МЕСР2
| МЕСР2 | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | MECP2 , AUTSX3, MRX16, MRX79, MRXS13, MRXSL, PPMX, RS, RTS, RTT, белок 2, связывающий метил-CpG | ||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 300005 ; МГИ : 99918 ; Гомологен : 3657 ; Генные карты : MECP2 ; OMA : MECP2 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||







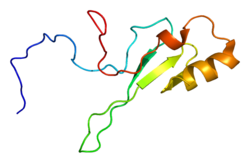
MECP2 ( метил-CpG-связывающий белок 2 ) представляет собой ген [ 5 ] который кодирует белок MECP2. [ 6 ] MECP2, по-видимому, необходим для нормального функционирования нервных клеток . Этот белок, по-видимому, особенно важен для зрелых нервных клеток, где он присутствует в больших количествах. Белок MECP2, вероятно, участвует в выключении («подавлении» или « замалчивании ») некоторых других генов. Это не позволяет генам производить белки, когда они не нужны. Недавняя работа показала, что MECP2 также может активировать другие гены. [ 7 ] Ген MECP2 расположен на длинном (q) плече Х-хромосомы в полосе 28 («Xq28»), от пары оснований 152 808 110 до пары оснований 152 878 611.
MECP2 является важным считывателем метилирования ДНК. Его метил-CpG-связывающий (MBD) домен распознает и связывает 5-mC области . MECP2 связан с Х-хромосомой и подвержен X-инактивации . Мутации гена MECP2 являются причиной большинства случаев синдрома Ретта , прогрессирующего неврологического нарушения развития и одной из наиболее частых причин когнитивных нарушений у женщин. [ 8 ] Обнаружено по меньшей мере 53 мутации этого гена, вызывающие заболевания. [ 9 ]
Функция
[ редактировать ]Белок MECP2 обнаруживается во всех клетках организма, включая мозг , и действует как репрессор и активатор транскрипции, в зависимости от контекста. Однако идея о том, что MECP2 действует как активатор, относительно нова и остается спорной. [ 10 ] В головном мозге он обнаруживается в высоких концентрациях в нейронах и связан с созреванием центральной нервной системы (ЦНС) и формированием синаптических контактов . [ 11 ]
Механизм действия
[ редактировать ]Белок MeCP2 связывается с ДНК метилированными формами . Затем белок MeCP2 взаимодействует с другими белками, образуя комплекс, который выключает этот ген. MeCP2 предпочитает связываться с участками генома с химическим изменением цитозина ( C), когда он встречается в определенной последовательности ДНК, « CpG ». Это форма метилирования ДНК . Многие гены имеют CpG-островки , которые часто встречаются вблизи начала гена. MECP2 в большинстве случаев не связывается с этими островками, поскольку они не метилированы. Экспрессия . некоторых генов может регулироваться посредством метилирования их CpG-островков, и MECP2 может играть роль в их подмножестве Исследователи еще не определили, на какие гены воздействует белок MeCP2, но такие гены, вероятно, важны для нормальной функции центральной нервной системы. Однако первое крупномасштабное картирование сайтов связывания MECP2 в нейронах показало, что только 6% сайтов связывания находятся в островках CpG, и что 63% связанных с MECP2 промоторов активно экспрессируются и только 6% сильно метилированы, что указывает на то, что Основная функция MECP2 – это нечто иное, чем подавление метилированных промоторов. [ 12 ]
После связывания MeCP2 конденсирует структуру хроматина , образует комплекс с деацетилазами гистонов (HDAC) или блокирует факторы транскрипции напрямую . Более поздние исследования показали, что MeCP2 может также функционировать как активатор транскрипции посредством рекрутирования транскрипционного фактора CREB1 . Это было неожиданное открытие, которое предполагает, что MeCP2 является ключевым регулятором транскрипции, потенциально играющим двойную роль в экспрессии генов. Фактически, большинство генов, регулируемых MeCP2, активируются, а не репрессируются. [ 13 ] Однако остается спорным, регулирует ли MeCP2 эти гены напрямую или эти изменения носят вторичный характер. [ 10 ] Дальнейшие исследования показали, что в некоторых случаях MeCP2 может напрямую связываться с неметилированной ДНК. [ 14 ] MeCP2 участвует в регуляции импринтированных генов и локусов, которые включают UBE3A и DLX5 . [ 15 ]
Снижение экспрессии MECP2 в нервных стволовых клетках Mecp2+/- вызывает усиление старения , нарушение пролиферативной способности и накопление невосстановленных повреждений ДНК . [ 16 ] После обработки клеток Mecp2+/- любым из трех различных агентов, повреждающих ДНК, клетки накапливали больше поврежденной ДНК и были более склонны к гибели клеток, чем контрольные клетки. [ 16 ] Был сделан вывод, что снижение экспрессии MECP2 приводит к снижению способности к восстановлению ДНК и, вероятно, способствует неврологическому ухудшению. [ 16 ]
Структура
[ редактировать ]MECP2 является частью семейства белков метил-CpG-связывающего домена (MBD), но обладает своими уникальными отличиями, которые помогают выделить его из этой группы. Он имеет две функциональные области:
- метилцитозин - связывающий домен (MBD), состоящий из 85 аминокислот ; и
- домен репрессии транскрипции (TRD), состоящий из 104 аминокислот
Домен MBD образует клин и прикрепляется к метилированным сайтам CpG на цепях ДНК. Затем область TRD реагирует с SIN3A, рекрутируя деацетилазы гистонов (HDAC). [ 17 ] На карбоксильном конце обнаружены также необычные повторяющиеся последовательности. Эта область тесно связана с семейством вилкообразных на уровне аминокислот. [ 18 ]
Роль в болезни
[ редактировать ]Роль MECP2 при заболевании в первую очередь связана либо с потерей функции (недостаточной экспрессией) гена MECP2, как при синдроме Ретта , либо с усилением функции (чрезмерной экспрессией), как при синдроме дупликации MECP2 . Многие мутации связаны с потерей экспрессии гена MECP2 и были выявлены у пациентов с синдромом Ретта. Эти мутации включают изменения в отдельных парах оснований ДНК ( SNP ), вставки или делеции ДНК в гене MECP2 , а также изменения, которые влияют на то, как информация гена преобразуется в белок ( сплайсинг РНК ). Мутации в гене изменяют структуру белка MeCP2 или приводят к уменьшению количества белка. В результате белок не может связываться с ДНК или включать или выключать другие гены. Гены, которые обычно репрессируются MeCP2, остаются активными, когда их продукты не нужны. Другие гены, которые обычно активируются MeCP2, остаются неактивными, что приводит к отсутствию генного продукта. Этот дефект, вероятно, нарушает нормальное функционирование нервных клеток, что приводит к появлению признаков и симптомов синдрома Ретта.
Синдром Ретта в основном встречается у девочек, его распространенность составляет около 1 на каждые 10 000; плоды мужского пола с нормальным кариотипом, страдающие этим заболеванием, редко доживают до срока, а если и доживают, то обычно умирают вскоре после рождения. Пациенты рождаются с очень трудно обнаруживаемыми признаками расстройства, но примерно через шесть месяцев-полтора года речевые и двигательные функции начинают снижаться. За этим следуют судороги , задержка роста, когнитивные и двигательные нарушения. [ 19 ] Локус MECP2 сцеплен с Х-хромосомой , и аллели, вызывающие заболевание, являются доминантными . Из-за того, что он распространен среди женщин, он связан с мужской смертностью или с преимущественной передачей инфекции через отцовскую Х-хромосому; тем не менее, в редких случаях синдромом Ретта могут страдать и некоторые мужчины. [ 20 ] Мужчины с дупликацией гена MECP-2 в локусе Xq28 также подвержены риску рецидивирующих инфекций и менингита в младенчестве.
Мутации в гене MECP2 также были выявлены у людей с рядом других заболеваний, поражающих центральную нервную систему. Например, мутации MECP2 связаны с некоторыми случаями Х-сцепленной умственной отсталости от умеренной до тяжелой степени. Мутации в этом гене также были обнаружены у мужчин с тяжелой мозговой дисфункцией ( неонатальной энцефалопатией ), которые доживают только до раннего детства. Кроме того, у нескольких людей с признаками синдрома Ретта и синдрома Ангельмана (состояние, характеризующееся умственной отсталостью, проблемами с движением, неуместным смехом и возбудимостью) наблюдаются мутации в гене MECP2. сообщалось о мутациях MECP2 или изменениях активности гена . Наконец, в некоторых случаях аутизма (нарушения развития, влияющего на общение и социальное взаимодействие) [ 21 ]
В более поздних исследованиях сообщалось о генетическом полиморфизме генов MeCP2 у пациентов с системной красной волчанкой (СКВ). [ 22 ] СКВ — системное аутоиммунное заболевание, которое может поражать несколько органов. Полиморфизм MeCP2 до сих пор был зарегистрирован у пациентов с волчанкой европейского и азиатского происхождения.
Генетическая потеря MECP2 была идентифицирована как изменение свойств клеток голубого пятна , единственного источника норадренергической иннервации коры головного мозга и гиппокампа . [ 23 ]
Исследователи пришли к выводу, что «поскольку эти нейроны являются основным источником норадреналина в стволе мозга и переднем мозге и участвуют в регуляции различных функций, нарушаемых при синдроме Ретта, таких как дыхание и познание, мы предполагаем, что голубое пятно является критическим местом в мозге». потеря MECP2 приводит к дисфункции ЦНС». [ 23 ]
Интерактивная карта маршрутов
[ редактировать ]Нажмите на гены, белки и метаболиты ниже, чтобы просмотреть соответствующие статьи. [ § 1 ]

Взаимодействия
[ редактировать ]Было показано, что MECP2 взаимодействует с белком SKI. [ 24 ] и ко-репрессор ядерных рецепторов 1 . [ 24 ] Считается, что в нейрональных клетках мРНК MECP2 взаимодействует с миР-132 , которая подавляет экспрессию белка. Это является частью гомеостатического механизма, который может регулировать уровень MECP2 в мозге. [ 25 ]
MeCP2 и гормоны
[ редактировать ]MeCP2 в развивающемся мозге крысы регулирует важное социальное развитие половым диморфизмом. Уровни MeCP2 различаются у самцов и самок в развивающемся мозге крысы через 24 часа после рождения в миндалевидном теле и гипоталамусе , но эта разница больше не наблюдается через 10 дней после рождения. В частности, мужчины экспрессируют меньше MeCP2, чем женщины. [ 26 ] и это согласуется с периодом времени, чувствительным к стероидам, в мозге новорожденных крыс. Снижение MeCP2 с помощью малой интерферирующей РНК (миРНК) в течение первых нескольких дней жизни снижает уровень социального игрового поведения юношей у самцов до типичных для самок уровней, но не влияет на игровое поведение юношей у самок. [ 27 ]
MeCP2 важен для организации гормонально-зависимого поведения и половых различий в развивающейся миндалине крысы. MeCP2, по-видимому, регулирует выработку аргинин-вазопрессина (AVP) и андрогенных рецепторов (AR) у самцов крыс, но не у самок. Известно, что вазопрессин регулирует многие виды социального поведения, включая формирование парных связей. [ 28 ] и общественное признание. [ 29 ] Хотя самцы крыс обычно имеют более высокий уровень вазопрессина в миндалевидном теле, [ 30 ] Снижение MeCP2 в течение первых 3 дней жизни вызывает стойкое снижение уровня вазопрессина в этой области мозга до типичного для женщин уровня, которое сохраняется на протяжении всей взрослой жизни. У самцов крыс со сниженным уровнем MeCP2 также наблюдается значительное снижение АР через две недели после инфузии, но этот эффект исчезает к взрослому возрасту. [ 31 ]
Стресс в раннем возрасте
[ редактировать ]MeCP2 отслеживает реакцию на стресс в раннем возрасте. Стресс в раннем возрасте коррелирует с гиперфосфорилированием белка MeCP2 в паравентрикулярном ядре гипоталамуса. [ 32 ] Таким образом, это вызывает снижение присутствия MeCP2 в промоторной области гена AVP и, следовательно, повышение уровней AVP. Вазопрессин — это основной гормон, участвующий в работе гипоталмо-гипофизарно-надпочечниковой системы, связующего звена в мозге, которое регулирует обработку и реакцию на стресс. Таким образом, снижение функционирования белка MeCP2 усиливает реакцию нейронов на стресс.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000169057 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000031393 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Амир Р.Э., Ван ден Вейвер И.Б., Ван М., Тран CQ, Франке У., Зогби Х.Ю. (октябрь 1999 г.). «Синдром Ретта вызван мутациями в X-связанном MECP2, кодирующем метил-CpG-связывающий белок 2». Нат. Жене . 23 (2): 185–8. дои : 10.1038/13810 . ПМИД 10508514 . S2CID 3350350 .
- ^ Льюис Дж.Д., Михан Р.Р., Хензель В.Дж., Маурер-Фоги И., Джеппесен П., Кляйн Ф., Берд А. (июнь 1992 г.). «Очистка, последовательность и клеточная локализация нового хромосомного белка, который связывается с метилированной ДНК». Клетка . 69 (6): 905–14. дои : 10.1016/0092-8674(92)90610-О . ПМИД 1606614 . S2CID 6825994 .
- ^ Чахрур М. и др. (2008). «MECP2, ключевой фактор неврологических заболеваний, активирует и подавляет транскрипцию» . Наука . 320 (5880): 1224–9. Бибкод : 2008Sci...320.1224C . дои : 10.1126/science.1153252 . ПМЦ 2443785 . ПМИД 18511691 .
- ^ «Ген Энтреза: белок 2, связывающий метил CpG MECP2 (синдром Ретта)» .
- ^ Шимчикова Д., Хенеберг П. (декабрь 2019 г.). «Уточнение прогнозов эволюционной медицины на основе клинических данных о проявлениях менделевских болезней» . Научные отчеты . 9 (1): 18577. Бибкод : 2019NatSR...918577S . дои : 10.1038/s41598-019-54976-4 . ПМК 6901466 . ПМИД 31819097 .
- ^ Перейти обратно: а б Коэн С., Чжоу З., Гринберг М.Е. (май 2008 г.). «Медицина. Активация репрессора» . Наука . 320 (5880): 1172–3. дои : 10.1126/science.1159146 . ПМЦ 2857976 . ПМИД 18511680 .
- ^ Луикенхейс С., Джакометти Э., Берд К.Ф., Джениш Р. (апрель 2004 г.). «Экспрессия MeCP2 в постмитотических нейронах спасает синдром Ретта у мышей» . Учеб. Натл. акад. наук. США . 101 (16): 6033–8. Бибкод : 2004PNAS..101.6033L . дои : 10.1073/pnas.0401626101 . ПМК 395918 . ПМИД 15069197 .
- ^ Ясуи Д.Х., Педдада С., Биеда М.К., Валлеро Р.О., Хогарт А., Нагараджан Р.П., Тэтчер К.Н., Фарнхэм П.Дж., Ласалле Дж.М. (декабрь 2007 г.). «Комплексный эпигеномный анализ нейронального MeCP2 выявляет роль дальнего взаимодействия с активными генами» . Учеб. Натл. акад. наук. США . 104 (49): 19416–21. Бибкод : 2007PNAS..10419416Y . дои : 10.1073/pnas.0707442104 . ПМК 2148304 . ПМИД 18042715 .
- ^ Чахрур М., Юнг С.Ю., Шоу С., Чжоу Х., Вонг С.Т., Цинь Дж., Зогби Х.И. (май 2008 г.). «MeCP2, ключевой фактор неврологических заболеваний, активирует и подавляет транскрипцию» . Наука . 320 (5880): 1224–9. Бибкод : 2008Sci...320.1224C . дои : 10.1126/science.1153252 . ПМЦ 2443785 . ПМИД 18511691 .
- ^ Джорджель П.Т., Горовиц-Шерер Р.А., Адкинс Н., Вудкок К.Л., Уэйд П.А., Хансен Дж.К. (август 2003 г.). «Уплотнение хроматина с помощью MeCP2 человека. Сборка новых вторичных структур хроматина в отсутствие метилирования ДНК» . Ж. Биол. Хим . 278 (34): 32181–8. дои : 10.1074/jbc.M305308200 . ПМИД 12788925 .
- ^ ЛаСалль Ж.М. (2007). «Одиссея MeCP2 и родительского импринтинга» . Эпигенетика . 2 (1): 5–10. дои : 10.4161/epi.2.1.3697 . ПМК 1866173 . ПМИД 17486180 .
- ^ Перейти обратно: а б с Алессио Н., Риччиелло Ф., Скилларо Т., Капассо С., Дель Гаудио С., Ди Бернардо Г., Чиполларо М., Мелоне МАБ, Пелузо Г., Галдеризи У. (март 2018 г.). «Нейральные стволовые клетки из мышиной модели синдрома Ретта склонны к старению, демонстрируют пониженную способность справляться с генотоксическим стрессом и нарушаются в процессе дифференцировки» . Эксп. Мол. Мед . 50 (3): 1. дои : 10.1038/s12276-017-0005-x . ПМК 6118406 . ПМИД 29563495 .
- ^ Уэйкфилд Р.И., Смит Б.О., Нэн Х, Фри А., Сотериу А., Ухрин Д., Берд А.П., Барлоу П.Н. (сентябрь 1999 г.). «Структура раствора домена MeCP2, который связывается с метилированной ДНК». Дж. Мол. Биол . 291 (5): 1055–65. дои : 10.1006/jmbi.1999.3023 . ПМИД 10518942 .
- ^ Пол А. Уэйд (декабрь 2001 г.). «Метил-CpG-связывающие белки и репрессия транскрипции» (PDF) . Биоэссе . 23 (12): 1131–1137. дои : 10.1002/bies.10008 . ПМИД 11746232 . S2CID 37525856 . Архивировано из оригинала (PDF) 14 августа 2007 г.
- ^ Кабальеро И.М., Хендрих Б. (апрель 2005 г.). «MeCP2 в нейронах: приближаемся к причинам синдрома Ретта» . Хм. Мол. Жене . 14 Спецификация № 1: R19–26. дои : 10.1093/hmg/ddi102 . ПМИД 15809268 .
- ^ Самако Р.К., Нагараджан Р.П., Брауншвейг Д., ЛаСалль Ж.М. (март 2004 г.). «Множественные пути регулируют экспрессию MeCP2 при нормальном развитии мозга и обнаруживают дефекты при расстройствах аутистического спектра» . Хм. Мол. Жене . 13 (6): 629–39. дои : 10.1093/hmg/ddh063 . ПМИД 14734626 .
- ^ Хант, Кэти (12 января 2016 г.). «Китайские учёные создали обезьян с геном аутизма» . Новости CNN . Проверено 27 января 2016 г.
- ^ Савалья А.Х., Уэбб Р., Хан С., Келли Дж.А., Кауфман К.М., Кимберли Р.П., Аларкон-Рикельме М.Э., Джеймс Дж.А., Вайс Т.Дж., Гилкесон Г.С., Чой CB, Скофилд Р.Х., Бэй СК, Нат СК, Харли Дж.Б. (2008). Джин Д.И. (ред.). «Распространенные варианты MECP2 повышают риск системной красной волчанки» . ПЛОС ОДИН . 3 (3): e1727. Бибкод : 2008PLoSO...3.1727S . дои : 10.1371/journal.pone.0001727 . ПМК 2253825 . ПМИД 18320046 .

- ^ Перейти обратно: а б Танея П., Ожье М., Брукс-Харрис Г., Шмид Д.А., Кац Д.М., Нельсон С.Б. (2009). «Патофизиология нейронов Locus Ceruleus в мышиной модели синдрома Ретта» . Журнал неврологии . 29 (39): 12187–12195. doi : 10.1523/JNEUROSCI.3156-09.2009 . ПМЦ 2846656 . ПМИД 19793977 .
- ^ Перейти обратно: а б Кокура К., Каул С.К., Вадхва Р., Номура Т., Хан М.М., Синагава Т., Ясукава Т., Кольменарес К., Исии С. (сентябрь 2001 г.). «Семейство белков Ski необходимо для репрессии транскрипции, опосредованной MeCP2» . Ж. Биол. Хим . 276 (36): 34115–21. дои : 10.1074/jbc.M105747200 . ПМИД 11441023 .
- ^ Кляйн М.Е., Лиой Д.Т., Ма Л., Импи С., Мандель Г., Гудман Р.Х. (декабрь 2007 г.). «Гомеостатическая регуляция экспрессии MeCP2 с помощью CREB-индуцированной микроРНК». Нат. Нейроски . 10 (12): 1513–4. дои : 10.1038/nn2010 . ПМИД 17994015 . S2CID 29308441 .
- ^ Куриан-младший, Форбс-Лорман РМ, Оже АП (сентябрь 2007 г.). «Половые различия в экспрессии mecp2 в критический период развития мозга крыс» . Эпигенетика . 2 (3): 173–8. дои : 10.4161/epi.2.3.4841 . ПМИД 17965589 .
- ^ Куриан-младший, Быховский М.Э., Форбс-Лорман Р.М., Оже С.Дж., Оже А.П. (июль 2008 г.). «Mecp2 организует социальное поведение несовершеннолетних в зависимости от пола» . Дж. Нейроски . 28 (28): 7137–42. doi : 10.1523/JNEUROSCI.1345-08.2008 . ПМК 2569867 . ПМИД 18614683 .
- ^ Уинслоу Дж.Т., Гастингс Н., Картер К.С., Харбо Ч.Р., Инсел Т.Р. (октябрь 1993 г.). «Роль центрального вазопрессина в образовании пар у моногамных степных полевок». Природа . 365 (6446): 545–8. Бибкод : 1993Natur.365..545W . дои : 10.1038/365545a0 . ПМИД 8413608 . S2CID 4333114 .
- ^ Бельский И.Ф., Ху С.Б., Сегда К.Л., Вестфаль Х., Янг Л.Дж. (март 2004 г.). «Глубокое нарушение социального распознавания и снижение тревожного поведения у мышей с нокаутом рецептора вазопрессина V1a» . Нейропсихофармакология . 29 (3): 483–93. дои : 10.1038/sj.npp.1300360 . ПМИД 14647484 .
- ^ Де Врис Г.Дж., Панзика Г.К. (2006). «Половая дифференциация центральных систем вазопрессина и вазотоцина у позвоночных: разные механизмы, схожие конечные точки» . Нейронаука . 138 (3): 947–55. doi : 10.1016/j.neuroscience.2005.07.050 . ПМЦ 1457099 . ПМИД 16310321 .
- ^ Форбс-Лорман Р.М., Раутио Дж.Дж., Куриан Дж.Р., Оже А.П., Оже С.Дж. (март 2012 г.). «Неонатальный MeCP2 важен для организации половых различий в экспрессии вазопрессина» . Эпигенетика . 7 (3): 230–8. дои : 10.4161/epi.7.3.19265 . ПМЦ 3335947 . ПМИД 22430799 .
- ^ Мургатройд С., Патчев А.В., Ву Ю., Микале В., Бокмюль Ю., Фишер Д., Холсбур Ф., Вотьяк К.Т., Алмейда О.Ф., Шпенглер Д. (декабрь 2009 г.). «Динамическое метилирование ДНК программирует стойкие неблагоприятные последствия стресса в раннем возрасте». Нат. Нейроски . 12 (12): 1559–66. дои : 10.1038/nn.2436 . ПМИД 19898468 . S2CID 3328884 .
Дальнейшее чтение
[ редактировать ]- Чахрур М., Зогби Х.И. (2007). «История синдрома Ретта: от клиники к нейробиологии» . Нейрон . 56 (3): 422–37. дои : 10.1016/j.neuron.2007.10.001 . ПМИД 17988628 . S2CID 16266882 .
- Карни Р.М., Вулперт С.М., Раван С.А., Шахбазян М., Эшли-Кох А., Куккаро М.Л., Вэнс Дж.М., Перикак-Вэнс М.А. (2003). «Идентификация мутаций MeCP2 у ряда женщин с аутистическим расстройством». Педиатр Нейрол . 28 (3): 205–11. дои : 10.1016/S0887-8994(02)00624-0 . ПМИД 12770674 .
- Керр А.М., Рэвин Д. (2003). «Обзорная статья: открытие новых горизонтов с синдромом Ретта» . J Отключение интеллекта Res . 47 (Часть 8): 580–7. дои : 10.1046/j.1365-2788.2003.00506.x . ПМИД 14641805 .
- Неул Дж.Л., Зогби Х.И. (2004). «Синдром Ретта: прототипическое расстройство нервного развития». Нейробиолог . 10 (2): 118–28. дои : 10.1177/1073858403260995 . ПМИД 15070486 . S2CID 9617631 .
- Шанен С., Хаувинк Э.Дж., Доррани Н., Лейн Дж., Эверетт Р., Фэн А., Кантор Р.М., Перси А. (2004). «Фенотипические проявления мутаций MECP2 при классическом и атипичном синдроме Ретта». Am J Med Genet A. 126 (2): 129–40. doi : 10.1002/ajmg.a.20571 . ПМИД 15057977 . S2CID 32897044 .
- Ван ден Вейвер И.Б., Зогби Х.Ю. (2001). «Мутации в гене, кодирующем метил-CpG-связывающий белок 2, вызывают синдром Ретта». Мозговой разработчик . 23 (Приложение 1): S147–51. дои : 10.1016/S0387-7604(01)00376-X . ПМИД 11738862 . S2CID 26138178 .
- Уэбб Т., Латиф Ф. (2001). «Синдром Ретта и ген MECP2» . Джей Мед Жене . 38 (4): 217–23. дои : 10.1136/jmg.38.4.217 . ПМЦ 1734858 . ПМИД 11283201 .
- Шахбазян, доктор медицинских наук, Зогби Х.Ю. (2003). «Синдром Ретта и MeCP2: связь эпигенетики и функции нейронов» . Являюсь. Дж. Хум. Жене . 71 (6): 1259–72. дои : 10.1086/345360 . ПМЦ 378559 . ПМИД 12442230 .
- Муг У., Смитс Э.Э., ван Роозендаль К.Е. и др. (2003). «Нарушения развития нервной системы у мужчин, связанные с геном, вызывающим синдром Ретта у женщин (MECP2)». Евро. Ж. Педиатр. Нейрол . 7 (1): 5–12. дои : 10.1016/S1090-3798(02)00134-4 . ПМИД 12615169 .
- Мильтенбергер-Милтени Г., Лакконе Ф. (2004). «Мутации и полиморфизмы в метил-CpG-связывающем белке человека MECP2» . Хм. Мутат . 22 (2): 107–15. дои : 10.1002/humu.10243 . ПМИД 12872250 . S2CID 42516576 .
- Уивинг Л.С., Эллауэй С.Дж., Геч Дж., Христодулу Дж. (2006). «Синдром Ретта: клинический обзор и генетическое обновление» . Дж. Мед. Жене . 42 (1): 1–7. дои : 10.1136/jmg.2004.027730 . ПМК 1735910 . ПМИД 15635068 .
- Бапат С., Галанд С. (2005). «Ассоциация по вине: идентификация DLX5 как мишени для MeCP2 обеспечивает молекулярную связь между геномным импринтингом и синдромом Ретта». Биоэссе . 27 (7): 676–80. дои : 10.1002/bies.20266 . ПМИД 15954098 .
- Златанова Ю (2005). «MeCP2: соединение хроматина и не только». Биохим. Клеточная Биол . 83 (3): 251–62. дои : 10.1139/o05-048 . ПМИД 15959553 .
- Кауфманн В.Е., Джонстон М.В., Blue ME (2006). «Экспрессия и функция MeCP2 во время развития мозга: значение для патогенеза и клинической эволюции синдрома Ретта». Мозговой разработчик . 27 (Приложение 1): S77–S87. дои : 10.1016/j.braindev.2004.10.008 . ПМИД 16182491 . S2CID 702975 .
- Армстронг Д.Д. (2006). «Можем ли мы связать дефицит MeCP2 со структурными и химическими аномалиями в мозге Ретта?». Мозговой разработчик . 27 (Приложение 1): S72–S76. дои : 10.1016/j.braindev.2004.10.009 . ПМИД 16182497 . S2CID 45587850 .
- Сантос М., Коэльо П.А., Масиэль П. (2006). «Ремоделирование хроматина и функция нейронов: захватывающие связи» . Гены, мозг и поведение . 5 (Приложение 2): 80–91. дои : 10.1111/j.1601-183X.2006.00227.x . hdl : 1822/67940 . ПМИД 16681803 .
- Бьенвеню Т., Челли Дж. (2006). «Молекулярная генетика синдрома Ретта: когда метилирование ДНК остается нераспознанным». Обзоры природы Генетика . 7 (6): 415–26. дои : 10.1038/nrg1878 . ПМИД 16708070 . S2CID 28215286 .
- Франке У (2007). «Механизмы заболевания: нейрогенетика дефицита MeCP2». Природная клиническая практика Неврология . 2 (4): 212–21. дои : 10.1038/ncpneuro0148 . ПМИД 16932552 . S2CID 22710951 .
Внешние ссылки
[ редактировать ]- Международный фонд синдрома Ретта
- Благотворительная организация Rett UK по поддержке и исследованиям
- Реверс вправо, Великобритания
- Правовой реестр Великобритании
- Фонд исследования синдрома Ретта
- Ансамбль Ген ссылка Белок ссылка
- GeneCard
- RettBASE: База данных вариантов IRSA MECP2
- Запись GeneReview/NIH/UW о расстройствах, связанных с MECP2
- Запись GeneReviews/NCBI/NIH/UW о синдроме дупликации MECP2
- Британский сайт для семей, пострадавших от MECP2.
- Сайт для семей, пострадавших от MECP2.
- Официальный французский сайт о дублировании MeCP2
галерея PDB |
|---|
| Базы данных органов управления : Национальные |
|---|