракцию метилированной ДНК, при этом большая часть ДНК остается неметилированной.
- ^ Jump up to: Перейти обратно: а б с Капуано Ф., Мюллер М., Кок Р., Блом Х.Дж., Ральсер М. (апрель 2014 г.). «Метилирование ДНК цитозина обнаружено у Drosophila melanogaster, но отсутствует у Saccharomyces cerevisiae, Schizosaccharomyces pombe и других видов дрожжей» . Аналитическая химия . 86 (8): 3697–3702. дои : 10.1021/ac500447w . ПМК 4006885 . ПМИД 24640988 .
- ^ Рател Д., Раванат Дж.Л., Бергер Ф., Вион Д. (март 2006 г.). «N6-метиладенин: другое метилированное основание ДНК» . Биоэссе . 28 (3): 309–315. doi : 10.1002/bies.20342 . ПМК 2754416 . ПМИД 16479578 .
- ^ Ву Т.П., Ван Т., Сетин М.Г., Лай Ю., Чжу С., Лин К. и др. (апрель 2016 г.). «Метилирование ДНК N (6)-аденина в эмбриональных стволовых клетках млекопитающих» . Природа . 532 (7599): 329–333. Бибкод : 2016Natur.532..329W . дои : 10.1038/nature17640 . ПМЦ 4977844 . ПМИД 27027282 .
- ^ Анжела Бекеши и Беата Дж. Вертесси «Урацил в ДНК: ошибка или сигнал?»
- ^ Рана АК, Анкри С (2016). «Возрождение мира РНК: взгляд на появление РНК-метилтрансфераз» . Границы генетики . 7:99 . дои : 10.3389/fgene.2016.00099 . ПМЦ 4893491 . ПМИД 27375676 .
- ^ Додж Дж. Э., Рамсахой Б. Х., Во З. Г., Окано М., Ли Э (май 2002 г.). «Метилирование de novo провируса MMLV в эмбриональных стволовых клетках: метилирование CpG и не-CpG». Джин . 289 (1–2): 41–48. дои : 10.1016/S0378-1119(02)00469-9 . ПМИД 12036582 .
- ^ Хейнс Т.Р., Роденхайзер Д.И., Эйнсворт П.Дж. (декабрь 2001 г.). «Аллель-специфическое не-CpG-метилирование гена Nf1 на ранних стадиях развития мышей» . Биология развития . 240 (2): 585–598. дои : 10.1006/dbio.2001.0504 . ПМИД 11784085 .
- ^ Jump up to: Перейти обратно: а б Листер Р., Пелиццола М., Доуэн Р.Х., Хокинс Р.Д., Хон Дж., Тонти-Филиппини Дж. и др. (ноябрь 2009 г.). «Метиломы ДНК человека при разрешении оснований демонстрируют широко распространенные эпигеномные различия» . Природа . 462 (7271): 315–322. Бибкод : 2009Natur.462..315L . дои : 10.1038/nature08514 . ПМК 2857523 . ПМИД 19829295 .
- ^ Листер Р., Мукамель Э.А., Нери Дж.Р., Урих М., Паддифут К.А., Джонсон Н.Д. и др. (август 2013 г.). «Глобальная эпигеномная реконфигурация во время развития мозга млекопитающих» . Наука . 341 (6146): 1237905. doi : 10.1126/science.1237905 . ПМК 3785061 . ПМИД 23828890 .
- ^ Jump up to: Перейти обратно: а б Кулис М., Меркель А., Хит С., Кейрос А.С., Шайлер Р.П., Кастеллано Г. и др. (июль 2015 г.). «Полногеномный отпечаток метилома ДНК во время дифференцировки В-клеток человека» . Природная генетика . 47 (7): 746–756. дои : 10.1038/ng.3291 . ПМЦ 5444519 . ПМИД 26053498 .
- ^ Тост Дж. (январь 2010 г.). «Метилирование ДНК: введение в биологию и связанные с болезнями изменения многообещающего биомаркера». Молекулярная биотехнология . 44 (1): 71–81. дои : 10.1007/s12033-009-9216-2 . ПМИД 19842073 . S2CID 20307488 .
- ^ Стадлер М.Б., Мурр Р., Бургер Л., Иванек Р., Линерт Ф., Шелер А. и др. (декабрь 2011 г.). «ДНК-связывающие факторы формируют метилом мыши в дистальных регуляторных областях» . Природа . 480 (7378): 490–495. дои : 10.1038/nature11086 . ПМИД 22170606 .
- ^ Jump up to: Перейти обратно: а б с Земах А., МакДэниел И.Е., Сильва П., Зильберман Д. (май 2010 г.). «Полногеномный эволюционный анализ метилирования ДНК эукариот» . Наука (отчет ScienceExpress). 328 (5980): 916–919. Бибкод : 2010Sci...328..916Z . дои : 10.1126/science.1186366 . ПМИД 20395474 . S2CID 206525166 .
Здесь мы количественно оцениваем метилирование ДНК в семнадцати геномах эукариот....
Дополнительные данные, судя по всему, доступны только через платный доступ science.sciencemag.org. - ^ Сузуки М.М., Керр А.Р., Де Соуза Д., Берд А. (май 2007 г.). «Метилирование CpG направлено на единицы транскрипции в геноме беспозвоночных» . Геномные исследования . 17 (5): 625–631. дои : 10.1101/гр.6163007 . ПМЦ 1855171 . ПМИД 17420183 .
- ^ Jump up to: Перейти обратно: а б Ландер Э.С., Линтон Л.М., Биррен Б., Нусбаум С., Зоди М.К., Болдуин Дж. и др. (февраль 2001 г.). «Первичное секвенирование и анализ генома человека» . Природа . 409 (6822): 860–921. Бибкод : 2001Natur.409..860L . дои : 10.1038/35057062 . hdl : 2027.42/62798 . ПМИД 11237011 .
- ^ Берд АП (15 мая 1986 г.). «Островки, богатые CpG, и функция метилирования ДНК». Природа . 321 (6067): 209–213. Бибкод : 1986Natur.321..209B . дои : 10.1038/321209a0 . ПМИД 2423876 . S2CID 4236677 .
- ^ Гардинер-Гарден М., Фроммер М. (июль 1987 г.). «CpG-островки в геномах позвоночных». Журнал молекулярной биологии . 196 (2): 261–282. дои : 10.1016/0022-2836(87)90689-9 . ПМИД 3656447 .
- ^ Иллингворт Р.С., Грюневальд-Шнайдер У., Уэбб С., Керр А.Р., Джеймс К.Д., Тернер Д.Д. и др. (сентябрь 2010 г.). «Островки-сироты CpG идентифицируют многочисленные консервативные промоторы в геноме млекопитающих» . ПЛОС Генетика . 6 (9): e1001134. дои : 10.1371/journal.pgen.1001134 . ПМЦ 2944787 . ПМИД 20885785 .
- ^ Саксонов С., Берг П., Брютлаг Д.Л. (январь 2006 г.). «Полногеномный анализ динуклеотидов CpG в геноме человека позволяет выделить два различных класса промоторов» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (5): 1412–1417. Бибкод : 2006PNAS..103.1412S . дои : 10.1073/pnas.0510310103 . ПМЦ 1345710 . ПМИД 16432200 .
- ^ Jump up to: Перейти обратно: а б Фэн С., Кокус С.Дж., Чжан X, Чен П.Ю., Бостик М., Голл М.Г. и др. (май 2010 г.). «Сохранение и расхождение паттернов метилирования у растений и животных» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (19): 8689–8694. дои : 10.1073/pnas.1002720107 . ПМЦ 2889301 . ПМИД 20395551 .
- ^ Мон Ф., Вебер М., Ребхан М., Ролофф Т.С., Рихтер Дж., Стадлер М.Б. и др. (июнь 2008 г.). «Специфические для линии мишени полисот и метилирование ДНК de novo определяют ограничение и потенциал предшественников нейронов» . Молекулярная клетка . 30 (6): 755–766. doi : 10.1016/j.molcel.2008.05.007 . ПМИД 18514006 .
- ^ Вебер М., Хеллманн И., Стадлер М.Б., Рамос Л., Паабо С., Ребхан М., Шубелер Д. (апрель 2007 г.). «Распространение, потенциал молчания и эволюционное влияние метилирования ДНК промотора в геноме человека». Природная генетика . 39 (4): 457–466. дои : 10.1038/ng1990 . ПМИД 17334365 . S2CID 22446734 .
- ^ Шюбелер Д. (январь 2015 г.). «Функция и информативность метилирования ДНК». Природа . 517 (7534): 321–326. Бибкод : 2015Natur.517..321S . дои : 10.1038/nature14192 . ПМИД 25592537 . S2CID 4403755 .
- ^ Чой М.К., Мовассаг М., Го Х.Г., Беннетт М.Р., Даун Т.А., Фу Р.С. (сентябрь 2010 г.). «Консервативные по всему геному консенсусные мотивы связывания факторов транскрипции гиперметилированы» . БМК Геномика . 11 (1): 519. дои : 10.1186/1471-2164-11-519 . ПМК 2997012 . ПМИД 20875111 .
- ^ Далет Т., Аргуэсо Лерида А., Аль Адхами Х., Дюма М., Бендер А., Нгондо Р.П. и др. (июнь 2020 г.). «Полногеномный анализ эмбрионов мышей показывает важность метилирования ДНК для целостности транскрипции» . Природные коммуникации . 11 (1): 3153. Бибкод : 2020NatCo..11.3153D . дои : 10.1038/s41467-020-16919-w . ПМК 7305168 . ПМИД 32561758 .
- ^ Хафф Дж.Т., Зильберман Д. (март 2014 г.). «Независимое от Dnmt1 метилирование CG способствует позиционированию нуклеосом у различных эукариот» . Клетка . 156 (6): 1286–1297. дои : 10.1016/j.cell.2014.01.029 . ПМЦ 3969382 . ПМИД 24630728 .
- ^ Йодер Дж.А., Уолш К.П., Бестор Т.Х. (август 1997 г.). «Метилирование цитозина и экология внутригеномных паразитов» . Тенденции в генетике . 13 (8): 335–340. дои : 10.1016/s0168-9525(97)01181-5 . ПМИД 9260521 .
- ^ Чжоу В., Лян Г., Моллой П.Л., Джонс П.А. (август 2020 г.). «Метилирование ДНК обеспечивает расширение генома за счет мобильных элементов» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (32): 19359–19366. Бибкод : 2020PNAS..11719359Z . дои : 10.1073/pnas.1921719117 . ПМК 7431005 . ПМИД 32719115 .
- ^ Лев Маор Г, Йеарим А, Аст Г (май 2015 г.). «Альтернативная роль метилирования ДНК в регуляции сплайсинга». Тенденции в генетике . 31 (5): 274–280. дои : 10.1016/j.tig.2015.03.002 . ПМИД 25837375 . S2CID 34258335 .
- ^ Маунакеа А.К., Нагараджан Р.П., Биленки М., Баллинджер Т.Дж., Д'Суза С., Фаус С.Д. и др. (июль 2010 г.). «Консервативная роль внутригенного метилирования ДНК в регуляции альтернативных промоторов» . Природа . 466 (7303): 253–257. Бибкод : 2010Natur.466..253M . дои : 10.1038/nature09165 . ПМЦ 3998662 . ПМИД 20613842 .
- ^ Карроцца М.Дж., Ли Б., Флоренс Л., Суганума Т., Суонсон С.К., Ли К.К. и др. (ноябрь 2005 г.). «Метилирование гистона H3 с помощью Set2 направляет деацетилирование кодирующих областей с помощью Rpd3S для подавления ложной внутригенной транскрипции» . Клетка . 123 (4): 581–592. дои : 10.1016/j.cell.2005.10.023 . ПМИД 16286007 . S2CID 9328002 .
- ^ Кедр Х., Бергман Ю. (июль 2012 г.). «Программирование закономерностей метилирования ДНК» . Ежегодный обзор биохимии . 81 : 97–117. doi : 10.1146/annurev-biochem-052610-091920 . ПМИД 22404632 . – через Ежегодные обзоры (требуется подписка)
- ^ Jump up to: Перейти обратно: а б Мужич Радович В., Буноза П., Марич Т., Химельрайх-Перич М., Булич-Якуш Ф., Такахаши М. и др. (июль 2022 г.). «Глобальное метилирование ДНК и хондрогенез зачатков конечностей крысы в трехмерной системе культуры органов» . Боснийский журнал фундаментальных медицинских наук . 22 (4): 560–568. дои : 10.17305/bjbms.2021.6584 . ПМЦ 9392980 . ПМИД 35188093 . S2CID 247010798 .
- ^ Бирд С., Ли Э., Джениш Р. (октябрь 1995 г.). «Потеря метилирования активирует Xist в соматических, но не в эмбриональных клетках» . Гены и развитие . 9 (19): 2325–2334. дои : 10.1101/gad.9.19.2325 . ПМИД 7557385 .
- ^ Ли Э., Бирд С., Джениш Р. (ноябрь 1993 г.). «Роль метилирования ДНК в геномном импринтинге». Природа . 366 (6453): 362–365. Бибкод : 1993Natur.366..362L . дои : 10.1038/366362a0 . ПМИД 8247133 . S2CID 4311091 .
- ^ Боргель Дж., Гиберт С., Ли Ю., Чиба Х., Шубелер Д., Сасаки Х. и др. (декабрь 2010 г.). «Цели и динамика метилирования ДНК промотора на раннем этапе развития мышей». Природная генетика . 42 (12): 1093–1100. дои : 10.1038/ng.708 . ПМИД 21057502 . S2CID 205357042 .
- ^ Зайзенбергер С., Пит Дж.Р., Хор Т.А., Сантос Ф., Дин В., Рейк В. (январь 2013 г.). «Перепрограммирование метилирования ДНК в жизненном цикле млекопитающих: построение и преодоление эпигенетических барьеров» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 368 (1609): 20110330. doi : 10.1098/rstb.2011.0330 . ПМЦ 3539359 . ПМИД 23166394 .
- ^ Ван Ю.П., Лэй Цюй (май 2018 г.). «Метаболическая перекодировка эпигенетики рака» . Раковые коммуникации . 38 (1): 25. дои : 10.1186/s40880-018-0302-3 . ПМЦ 5993135 . ПМИД 29784032 .
- ^ Даура-Оллер Э., Кабре М., Монтеро М.А., Патернэн Дж.Л., Ромеу А. (апрель 2009 г.). «Специфическое гипометилирование генов и рак: новое понимание тенденций особенностей кодирующих регионов» . Биоинформация . 3 (8): 340–343. дои : 10.6026/97320630003340 . ПМК 2720671 . ПМИД 19707296 .
- ^ Крейг Дж. М., Вонг, Северная Каролина, ред. (2011). Эпигенетика: Справочное руководство . Кайстер Академик Пресс . ISBN 978-1-904455-88-2 .
- ^ Jump up to: Перейти обратно: а б с Гонсало С. (август 2010 г.). «Эпигенетические изменения при старении» . Журнал прикладной физиологии . 109 (2): 586–597. doi : 10.1152/japplphysicalol.00238.2010 . ПМЦ 2928596 . ПМИД 20448029 .
- ^ Лунд Г., Андерссон Л., Лаурия М., Линдхольм М., Фрага М.Ф., Вильяр-Гареа А. и др. (июль 2004 г.). «Полиморфизмы метилирования ДНК предшествуют любым гистологическим признакам атеросклероза у мышей, лишенных аполипопротеина Е» . Журнал биологической химии . 279 (28): 29147–29154. дои : 10.1074/jbc.m403618200 . hdl : 2445/176763 . ПМИД 15131116 .
- ^ Кастро Р., Ривера И., Струйс Э.А., Янсен Э.Э., Раваско П., Камило М.Е. и др. (август 2003 г.). «Повышение концентрации гомоцистеина и S-аденозилгомоцистеина и гипометилирование ДНК при сосудистых заболеваниях» . Клиническая химия . 49 (8): 1292–1296. дои : 10.1373/49.8.1292 . ПМИД 12881445 .
- ^ Хуан Ю.С., Чжи Ю.Ф., Ван С.Р. (октябрь 2009 г.). «Гиперметилирование гена альфа-рецептора эстрогена у пациентов с атероматозом и его корреляция с гомоцистеином». Патофизиология . 16 (4): 259–265. doi : 10.1016/j.pathophys.2009.02.010 . ПМИД 19285843 .
- ^ Донг С., Юн В., Гольдшмидт-Клермонт П.Дж. (август 2002 г.). «Метилирование ДНК и атеросклероз» . Журнал питания . 132 (8 дополнений): 2406S–2409S. дои : 10.1093/jn/132.8.2406S . ПМИД 12163701 .
- ^ Инь А.К., Хассанаин Х.Х., Роос К.М., Смираглия Д.Д., Исса Дж.Дж., Михлер Р.Э. и др. (апрель 2000 г.). «Метилирование промотора гена эстрогенового рецептора-альфа избирательно увеличивается в пролиферирующих гладкомышечных клетках аорты человека» . Сердечно-сосудистые исследования . 46 (1): 172–179. дои : 10.1016/s0008-6363(00)00004-3 . ПМИД 10727665 .
- ^ Чжу С., Гольдшмидт-Клермон П.Дж., Донг К. (август 2005 г.). «Инактивация монокарбоксилатного транспортера MCT3 путем метилирования ДНК при атеросклерозе» . Тираж . 112 (9): 1353–1361. дои : 10.1161/circulationaha.104.519025 . ПМИД 16116050 .
- ^ Пепин М.Е., Ха СМ, Кроссман Д.К., Литовский С.Х., Варамбалли С., Барчу Дж.П. и др. (март 2019 г.). «Полногеномное метилирование ДНК кодирует перепрограммирование сердечной транскрипции при ишемической сердечной недостаточности у человека» . Лабораторные исследования; Журнал технических методов и патологии . 99 (3): 371–386. дои : 10.1038/s41374-018-0104-x . ПМК 6515060 . ПМИД 30089854 .
- ^ Пепин М.Е., Ха С.М., Поттер Л.А., Бакши С., Барчу Дж.П., Хадж Асаад А. и др. (май 2021 г.). «Расовое и социально-экономическое неравенство связано с различиями в метилировании сердечной ДНК среди мужчин с терминальной стадией сердечной недостаточности» . Американский журнал физиологии. Физиология сердца и кровообращения . 320 (5): H2066–H2079. дои : 10.1152/ajpheart.00036.2021 . ПМК 8163657 . ПМИД 33769919 .
- ^ Хорват С (2013). «Возраст метилирования ДНК тканей и типов клеток человека» . Геномная биология . 14 (10): 115 р. дои : 10.1186/gb-2013-14-10-r115 . ПМК 4015143 . ПМИД 24138928 . (Ошибка: два : 10.1186/s13059-015-0649-6 , PMID 25968125 , Часы втягивания )
- ^ Вонг CC, Каспи А., Уильямс Б., Крейг И.В., Хаутс Р., Эмблер А. и др. (август 2010 г.). «Продольное исследование эпигенетических вариаций у близнецов» . Эпигенетика . 5 (6): 516–526. дои : 10.4161/epi.5.6.12226 . ПМЦ 3322496 . ПМИД 20505345 .
- ^ Jump up to: Перейти обратно: а б Хейн Х., Ли Н., Феррейра Х.Дж., Моран С., Пизано Д.Г., Гомес А. и др. (июнь 2012 г.). «Различные метиломы ДНК новорожденных и долгожителей» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (26): 10522–10527. Бибкод : 2012PNAS..10910522H . дои : 10.1073/pnas.1120658109 . ПМК 3387108 . ПМИД 22689993 .
- ^ Jump up to: Перейти обратно: а б Баррес Р., Ян Дж., Иган Б., Трибак Дж.Т., Расмуссен М., Фриц Т. и др. (март 2012 г.). «Острые физические нагрузки изменяют метилирование промотора в скелетных мышцах человека» . Клеточный метаболизм . 15 (3): 405–411. дои : 10.1016/j.cmet.2012.01.001 . ПМИД 22405075 .
- ^ Рённ Т., Волков П., Давегард С., Дайе Т., Холл Е., Олссон А.Х. и др. (июнь 2013 г.). «Шестимесячное занятие физическими упражнениями влияет на полногеномный характер метилирования ДНК в жировой ткани человека» . ПЛОС Генетика . 9 (6): e1003572. дои : 10.1371/journal.pgen.1003572 . ПМЦ 3694844 . ПМИД 23825961 .
- ^ Чжан Ф.Ф., Кардарелли Р., Кэрролл Дж., Чжан С., Фульда К.Г., Гонсалес К. и др. (март 2011 г.). «Физическая активность и глобальное метилирование геномной ДНК в популяции, свободной от рака» . Эпигенетика . 6 (3): 293–299. дои : 10.4161/epi.6.3.14378 . ПМК 3092677 . ПМИД 21178401 .
- ^ Jump up to: Перейти обратно: а б Суэтт Джей Ди (май 2016 г.). «Динамическое метилирование ДНК контролирует перемещение глутаматных рецепторов и масштабирование синапсов» . Журнал нейрохимии . 137 (3): 312–330. дои : 10.1111/jnc.13564 . ПМЦ 4836967 . ПМИД 26849493 .
- ^ Ким С., Каанг Б.К. (январь 2017 г.). «Эпигенетическая регуляция и ремоделирование хроматина в обучении и памяти» . Экспериментальная и молекулярная медицина . 49 (1): e281. дои : 10.1038/emm.2016.140 . ПМК 5291841 . ПМИД 28082740 .
- ^ Шафе Г.Е., Надель Н.В., Салливан Г.М., Харрис А., Леду Дж.Е. (1999). «Консолидация памяти для контекстуального и слухового формирования страха зависит от синтеза белка, PKA и киназы MAP» . Обучение и память . 6 (2): 97–110. дои : 10.1101/lm.6.2.97 . ПМК 311283 . ПМИД 10327235 .
- ^ Jump up to: Перейти обратно: а б с Хальдер Р., Хеннион М., Видал Р.О., Шомрони О., Рахман Р.У., Раджпут А. и др. (январь 2016 г.). «Изменения метилирования ДНК в генах пластичности сопровождают формирование и поддержание памяти» . Природная неврология . 19 (1): 102–110. дои : 10.1038/nn.4194 . ПМК 4700510 . ПМИД 26656643 .
- ^ Jump up to: Перейти обратно: а б Дюк К.Г., Кеннеди А.Дж., Гэвин К.Ф., Дэй Дж.Дж., Суэтт Дж.Д. (июль 2017 г.). «Эпигеномная реорганизация в гиппокампе, зависящая от опыта» . Обучение и память . 24 (7): 278–288. дои : 10.1101/lm.045112.117 . ПМК 5473107 . ПМИД 28620075 .
- ^ Jump up to: Перейти обратно: а б Бернштейн С (2022). «Метилирование ДНК и установление памяти» . Эпигенетические выводы . 15 : 25168657211072499. дои : 10.1177/25168657211072499 . ПМЦ 8793415 . ПМИД 35098021 .
- ^ Грачев А. «Обзор по метилированию ДНК» . МЕТОДЫ.info .
- ^ Барау Дж., Тейсандье А., Самудио Н., Рой С., Налессо В., Эро И. и др. (ноябрь 2016 г.). «ДНК-метилтрансфераза DNMT3C защищает мужские половые клетки от активности транспозонов». Наука . 354 (6314): 909–912. Бибкод : 2016Sci...354..909B . дои : 10.1126/science.aah5143 . ПМИД 27856912 . S2CID 30907442 .
- ^ Джайн Д., Мейдан С., Ланге Дж., Клейс Буарт С., Лайллер Н., Мейсон С.Э. и др. (август 2017 г.). «rahu представляет собой мутантный аллель Dnmt3c, кодирующий гомолог ДНК-метилтрансферазы, необходимый для мейоза и репрессии транспозонов в зародышевой линии мышей-самцов» . ПЛОС Генетика . 13 (8): e1006964. дои : 10.1371/journal.pgen.1006964 . ПМК 5607212 . ПМИД 28854222 .
- ^ Голл М.Г., Кирпекар Ф., Маггерт К.А., Йодер Дж.А., Се К.Л., Чжан X и др. (январь 2006 г.). «Метилирование тРНКАсп гомологом ДНК-метилтрансферазы Dnmt2». Наука . 311 (5759): 395–398. Бибкод : 2006Sci...311..395G . дои : 10.1126/science.1120976 . ПМИД 16424344 . S2CID 39089541 .
- ^ Jump up to: Перейти обратно: а б Чжан Х., Лан З., Чжу Дж. К. (август 2018 г.). «Динамика и функция метилирования ДНК у растений». Обзоры природы. Молекулярно-клеточная биология . 19 (8): 489–506. дои : 10.1038/s41580-018-0016-z . ПМИД 29784956 . S2CID 256745172 .
- ^ Дубин М.Дж., Чжан П., Мэн Д., Ремигеро М.С., Осборн Э.Дж., Паоло Казале Ф. и др. (май 2015 г.). Дермицакис ET (ред.). «Метилирование ДНК у арабидопсиса имеет генетическую основу и свидетельствует о местной адаптации» . электронная жизнь . 4 : e05255. doi : 10.7554/eLife.05255 . ПМЦ 4413256 . ПМИД 25939354 .
- ^ Цао X, Якобсен С.Е. (декабрь 2002 г.). «Локус-специфический контроль асимметричного метилирования и метилирования CpNpG с помощью генов метилтрансферазы DRM и CMT3» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (Приложение 4): 16491–16498. Бибкод : 2002PNAS...9916491C . дои : 10.1073/pnas.162371599 . ПМК 139913 . ПМИД 12151602 .
- ^ Нидерхут С.Э., Бьюик А.Дж., Джи Л., Алабади М.С., Ким К.Д., Ли Кью и др. (сентябрь 2016 г.). «Широко распространенное естественное изменение метилирования ДНК у покрытосеменных растений» . Геномная биология . 17 (1): 194. дои : 10.1186/s13059-016-1059-0 . ПМК 5037628 . ПМИД 27671052 .
- ^ Земах А., МакДэниел И.Е., Сильва П., Зильберман Д. (май 2010 г.). «Полногеномный эволюционный анализ метилирования ДНК эукариот» . Наука . 328 (5980): 916–919. Бибкод : 2010Sci...328..916Z . дои : 10.1126/science.1186366 . ПМИД 20395474 . S2CID 206525166 .
- ^ Jump up to: Перейти обратно: а б Ауфзац В., Метте М.Ф., ван дер Винден Дж., Мацке А.Дж., Мацке М. (декабрь 2002 г.). «РНК-направленное метилирование ДНК у Arabidopsis» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (Приложение 4): 16499–16506. Бибкод : 2002PNAS...9916499A . дои : 10.1073/pnas.162371499 . ПМК 139914 . ПМИД 12169664 .
- ^ Бьюик А.Дж., Фогель К.Дж., Мур А.Дж., Шмитц Р.Дж. (март 2017 г.). «Эволюция метилирования ДНК у насекомых» . Молекулярная биология и эволюция . 34 (3): 654–665. дои : 10.1093/molbev/msw264 . ПМК 5400375 . ПМИД 28025279 .
- ^ Ван Ю., Джорда М., Джонс П.Л., Малешка Р., Линг Х., Робертсон Х.М. и др. (октябрь 2006 г.). «Функциональная система метилирования CpG у общественного насекомого». Наука . 314 (5799): 645–647. Бибкод : 2006Sci...314..645W . дои : 10.1126/science.1135213 . ПМИД 17068262 . S2CID 31709665 .
- ^ Ин и Ли-Бьярлай (2015). Физиологические и молекулярные механизмы питания медоносных пчел . Достижения в физиологии насекомых. Том. 49. стр. 25–58. дои : 10.1016/bs.aiip.2015.06.002 . ISBN 9780128025864 .
- ^ Ли-Бьярлей Х., Ли Ю., Страуд Х., Фэн С., Ньюман Т.С., Канеда М. и др. (июль 2013 г.). «Нокаут ДНК-метилтрансферазы 3 РНК-интерференцией влияет на альтернативный сплайсинг генов у медоносной пчелы» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (31): 12750–12755. Бибкод : 2013PNAS..11012750L . дои : 10.1073/pnas.1310735110 . ПМЦ 3732956 . ПМИД 23852726 .
- ^ Смит С.С., Томас Калифорния (май 1981 г.). «Двумерный рестрикционный анализ ДНК дрозофилы: самцов и самок». Джин . 13 (4): 395–408. дои : 10.1016/0378-1119(81)90019-6 . ПМИД 6266924 .
- ^ Лико Ф., Рамсахой Б.Х., Джениш Р. (ноябрь 2000 г.). «Метилирование ДНК у Drosophila melanogaster». Природа . 408 (6812): 538–540. дои : 10.1038/35046205 . ПМИД 11117732 . S2CID 4427540 .
- ^ Такаяма С., Дахби Дж., Робертс А., Мао Г., Хио С.Дж., Пахтер Л. и др. (май 2014 г.). «Метилирование генома у D. melanogaster обнаруживается в специфических коротких мотивах и не зависит от активности DNMT2» . Геномные исследования . 24 (5): 821–830. дои : 10.1101/гр.162412.113 . ПМК 4009611 . ПМИД 24558263 .
- ^ Чжан Г., Хуан Х., Лю Д., Ченг Ю., Лю С., Чжан В. и др. (май 2015 г.). «Модификация ДНК N6-метиладенина у дрозофилы» . Клетка . 161 (4): 893–906. дои : 10.1016/j.cell.2015.04.018 . ПМИД 25936838 .
- ^ Антекера Ф, Тамаме М, Вильянуэва-младший, Сантос Т (июль 1984 г.). «Метилирование ДНК у грибов» . Журнал биологической химии . 259 (13): 8033–8036. дои : 10.1016/S0021-9258(17)39681-3 . ПМИД 6330093 .
- ^ Бинц Т., Д'Мелло Н., Хорген П.А. (1998). «Сравнение уровней метилирования ДНК у избранных изолятов высших грибов». Микология . 90 (5): 785–790. дои : 10.2307/3761319 . JSTOR 3761319 .
- ^ Лю С.Ю., Линь Дж.К., Ву Х.Л., Ван CC, Хуан С.Дж., Луо Ю.Ф. и др. (2012). «Бисульфитное секвенирование показывает, что Aspergillus flavus имеет пробел в метилировании ДНК» . ПЛОС ОДИН . 7 (1): e30349. Бибкод : 2012PLoSO...730349L . дои : 10.1371/journal.pone.0030349 . ПМК 3262820 . ПМИД 22276181 .
- ^ Селкер ЕС, Тунтас Н.А., Кросс С.Х., Марголин Б.С., Мерфи Дж.Г., Берд А.П., Фрейтаг М. (апрель 2003 г.). «Метилированный компонент генома Neurospora crassa». Природа . 422 (6934): 893–897. Бибкод : 2003Natur.422..893S . дои : 10.1038/nature01564 . hdl : 1842/694 . ПМИД 12712205 . S2CID 4380222 .
- ^ Смит С.С., Ратнер Д.И. (июль 1991 г.). «Отсутствие 5-метилцитозина в ДНК Dictyostelium discoideum» . Биохимический журнал . 277 (Часть 1) (Часть 1): 273–275. дои : 10.1042/bj2770273 . ПМЦ 1151219 . ПМИД 1713034 .
- ^ Рейли Дж.Г., Браун Р., Томас К.А. (июль 1980 г.). «Метилирование в ДНК Physarum». Письма ФЭБС . 116 (2): 181–184. дои : 10.1016/0014-5793(80)80638-7 . ПМИД 6250882 . S2CID 83941623 .
- ^ Мэтью Дж. Блоу, Тайсон А. Кларк, Крис Г. Даум, Адам М. Дойчбауэр, Алексей Фоменков, Роксана Фрайс, Джефф Фрула, Донван Д. Канг, Рекс Р. Мальмстрем, Ричард Д. Морган, Янош Посфаи, Канвар Сингх , Аксель Визель, Келли Ветмор, Чжиин Чжао, Эдвард М. Рубин, Джонас Корлах, Лен А. Пеннаккио, Ричард Дж. Робертс: Эпигеномный ландшафт прокариотов. В: PLoS Genet. 12(2), февраль 2016 г., S. e1005854. doi:10.1371/journal.pgen.1005854. ПМИД 26870957. ПМК 4752239
- ^ Оливейра, PH (август 2021 г.). «Бактериальная эпигеномика: достижение совершеннолетия» . mSystems . 6 (4): e0074721. doi : 10.1128/mSystems.00747-21 . ПМЦ 8407109 . ПМИД 34402642 .
- ^ Оливейра П.Х., Рибис Дж.В., Гарретт Э.М., Трзилова Д., Ким А., Секулович О. и др. (январь 2020 г.). «Эпигеномная характеристика Clostridioides difficile выявила консервативную ДНК-метилтрансферазу, которая опосредует споруляцию и патогенез» . Природная микробиология . 5 (1): 166–180. дои : 10.1038/s41564-019-0613-4 . ПМЦ 6925328 . ПМИД 31768029 .
- ^ Палмер Б.Р., Маринус М.Г. (май 1994 г.). «Штаммы Escherichia coli dam и dcm - обзор». Джин . 143 (1): 1–12. дои : 10.1016/0378-1119(94)90597-5 . ПМИД 8200522 .
- ^ «Создание неметилированной (dam-/dcm-) ДНК» . Архивировано из оригинала 6 января 2011 г.
- ^ Рана АК (январь 2018 г.). «Расследование преступлений посредством анализа метилирования ДНК: методы и применение в криминалистике» . Египетский журнал судебной медицины . 8 (7). дои : 10.1186/s41935-018-0042-1 .
- ^ Эрнандес Х.Г., Це М.Ю., Панг С.К., Арболеда Х., Фореро Д.А. (октябрь 2013 г.). «Оптимизация методологии анализа метилирования ДНК на основе ПЦР» . БиоТехники . 55 (4): 181–197. дои : 10.2144/000114087 . ПМИД 24107250 .
- ^ Фэн, Сухуа; Чжун, Чжэньхуэй; Ван, Мин; Якобсен, Стивен Э. (07 октября 2020 г.). «Эффективное и точное определение закономерностей метилирования ДНК по всему геному у Arabidopsis thaliana с помощью ферментативного секвенирования метила» . Эпигенетика и хроматин . 13 (1): 42. дои : 10.1186/s13072-020-00361-9 . ISSN 1756-8935 . ПМЦ 7542392 . ПМИД 33028374 .
- ^ Вуд Р.Дж., Мейнард-Смит, доктор медицинских наук, Робинсон В.Л., Ойстон ПК, Титболл Р.В., Роуч П.Л. (август 2007 г.). Фугманн С. (ред.). «Кинетический анализ активности ДНК-аденин-метилтрансферазы Yersinia pestis с использованием полуметилированного молекулярного разрывающегося легкого олигонуклеотида» . ПЛОС ОДИН . 2 (8): е801. Бибкод : 2007PLoSO...2..801W . дои : 10.1371/journal.pone.0000801 . ЧВК 1949145 . ПМИД 17726531 .
- ^ Ли Дж., Ян Х., Ван К., Тан В., Чжоу Икс (февраль 2007 г.). «Шпилька-флуоресцентный ДНК-зонд для мониторинга метилирования ДНК в реальном времени». Аналитическая химия . 79 (3): 1050–1056. дои : 10.1021/ac061694i . ПМИД 17263334 .
- ^ Войдач Т.К., Добрович А. (2007). «Плавление с высоким разрешением, чувствительное к метилированию (MS-HRM): новый подход к чувствительной и высокопроизводительной оценке метилирования» . Исследования нуклеиновых кислот . 35 (6): е41. дои : 10.1093/нар/gkm013 . ПМЦ 1874596 . ПМИД 17289753 .
- ^ Малентакки Ф, Форни Дж, Винчи С, Орландо К (июль 2009 г.). «Количественная оценка метилирования ДНК путем оптимизации протокола дифференциального анализа расплава с высоким разрешением» . Исследования нуклеиновых кислот . 37 (12): е86. дои : 10.1093/нар/gkp383 . ПМК 2709587 . ПМИД 19454604 .
- ^ Гохман Д., Лави Э., Прюфер К., Фрага М.Ф., Рианчо Дж.А., Келсо Дж. и др. (май 2014 г.). «Реконструкция карт метилирования ДНК неандертальцев и денисовцев» . Наука . 344 (6183): 523–527. Бибкод : 2014Sci...344..523G . дои : 10.1126/science.1250368 . ПМИД 24786081 . S2CID 28665590 .
- ^ Гохман Д., Мишол Н., де Мануэль М., де Хуан Д., Шукрун Дж., Мешорер Э. и др. (сентябрь 2019 г.). «Реконструкция денисовской анатомии с использованием карт метилирования ДНК» . Клетка . 179 (1): 180–192.e10. дои : 10.1016/j.cell.2019.08.035 . ПМИД 31539495 . S2CID 202676502 .
- ^ Jump up to: Перейти обратно: а б Форат С., Хюттель Б., Рейнхардт Р., Фиммерс Р., Хайдл Г., Деншлаг Д., Олек К. (01.02.2016). «Маркеры метилирования для идентификации жидкостей организма и тканей по судебно-медицинским доказательствам» . ПЛОС ОДИН . 11 (2): e0147973. Бибкод : 2016PLoSO..1147973F . дои : 10.1371/journal.pone.0147973 . ПМЦ 4734623 . ПМИД 26829227 .
- ^ «Анализ метилирования инфиния | Опрос отдельных сайтов CpG» . www.illumina.com . Проверено 10 января 2020 г.
- ^ «Набор Infinium MmethylationEPIC | Массив для определения профилей метилирования для EWAS» . www.illumina.com . Проверено 10 января 2020 г.
- ^ Ракян В.К., Даун Т.А., Торн Н.П., Фличек П., Кулеша Э., Граф С. и др. (сентябрь 2008 г.). «Интегрированный ресурс для полногеномной идентификации и анализа тканеспецифичных дифференциально метилированных областей (tDMR) человека» . Геномные исследования . 18 (9): 1518–1529. дои : 10.1101/гр.077479.108 . ПМК 2527707 . ПМИД 18577705 .
- ^ Ли ХИ, Пак М.Дж., Чхве А., Ан Дж.Х., Ян В.И., Шин К.Дж. (январь 2012 г.). «Потенциальное судебно-медицинское применение профилирования метилирования ДНК для идентификации жидкостей организма». Международный журнал юридической медицины . 126 (1): 55–62. дои : 10.1007/s00414-011-0569-2 . ПМИД 21626087 . S2CID 22243051 .
- ^ Иризарри Р.А., Лэдд-Акоста С., Вэнь Б., Ву З., Монтано С., Оньянго П. и др. (февраль 2009 г.). «Метилом рака толстой кишки человека демонстрирует сходное гипо- и гиперметилирование на берегах консервативных тканеспецифичных CpG-островков» . Природная генетика . 41 (2): 178–186. дои : 10.1038/ng.298 . ПМЦ 2729128 . ПМИД 19151715 .
- ^ Рейк В., Дин В., Уолтер Дж. (август 2001 г.). «Эпигенетическое перепрограммирование в развитии млекопитающих». Наука . 293 (5532): 1089–1093. дои : 10.1126/science.1063443 . ПМИД 11498579 . S2CID 17089710 .
- ^ Мейснер А., Миккельсен Т.С., Гу Х., Верниг М., Ханна Дж., Сиваченко А. и др. (август 2008 г.). «Карты метилирования ДНК плюрипотентных и дифференцированных клеток в масштабе генома» . Природа . 454 (7205): 766–770. Бибкод : 2008Natur.454..766M . дои : 10.1038/nature07107 . ПМЦ 2896277 . ПМИД 18600261 .
- ^ Дой А., Пак И.Х., Вэнь Б., Мураками П., Арье М.Дж., Иризарри Р. и др. (декабрь 2009 г.). «Дифференциальное метилирование ткане- и ракоспецифичных берегов CpG-островков отличает индуцированные человеком плюрипотентные стволовые клетки, эмбриональные стволовые клетки и фибробласты» . Природная генетика . 41 (12): 1350–1353. дои : 10.1038/ng.471 . ПМК 2958040 . ПМИД 19881528 .
- ^ Бьорнссон Х.Т., Сигурдссон М.И., Фаллин М.Д., Иризарри Р.А., Аспелунд Т., Куи Х. и др. (июнь 2008 г.). «Внутрииндивидуальные изменения метилирования ДНК с течением времени при семейной кластеризации» . ДЖАМА . 299 (24): 2877–2883. дои : 10.1001/jama.299.24.2877 . ПМК 2581898 . ПМИД 18577732 .
- ^ Бок С., Уолтер Дж., Полсен М., Ленгауэр Т. (июнь 2008 г.). «Межиндивидуальные вариации метилирования ДНК и их значение для крупномасштабного картирования эпигенома» . Исследования нуклеиновых кислот . 36 (10): е55. дои : 10.1093/нар/gkn122 . ПМЦ 2425484 . ПМИД 18413340 .
- ^ «QDMR: количественный метод идентификации дифференциально метилированных областей по энтропии» . bioinfo.hrbmu.edu.cn . Архивировано из оригинала 23 октября 2015 г. Проверено 9 марта 2013 г.
- ^ Чжан Ю, Лю Х, Льв Дж, Сяо Икс, Чжу Дж, Лю Икс и др. (май 2011 г.). «QDMR: количественный метод идентификации дифференциально метилированных областей по энтропии» . Исследования нуклеиновых кислот . 39 (9): е58. дои : 10.1093/nar/gkr053 . ПМК 3089487 . ПМИД 21306990 .
- ^ Гилихер П., Хартнетт Л., Иган Л.Дж., Голден А., Раджа Али Р.А., Сои С. (август 2013 г.). «Анализ набора генов сильно искажается, когда применяется к данным о полногеномном метилировании» . Биоинформатика . 29 (15): 1851–1857. doi : 10.1093/биоинформатика/btt311 . ПМИД 23732277 .
- ^ Лю Х, Лю X, Чжан С, Льв Дж, Ли С, Шан С и др. (январь 2016 г.). «Систематическая идентификация и аннотация меток метилирования человека на основе метиломов бисульфитного секвенирования выявляет четкую роль гипометилирования, специфичного для типа клеток, в регуляции генов идентичности клеток» . Исследования нуклеиновых кислот . 44 (1): 75–94. дои : 10.1093/nar/gkv1332 . ПМЦ 4705665 . ПМИД 26635396 .
- ^ Хунбо Л. (2016). «SMART 2: инструмент комплексного анализа данных бисульфитного секвенирования» . сайт известности.edbc.org .
- ^ Сиджен Т. (сентябрь 2015 г.). «Молекулярные подходы к судебно-медицинской идентификации типов клеток: мРНК, микроРНК, метилирование ДНК и микробные маркеры». Международная судебно-медицинская экспертиза. Генетика . 18 : 21–32. дои : 10.1016/j.fsigen.2014.11.015 . ПМИД 25488609 .
- ^ Jump up to: Перейти обратно: а б с Кадер Ф., Гай М. (апрель 2015 г.). «Метилирование ДНК и применение в судебной медицине». Международная судебно-медицинская экспертиза . 249 : 255–265. doi : 10.1016/j.forsciint.2015.01.037 . ПМИД 25732744 .
- ^ Сильва Д.С., Антунес Дж., Баламуруган К., Дункан Дж., Алхо К.С., МакКорд Б. (июль 2016 г.). «Исследование по проверке эпигенетических маркеров метилирования ДНК для обнаружения образцов крови, спермы и слюны». Международная судебно-медицинская экспертиза. Генетика . 23 : 55–63. дои : 10.1016/j.fsigen.2016.01.017 . hdl : 10923/23633 . ПМИД 27010659 . S2CID 7438775 .
- ^ Дор Ю, Кедр Ч (сентябрь 2018 г.). «Принципы метилирования ДНК и их значение для биологии и медицины». Ланцет . 392 (10149): 777–786. дои : 10.1016/S0140-6736(18)31268-6 . ПМИД 30100054 . S2CID 51965986 .
- ^ Лю MC, Окснард GR, Кляйн EA, Суонтон C, Зайден MV (июнь 2020 г.). «Чувствительное и специфическое обнаружение и локализация множественного рака с использованием сигнатур метилирования в бесклеточной ДНК» . Анналы онкологии . 31 (6): 745–759. дои : 10.1016/j.annonc.2020.02.011 . ПМК 8274402 . ПМИД 33506766 .
- ^ Мосс Дж., Магенхайм Дж., Нейман Д., Земмур Х., Лойфер Н., Корах А. и др. (ноябрь 2018 г.). «Комплексный атлас метилирования типов клеток человека раскрывает происхождение циркулирующей внеклеточной ДНК в норме и при заболеваниях» . Природные коммуникации . 9 (1): 5068. Бибкод : 2018NatCo...9.5068M . дои : 10.1038/s41467-018-07466-6 . ПМК 6265251 . ПМИД 30498206 .
- ^ Бхасин М., Чжан Х., Рейнхерц Э.Л., Рече П.А. (август 2005 г.). «Прогнозирование метилированных CpG в последовательностях ДНК с использованием машины опорных векторов» (PDF) . Письма ФЭБС . 579 (20): 4302–4308. doi : 10.1016/j.febslet.2005.07.002 . ПМИД 16051225 . S2CID 14487630 .
- ^ Бок С., Полсен М., Тирлинг С., Микеска Т., Ленгауэр Т., Уолтер Дж. (март 2006 г.). «Метилирование CpG-островков в лимфоцитах человека сильно коррелирует с последовательностью ДНК, повторами и прогнозируемой структурой ДНК» . ПЛОС Генетика . 2 (3): е26. дои : 10.1371/journal.pgen.0020026 . ПМЦ 1386721 . ПМИД 16520826 .
- ^ Чжэн Х, Цзян СВ, Ву Х (2011). «Повышение прогностической способности модели прогнозирования метилирования геномной ДНК человека». Международная конференция по биоинформатике и вычислительной биологии (BIOCOMP'11) .
- ^ Чжэн Х, Ву Х, Ли Дж, Цзян С.В. (2013). «CpGIMethPred: вычислительная модель для прогнозирования статуса метилирования CpG-островков в геноме человека» . BMC Медицинская Геномика . 6 (Приложение 1): S13. дои : 10.1186/1755-8794-6-S1-S13 . ПМЦ 3552668 . ПМИД 23369266 .
Дальнейшее чтение [ править ]
- Закон JA, Якобсен SE (март 2010 г.). «Установление, поддержание и изменение закономерностей метилирования ДНК у растений и животных» . Обзоры природы. Генетика . 11 (3): 204–220. дои : 10.1038/nrg2719 . ПМК 3034103 . ПМИД 20142834 .
- Штраусман Р., Нейман Д., Робертс Д., Стейнфельд И., Блюм Б., Бенвенисти Н. и др. (май 2009 г.). «Программирование развития профилей метилирования CpG-островков в геноме человека». Структурная и молекулярная биология природы . 16 (5): 564–571. дои : 10.1038/nsmb.1594 . ПМИД 19377480 . S2CID 8804930 .
- Патра СК (апрель 2008 г.). «Ras-регуляция метилирования ДНК и рак». Экспериментальные исследования клеток . 314 (6): 1193–1201. doi : 10.1016/j.yexcr.2008.01.012 . ПМИД 18282569 .
- Патра С.К., Патра А., Рицци Ф., Гош Т.К., Беттуцци С. (июнь 2008 г.). «Деметилирование (цитозин-5-С-метил) ДНК и регуляция транскрипции в эпигенетических путях развития рака». Обзоры рака и метастазов . 27 (2): 315–334. дои : 10.1007/s10555-008-9118-y . hdl : 11381/1797001 . ПМИД 18246412 . S2CID 22435914 .
- Нагпал Г., Шарма М., Кумар С., Чаудхари К., Гупта С., Гаутам А., Рагхава Г.П. (февраль 2014 г.). «PCMdb: база данных метилирования рака поджелудочной железы» . Научные отчеты . 4 : 4197. Бибкод : 2014NatSR...4E4197N . дои : 10.1038/srep04197 . ПМЦ 3935225 . ПМИД 24569397 .
Внешние ссылки [ править ]
- ДНК + метилирование в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Обозреватель потоков ENCODE. Характеристика некодирующих РНК. Природа (журнал)
- База данных PCMdb по метилированию рака поджелудочной железы.
- Инструмент SMART для анализа специфического метилирования и составления отчетов
- Метилирование человека Марк Атлас
- БолезньМет. Архивировано 27 января 2020 г. в Wayback Machine. базе данных метилирования болезней человека
- Атлас EWAS База знаний об исследованиях эпигеномных ассоциаций.