Переключение класса иммуноглобулина
Переключение класса иммуноглобулина , также известное как переключение изотипа , изотипическая коммутация или рекомбинация с переключением класса ( CSR ), представляет собой биологический механизм, который изменяет В-клетками выработку иммуноглобулина с одного типа на другой, например, с изотипа IgM на изотип. IgG . [ 1 ] Во время этого процесса часть константной области тяжелой цепи антитела изменяется, но вариабельная область тяжелой цепи остается прежней (термины «вариабельная» и «константа» относятся к изменениям или их отсутствию между антителами, нацеленными на разные эпитопы ). Поскольку вариабельная область не меняется, переключение классов не влияет на специфичность антигена. Вместо этого антитело сохраняет сродство к одним и тем же антигенам, но может взаимодействовать с разными эффекторными молекулами.
Механизм
[ редактировать ]Переключение классов происходит после активации зрелой B-клетки через ее мембраносвязанную молекулу антитела (или рецептор B-клетки ) для генерации различных классов антител, все с теми же вариабельными доменами, что и исходное антитело, генерируемое в незрелой B-клетке во время процесса. рекомбинации V(D)J , но обладающие отдельными константными доменами в своих тяжелых цепях . [ 2 ]
Наивные зрелые В-клетки производят как IgM , так и IgD , которые являются первыми двумя сегментами тяжелой цепи в локусе иммуноглобулина . После активации антигеном эти В-клетки пролиферируют. Если эти активированные В-клетки сталкиваются со специфическими сигнальными молекулами через свои рецепторы CD40 и цитокинов (оба модулируются Т-хелперными клетками ), они подвергаются переключению класса антител для производства антител IgG, IgA или IgE. Во время переключения класса константная область тяжелой цепи иммуноглобулина меняется, а вариабельные области — нет, и, следовательно, антигенная специфичность остается прежней. Это позволяет различным дочерним клеткам одной и той же активированной В-клетки продуцировать антитела разных изотипов или подтипов (например, IgG1, IgG2 и т. д.). [ 3 ]
У человека порядок экзонов тяжелой цепи следующий:
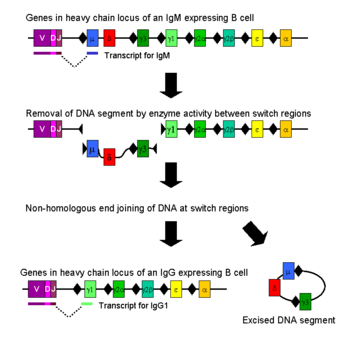
Переключение классов происходит с помощью механизма, называемого привязкой рекомбинации переключения классов (CSR). Рекомбинация переключения класса — это биологический механизм, который позволяет классу антител, продуцируемому активированной В-клеткой, изменяться во время процесса, известного как переключение изотипа или класса. тяжелой цепи антитела Во время CSR части локуса удаляются из хромосомы , а сегменты гена, окружающие удаленную часть, воссоединяются, чтобы сохранить функциональный ген антитела, который продуцирует антитело другого изотипа . Двухцепочечные разрывы генерируются в ДНК на консервативных нуклеотидных мотивах, называемых переключающими (S) областями, которые расположены выше генных сегментов, кодирующих константные области тяжелых цепей антител ; они расположены рядом со всеми генами константной области тяжелой цепи, за исключением δ-цепи. ДНК разрывается в двух выбранных S-областях под действием ряда ферментов , включая индуцируемую активацией (цитидин) деаминазу (AID), урацил- ДНК-гликозилазу и апиримидные/апуриновые (AP)-эндонуклеазы. . [ 5 ] [ 6 ] AID начинает процесс переключения классов с дезаминирования (удаления аминогруппы) цитозинов в S-областях, превращая исходные основания C в дезоксиуридин и позволяя урацилгликозилазе удалить основание. Это позволяет AP-эндонуклеазам разрезать новообразованный абазисный сайт, создавая начальные SSB, которые спонтанно образуют DSB. [ 7 ] Промежуточная ДНК между S-областями впоследствии удаляется из хромосомы, удаляя нежелательные экзоны константной области тяжелой цепи μ или δ и обеспечивая замену генного сегмента константной области γ, α или ε. Свободные концы ДНК воссоединяются с помощью процесса, называемого негомологичным соединением концов вариабельного домена (NHEJ), чтобы связать экзон с желаемым расположенным ниже экзоном константного домена тяжелой цепи антитела. [ 8 ] В отсутствие негомологичного соединения концов свободные концы ДНК могут быть воссоединены альтернативным путем, склонным к микрогомологическим соединениям. [ 9 ] За исключением генов μ и δ, только один класс антител экспрессируется В-клеткой в любой момент времени. Хотя рекомбинация переключения класса в основном представляет собой делеционный процесс, перестраивающий хромосому в «цис», она также может происходить (в 10–20% случаев, в зависимости от класса Ig) как межхромосомная транслокация, смешивающая гены тяжелой цепи иммуноглобулина из обоих аллели. [ 10 ] [ 11 ]
Цитокины, ответственные за переключение классов
[ редактировать ]Цитокины Т-клеток модулируют переключение классов у мышей (таблица 1) и человека (таблица 2). [ 12 ] [ 13 ] Эти цитокины могут оказывать подавляющее действие на продукцию IgM.
| Т-клетки | Цитокины | Классы иммуноглобулинов | |||||
|---|---|---|---|---|---|---|---|
| IgG1 | IgG2a | IgG2b | IgG3 | IgG4 | IgE | ||
| Т ч 2 | Ил-4 | ↑ | ↓ | ↓ | ↓ | ↓ | ↑ |
| Ил-5 | ↑ | ||||||
| Че 1 | ИФНγ | ↓ | ↑ | ↓ | ↑ | ↓ | ↓ |
| Трег | ТФРβ | ↑ | ↓ | ↑ | |||
| Ил-10 [ 14 ] | ↑ | ||||||
| Т-клетки | Цитокины | Классы иммуноглобулинов | |||||
|---|---|---|---|---|---|---|---|
| IgG1 | IgG2 | IgG3 | IgG4 | IgA | IgE | ||
| Т ч 2 | Ил-4 | ↑ | ↓ | ↑ | ↑ | ||
| Ил-5 | ↑ | ||||||
| Че 1 | ИФНγ | ↓ | ↑ | ↓ | |||
| Трег | ТФРβ | ↓ | ↑ | ||||
| Ил-10 [ 15 ] [ 16 ] | ↑ | ↑ | |||||
Регуляторные последовательности генов, ответственные за переключение классов
[ редактировать ]В дополнение к высокоповторяющейся структуре целевых S-областей, процесс переключения классов требует, чтобы S-области были сначала транскрибированы и сплайсированы из транскриптов тяжелой цепи иммуноглобулина (где они лежат внутри интронов). Ремоделирование хроматина, доступность транскрипции и AID, а также синапсис сломанных S-областей находятся под контролем большого суперэнхансера, расположенного ниже более дистального гена Calpha, 3'-регуляторной области (3'RR). [ 17 ] В некоторых случаях суперэнхансер 3'RR сам может быть нацелен на AID и подвергаться разрывам ДНК и соединению с Sμ, что затем удаляет локус тяжелой цепи Ig и определяет локус суицидальной рекомбинации (LSR). [ 18 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Бауэр Г (2020). «Вариабельность серологического ответа на SARS-коронавирус-2: потенциальное разрешение неоднозначности посредством определения авидности (функциональной близости)» . Журнал медицинской вирусологии . 93 (1): 311–322. дои : 10.1002/jmv.26262 . ПМЦ 7361859 . ПМИД 32633840 .
- ^ Элеонора Маркет, Ф. Нина Папавасилиу (2003) Рекомбинация V (D) J и эволюция адаптивной иммунной системы PLoS Biology 1 (1): e16.
- ^ Ставнезер Дж., Амемия К.Т. (2004). «Эволюция переключения изотипов». Семин. Иммунол . 16 (4): 257–75. дои : 10.1016/j.smim.2004.08.005 . ПМИД 15522624 .
- ^ Пархэм, Питер (2015). Иммунная система (4-е изд.). Гирляндная наука.
- ^ Дюранди А. (2003). «Цитидиндезаминаза, индуцированная активацией: двойная роль в рекомбинации с переключением классов и соматической гипермутации». Евро. Дж. Иммунол . 33 (8): 2069–73. дои : 10.1002/eji.200324133 . ПМИД 12884279 . S2CID 32059768 .
- ^ Казали П., Зан Х (2004). «Переключение классов и транслокация Myc: как ломается ДНК?» . Нат. Иммунол . 5 (11): 1101–3. дои : 10.1038/ni1104-1101 . ПМЦ 4625794 . ПМИД 15496946 .
- ^ Ставнезер Дж., Гикема Дж.Э., Шрейдер К.Э. (2008). «Механизм и регуляция рекомбинации переключателей классов» . Анну. Преподобный Иммунол . 26 : 261–292. doi : 10.1146/annurev.immunol.26.021607.090248 . ПМК 2707252 . ПМИД 18370922 .
- ^ Либер М.Р., Ю.К., Рагхаван С.К. (2006). «Роль негомологичного соединения концов ДНК, рекомбинации V (D) J и рекомбинации переключения классов в хромосомных транслокациях». Восстановление ДНК (Амст.) . 5 (9–10): 1234–45. дои : 10.1016/j.dnarep.2006.05.013 . ПМИД 16793349 .
- ^ Ян КТ, Бобойла С, Соуза Е.К., Франко С., Хикернелл Т.Р., Мерфи М., Гумасте С., Гейер М., Заррин А.А., Манис Дж.П., Раевски К., Альт Ф.В. (2007). «Переключение класса IgH и транслокации используют надежный неклассический путь соединения концов». Природа . 449 (7161): 478–82. Бибкод : 2007Natur.449..478Y . дои : 10.1038/nature06020 . ПМИД 17713479 . S2CID 4341381 .
- ^ Рейно С., Дельпи Л., Флёри Л., Дужье Х.Л., Сирак С., Конье М. (май 2005 г.). «Межаллельная рекомбинация переключения классов вносит значительный вклад в переключение классов в В-клетках мыши» . Журнал иммунологии . 174 (10): 6176–83. дои : 10.4049/jimmunol.174.10.6176 . ПМИД 15879114 .
- ^ Лаффлер Б., Барде С.М., Гаро А., Брус М., Байле А., Конье М. (2014). «Гены иммуноглобулинов подвергаются законной репарации в В-клетках человека не только после цис-, но и частой рекомбинации транс-класса» . Гены и иммунитет . 15 (5): 341–6. дои : 10.1038/gen.2014.25 . ПМИД 24848929 .
- ^ Джейнвей Калифорния младший; Трэверс П; Уолпорт М; Шломчик М.Ю. (2001). Иммунобиология (5-е изд.). Издательство «Гирлянда». ISBN 978-0-8153-3642-6 . (через книжную полку NCBI) .
- ^ Мужчина Д., Бростофф Дж., Рот Д.Б., Ройтт I (2006). Иммунология, 7-е изд. Филадельфия: Мосби Эльзевир, ISBN 978-0-323-03399-2 (пбк.)
- ^ Шпараго, Н.; Желазовский П.; Джин, Л.; Макинтайр, ТМ; Стубер, Э.; Песанья, LM; Кери, MR; Монд, Джей-Джей; Макс, Э.Э. (1 мая 1996 г.). «IL-10 избирательно регулирует переключение изотипа мышиного Ig» . Международная иммунология . 8 (5): 781–790. дои : 10.1093/интимм/8.5.781 . ISSN 0953-8178 . ПМИД 8671667 .
- ^ Бриер Ф., Серве-Дельпра С., Бридон Ж.М., Сен-Реми Ж.М., Баншеро Ж. (февраль 1994 г.). «Человеческий интерлейкин 10 индуцирует наивные поверхностные В-клетки иммуноглобулина D+ (sIgD+) к секреции IgG1 и IgG3» . Журнал экспериментальной медицины . 179 (2): 757–62. дои : 10.1084/jem.179.2.757 . ПМК 2191366 . ПМИД 8294883 .
- ^ Малисан Ф, Бриер Ф, Брайдон Дж.М., Хариндранат Н., Миллс ФК, Макс Э.Э., Баншеро Дж., Мартинес-Вальдес Х. (март 1996 г.). «Интерлейкин-10 индуцирует рекомбинацию переключения изотипа иммуноглобулина G в CD40-активированных наивных В-лимфоцитах человека» . Журнал экспериментальной медицины . 183 (3): 937–47. дои : 10.1084/jem.183.3.937 . ПМК 2192363 . ПМИД 8642297 .
- ^ Пино Э., Марке М., Фиансетт Р., Перон С., Винсент-Фабер С., Денизо Ю., Конье М. (2011). Регуляторная область 3' локуса IgH: дергая за ниточки сзади . Достижения иммунологии. Том. 110. стр. 27–70. дои : 10.1016/B978-0-12-387663-8.00002-8 . ISBN 978-0-12-387663-8 . ПМИД 21762815 .
- ^ Перон С., Лаффлер Б., Дени-Лагаш Н., Кук-Моро Дж., Тэнгли А., Дельпи Л., Денизо Ю., Пино Э., Конье М. (май 2012 г.). «Удаление, вызванное AID, вызывает самоубийственную рекомбинацию локуса тяжелой цепи иммуноглобулина в B-клетках». Наука . 336 (6083): 931–4. Бибкод : 2012Sci...336..931P . дои : 10.1126/science.1218692 . ПМИД 22539552 . S2CID 1513560 .
Внешние ссылки
[ редактировать ]- Иммуноглобулин+класс+переключение Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)