CD40 (белок)
| CD40 | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | CD40 , Bp50, CDW40, TNFRSF5, p50, CD40 (белок), молекула CD40 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 109535 ; МГИ : 88336 ; Гомологен : 954 ; Генные карты : CD40 ; ОМА : CD40 - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||




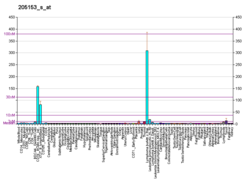
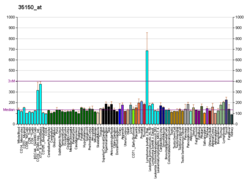
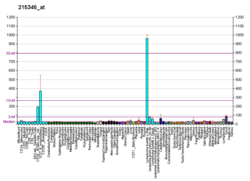
Кластер дифференцировки 40. CD40 представляет собой трансмембранный белок типа I, обнаруженный на антигенпрезентирующих клетках и необходимый для их активации. Связывание CD154 ( CD40L ) на Т H -клетках с CD40 активирует антигенпредставляющие клетки и индуцирует множество последующих эффектов.
Активированные CD4+ Т-клетки в первую очередь экспонируют свой лиганд CD154 на антигенпрезентирующие клетки, включая дендритные клетки (ДК), В-клетки, макрофаги, классические и неклассические моноциты, на различные неиммунные клетки, включая тромбоциты и эндотелиальные клетки, а также на некоторые виды опухолевых клеток. [ 5 ]
Мутации, затрагивающие этот ген, являются причиной аутосомно-рецессивного иммунодефицита гипер-IgM.
Открытие
[ редактировать ]В период с конца 1950-х по середину 1980-х годов несколько иммунологических лабораторий начали использовать новую гибридомную технологию для разработки моноклональных антител (мАт) и определения рецепторов, экспрессируемых на разных стадиях дифференцировки гемопоэтических клеток . Целью этих экспериментов была идентификация дифференцировочных антигенов, которые можно было бы использовать для описания стадий дифференцировки лимфоцитов и различных функциональных субпопуляций клеток. В ходе этих экспериментов было разработано несколько моноклональных антител против белка CD40, поверхностного рецептора В-клеток , который можно поликлонально активировать связывающим лигандом. Со временем были обнаружены многие особенности и цели сигнального пути CD40, включая открытие лиганда CD40 (CD154), молекулы на поверхности Т-клеток , которая способна индуцировать контактно-зависимую дифференцировку В-клеток. [ 6 ]
Функция
[ редактировать ]Белковый рецептор, кодируемый этим геном, является членом суперсемейства рецепторов TNF . Было обнаружено, что этот рецептор играет важную роль в опосредовании широкого спектра иммунных и воспалительных реакций, включая Т-клеточно-зависимое переключение класса иммуноглобулина, развитие В-клеток памяти и образование зародышевых центров . [ 7 ] AT-крючка Сообщается, что фактор транскрипции AKNA координирует экспрессию этого рецептора и его лиганда, что может быть важно для гомотипических клеточных взаимодействий. Установлено, что взаимодействие этого рецептора и его лиганда необходимо для бета-амилоидом индуцированной активации микроглии, , и, таким образом, считается ранним событием в болезни Альцгеймера патогенезе . Сообщалось о двух альтернативно сплайсированных вариантах транскрипта этого гена, кодирующих разные изоформы. [ 8 ]
Специфическое воздействие на клетки
[ редактировать ]В макрофагах первичным сигналом активации является IFN-γ от Т Th1 -клеток CD4 типа . Вторичным сигналом является CD40L (CD154) на клетке Th1 , который связывает CD40 на поверхности клетки-макрофага. В результате макрофаг экспрессирует на своей поверхности больше рецепторов CD40 и TNF , что помогает повысить уровень активации. Увеличение активации приводит к индукции в макрофагах мощных микробицидных веществ, включая активные формы кислорода и оксид азота , что приводит к уничтожению проглоченного микроба.
может В-клетка представлять антигены хелперам Т- . Если активированная Т-клетка распознает пептид, представленный В-клеткой, CD40L на Т-клетке связывается с рецептором CD40 В-клетки, вызывая активацию В-клетки. Т-клетка также производит IL-2 , который напрямую влияет на B-клетки. В результате этой чистой стимуляции В-клетка может подвергаться делению, антител переключению изотипа и дифференцировке в плазматические клетки . Конечным результатом являются В-клетки, способные массово продуцировать специфические антитела против антигенной мишени. Ранними доказательствами этих эффектов было то, что у мышей с дефицитом CD40 или CD154 происходит незначительное переключение классов или образование зародышевых центров . [ 9 ] и иммунные реакции сильно подавляются.
Экспрессия CD40 разнообразна. CD40 конститутивно экспрессируется антигенпрезентирующими клетками, включая дендритные клетки , В-клетки и макрофаги . Он также может экспрессироваться эндотелиальными клетками , гладкомышечными клетками , фибробластами и эпителиальными клетками. [ 10 ] В соответствии с его широко распространенной экспрессией на нормальных клетках, CD40 также экспрессируется в широком спектре опухолевых клеток, включая неходжкинские и ходжкинские лимфомы, миелому и некоторые карциномы, включая носоглотку, мочевой пузырь, шейку матки, почки и яичники. CD40 также экспрессируется на предшественниках В-клеток в костном мозге, и есть некоторые доказательства того, что взаимодействия CD40-CD154 могут играть роль в контроле В-клеточного кроветворения . [ 11 ]
Взаимодействия
[ редактировать ]Было показано, что CD40 (белок) взаимодействует с TRAF2 , [ 12 ] [ 13 ] [ 14 ] ТРАФ3 , [ 13 ] [ 15 ] [ 16 ] [ 17 ] ТРАФ6 , [ 13 ] [ 17 ] ТРАФ5 [ 13 ] [ 18 ] и ТТРАП . [ 19 ] Оставшийся член семейства TRAF4, а именно TRAF4 , положительно регулирует передачу сигналов CD40, но взаимодействует с CD40 опосредованно. [ 20 ]
CD40 также взаимодействует с CD40L из-за роли CD40 в стимуляции иммунных синапсов , когда это взаимодействие происходит с CD40L, активирующим дендритные клетки для активации антигенспецифических Т-клеток. Это происходит за счет усиления экспрессии молекул главного комплекса гистосовместимости , повышения экспрессии костимулирующих молекул CD86/CD80 и усиления лигандов суперсемейства TNF на поверхности дендритных клеток, а также секреции интерлейкина-12 (IL-12), который способствует CD8+. Активация Т-клеток . Более того, взаимодействия CD40/CD40L провоцируют противоопухолевые иммунные ответы за счет увеличения гибели опухолевых иммуногенных клеток (ICD), активации APC, иммуногенности опухоли за счет активации молекул главного комплекса гистосовместимости (MHC), продукции провоспалительных факторов, совместной стимуляции CD4 + и CD8 + Т-клеток, и чувствительность опухолевых клеток к лизису Т-клеток. Кроме того, ось CD40/CD40LG важна для обновления иммунных клеток и гомеостаза в нормальных условиях. Предполагается, что самая тесная связь пролиферации клеток наблюдается с CD40LG и проапоптотическим маркером. BAX также играет решающую роль в стимулировании активации и пролиферации В-клеток, иммунных синапсов ВТ-клеток и презентации антигена. [ 21 ] [ 5 ]
CD40 как мишень для лечения рака
[ редактировать ]Молекула CD40 является потенциальной мишенью для иммунотерапии рака . Моноклональные антитела против CD40 могут способствовать уничтожению раковых клеток эффекторными клетками . Аналогичным образом, лигирование CD40 может привести к гибели некоторых опухолевых клеток, поскольку оно экспрессируется при всех лимфоидных злокачественных новообразованиях и в ряде карцином . [ 6 ] Существует ряд завершенных и продолжающихся клинических испытаний с использованием агонистических моноклональных антител против CD40 для индукции противоопухолевого Т-клеточного ответа посредством активации дендритных клеток. За последние 20 лет были разработаны и оценены в клинических испытаниях многочисленные моноклональные антитела человека к CD40, что способствовало повышению вариабельности моделей рака на животных. Агонистические анти-CD-40-Ab созданы для имитации CD40L путем перекрестного связывания CD40 и, таким образом, способствуют созреванию ДК и усилению их способности к презентации антигена. Это приводит к увеличению количества цитотоксических Т-клеток, специфичных к опухолевому антигену, что может привести к эрадикации опухоли. С другой стороны, доклиническая эффективность еще не была проверена в клинических условиях, и ни одно из этих моноклональных антител не продвинулось дальше ранних этапов тестирования. Из-за токсичности использование моноклональных антител к CD40 было ограничено субоптимальными дозами, что приводило к неадекватной иммунной активации и противоопухолевой активности. [ 5 ] Совсем недавно было показано, что агонистическая терапия CD40 снижает цитотоксичность Т-клеток на доклинических моделях глиомы и фактически влияет на эффективность блокады иммунных контрольных точек. Вероятно, это связано с высокой мутационной нагрузкой, которую демонстрируют большинство этих моделей, что заставляет их лучше реагировать на блокаду иммунных контрольных точек, чем глиома человека, но, тем не менее, это актуальная информация для исследований в области иммуномодулирующей терапии. [ 22 ]
Гипериммунодефицит Ig-M и CD40
[ редактировать ]Синдром гипер-IgM — это первичное иммунодефицитное заболевание, характеризующееся повышением уровня иммуноглобулина (Ig) M в сыворотке крови и снижением уровней IgG, IgA и IgE. CD40 участвует в развитии синдрома гипер-IgM, поскольку он служит костимулирующей молекулой при активации дифференцировки В-клеток, которые играют ключевую роль в выработке иммуноглобулинов. При синдроме гипер-IgM мутации в генах, участвующих в передаче сигналов CD40, приводят к нарушению активации и дифференцировки В-клеток, что приводит к увеличению продукции IgM и снижению продукции других иммуноглобулинов. В результате люди с синдромом гипер-IgM восприимчивы к широкому спектру инфекций и имеют повышенный риск аутоиммунных заболеваний и рака. В настоящее время лечение синдрома гипер-IgM включает замену недостающих иммуноглобулинов, а также другие методы лечения для укрепления иммунной системы и предотвращения инфекций. Исследования продолжаются, чтобы лучше понять роль CD40 в синдроме гипер-IgM и разработать новые методы лечения этого заболевания. [ нужна ссылка ] [ 23 ]
CD40 и разработка лекарств
[ редактировать ]CD40 является многообещающей мишенью для разработки лекарств для лечения различных заболеваний, включая рак, аутоиммунные заболевания и хроническое воспаление. Воздействуя на CD40, можно модулировать иммунный ответ и повышать способность организма бороться с болезнями. Например, препараты, блокирующие передачу сигналов CD40, оказались многообещающими в лечении аутоиммунных заболеваний, таких как ревматоидный артрит, путем подавления сверхактивного иммунного ответа. С другой стороны, препараты, активирующие передачу сигналов CD40, показали эффективность в лечении рака, усиливая иммунный ответ против опухолевых клеток. CD40 также играет роль в развитии хронического воспаления, а воздействие на CD40 с помощью лекарств потенциально может помочь в лечении таких заболеваний, как болезнь Крона и язвенный колит. В целом CD40 представляет собой многообещающую цель для разработки лекарств для лечения широкого спектра заболеваний. [ 24 ] [ 25 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000101017 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000017652 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б с Саломон Р., Дахан Р. (2022). «Агонистические антитела к CD40 следующего поколения для иммунотерапии рака» . Границы в иммунологии . 13 : 940674. дои : 10.3389/fimmu.2022.940674 . ПМК 9326085 . ПМИД 35911742 .
- ^ Jump up to: а б Кларк Э.А. (2014). «Краткая история поверхностной молекулы CD40, ассоциированной с B-клетками» . Границы в иммунологии . 5 : 472. дои : 10.3389/fimmu.2014.00472 . ПМЦ 4179537 . ПМИД 25324844 .
- ^ Гревал И.С., Флавелл Р.А. (1998). «CD40 и CD154 в клеточном иммунитете». Ежегодный обзор иммунологии . 16 : 111–135. doi : 10.1146/annurev.immunol.16.1.111 . ПМИД 9597126 .
- ^ «Ген Энтрез: молекула CD40 CD40, член 5 суперсемейства рецепторов TNF» .
- ^ Кавабе Т., Нака Т., Ёсида К., Танака Т., Фудзивара Х., Суэмацу С. и др. (июнь 1994 г.). «Иммунные реакции у мышей с дефицитом CD40: нарушение переключения классов иммуноглобулинов и образование зародышевых центров». Иммунитет . 1 (3): 167–178. дои : 10.1016/1074-7613(94)90095-7 . ПМИД 7534202 .
- ^ Хацигеоргиу А, Либери М, Хатзилимперис Г, Незос А, Кампер Э (2009). «Передача сигналов CD40/CD40L и ее значение для здоровья и болезней». Биофакторы . 35 (6): 474–483. дои : 10.1002/биоф.62 . ПМИД 19904719 . S2CID 22911861 .
- ^ Карлринг Дж., Альтахер Х.М., Кларк С., Чен Х., Латимер С.Л., Дженнер Т. и др. (май 2011 г.). «Взаимодействие CD154-CD40 в контроле кроветворения В-клеток мышей» . Журнал биологии лейкоцитов . 89 (5): 697–706. дои : 10.1189/jlb.0310179 . ПМЦ 3382295 . ПМИД 21330346 .
- ^ МакВиртер С.М., Пуллен С.С., Холтон Дж.М., Крут Дж.Дж., Кери М.Р., Альбер Т. (июль 1999 г.). «Кристаллографический анализ распознавания и передачи сигналов CD40 с помощью TRAF2 человека» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (15): 8408–8413. Бибкод : 1999PNAS...96.8408M . дои : 10.1073/pnas.96.15.8408 . ЧВК 17529 . ПМИД 10411888 .
- ^ Jump up to: а б с д Цукамото Н., Кобаяши Н., Адзума С., Ямамото Т., Иноуэ Дж. (февраль 1999 г.). «Два по-разному регулируемых пути активации ядерного фактора каппаВ, запускаемые цитоплазматическим хвостом CD40» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (4): 1234–1239. Бибкод : 1999PNAS...96.1234T . дои : 10.1073/pnas.96.4.1234 . ПМК 15446 . ПМИД 9990007 .
- ^ Малинин Н.Л., Болдин М.П., Коваленко А.В., Уоллак Д. (февраль 1997 г.). «MAP3K-родственная киназа, участвующая в индукции NF-kappaB с помощью TNF, CD95 и IL-1». Природа . 385 (6616): 540–544. дои : 10.1038/385540a0 . ПМИД 9020361 . S2CID 4366355 .
- ^ Ху Х.М., О'Рурк К., Богуски М.С., Диксит В.М. (декабрь 1994 г.). «Новый белок RING-пальца взаимодействует с цитоплазматическим доменом CD40» . Журнал биологической химии . 269 (48): 30069–30072. дои : 10.1016/S0021-9258(18)43772-6 . ПМИД 7527023 .
- ^ Ni CZ, Welsh K, Leo E, Chiou CK, Wu H, Reed JC и др. (сентябрь 2000 г.). «Молекулярная основа передачи сигналов CD40, опосредованной TRAF3» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (19): 10395–10399. Бибкод : 2000PNAS...9710395N . дои : 10.1073/pnas.97.19.10395 . ПМК 27035 . ПМИД 10984535 .
- ^ Jump up to: а б Рой Н., Деверо К.Л., Такахаши Р., Салвесен Г.С., Рид Дж.К. (декабрь 1997 г.). «Белки c-IAP-1 и c-IAP-2 являются прямыми ингибиторами специфических каспаз» . Журнал ЭМБО . 16 (23): 6914–6925. дои : 10.1093/emboj/16.23.6914 . ПМК 1170295 . ПМИД 9384571 .
- ^ Исида Т.К., Тодзё Т., Аоки Т., Кобаяши Н., Охиши Т., Ватанабэ Т. и др. (сентябрь 1996 г.). «TRAF5, новый белок семейства факторов, ассоциированный с рецептором фактора некроза опухоли, опосредует передачу сигналов CD40» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (18): 9437–9442. Бибкод : 1996PNAS...93.9437I . дои : 10.1073/pnas.93.18.9437 . ПМК 38446 . ПМИД 8790348 .
- ^ Пайп С., Деклерк В., Ибрагими А., Михилс С., Ван Ритчотен Дж.Г., Девульф Н. и др. (июнь 2000 г.). «TTRAP, новый белок, который связывается с CD40, рецептором фактора некроза опухоли (TNF)-75 и факторами, связанными с рецептором TNF (TRAF), и ингибирует активацию ядерного фактора каппа B» . Журнал биологической химии . 275 (24): 18586–18593. дои : 10.1074/jbc.M000531200 . PMID 10764746 .
- ^ Шарма С., Павласова Г.М., Седа В., Черна К.А., Воджакова Е., Филип Д. и др. (май 2021 г.). «МиР-29 модулирует передачу сигналов CD40 при хроническом лимфоцитарном лейкозе, нацеливаясь на TRAF4: ось, на которую влияют ингибиторы BCR» . Кровь . 137 (18): 2481–2494. дои : 10.1182/blood.2020005627 . ПМЦ 7610744 . ПМИД 33171493 .
- ^ Ян С., Ричмонд А (ноябрь 2021 г.). «Спрятаться в темноте: характеристика экспрессии панрака и клиническая значимость CD40 для терапии блокады иммунных контрольных точек» . Молекулярный рак . 20 (1): 146. дои : 10.1186/s12943-021-01442-3 . ПМЦ 8582157 . ПМИД 34758832 .
- ^ ван Хурен Л., Ваккаро А., Рамачандран М., Вазайос К., Либард С., ван де Валле Т. и др. (июль 2021 г.). «Агонистическая терапия CD40 индуцирует третичные лимфоидные структуры, но ухудшает реакцию на блокаду контрольных точек при глиоме» . Природные коммуникации . 12 (1): 4127. Бибкод : 2021NatCo..12.4127V . дои : 10.1038/s41467-021-24347-7 . ПМЦ 8257767 . ПМИД 34226552 .
- ^ Яздани Р., Фекрванд С., Шахкарами С., Азизи Г., Моаззами Б., Аболхассани Х. и Агамохаммади А. (2019). Синдромы гиперIgM: эпидемиология, патогенез, клинические проявления, диагностика и лечение. Клиническая иммунология (Орландо, Флорида), 198, 19–30. https://doi.org/10.1016/j.clim.2018.11.007
- ^ Сингх Т., Фатехи Хасанабад М., Фатехи Хасанабад А. (декабрь 2021 г.). «Немелкоклеточный рак легкого: новые молекулярно-таргетные и иммунотерапевтические агенты». Биохимика и биофизика Acta. Обзоры на Рак . 1876 (2): 188636. doi : 10.1016/j.bbcan.2021.188636 . ПМИД 34655692 . S2CID 239010744 .
- ^ Саломон Р., Дахан Р. (13 июля 2022 г.). «Агонистические антитела к CD40 следующего поколения для иммунотерапии рака» . Границы в иммунологии . 13 : 940674. дои : 10.3389/fimmu.2022.940674 . ПМК 9326085 . ПМИД 35911742 .
Внешние ссылки
[ редактировать ]- человека Расположение генома CD40 и страница сведений о гене CD40 в браузере генома UCSC .
- PDBe-KB предоставляет обзор всей информации о структуре, доступной в PDB для члена 5 суперсемейства рецепторов фактора некроза опухоли человека (CD40).
Дальнейшее чтение
[ редактировать ]- Пархэм П. (2004). Иммунная система (2-е изд.). Гирляндная наука. стр. 169–173 . ISBN 978-0-8153-4093-5 .
- Ван Дж.Х., Чжан Ю.В., Чжан П., Дэн Б.К., Дин С., Ван З.К. и др. (сентябрь 2013 г.). «Лиганд CD40 как потенциальный биомаркер атеросклеротической нестабильности» . Неврологические исследования . 35 (7): 693–700. дои : 10.1179/1743132813Y.0000000190 . ПМК 3770830 . ПМИД 23561892 .
- Баншеро Дж. , Базан Ф., Бланшар Д., Бриер Ф., Галицци Дж. П., ван Кутен С. и др. (1994). «Антиген CD40 и его лиганд». Ежегодный обзор иммунологии . 12 : 881–922. дои : 10.1146/annurev.iy.12.040194.004313 . ПМИД 7516669 .
- ван Кутен С., Баншеро Дж. (январь 2000 г.). «Лиганд CD40-CD40». Журнал биологии лейкоцитов . 67 (1): 2–17. дои : 10.1002/jlb.67.1.2 . ПМИД 10647992 . S2CID 35592719 .
- Шаттнер Э.Дж. (май 2000 г.). «Лиганд CD40 в патогенезе и терапии ХЛЛ». Лейкемия и лимфома . 37 (5–6): 461–472. дои : 10.3109/10428190009058499 . ПМИД 11042507 . S2CID 39398949 .
- Бхушан А., Кови Л.Р. (2002). «Взаимодействие CD40:CD40L при Х-связанных и не-Х-связанных синдромах гипер-IgM». Иммунологические исследования . 24 (3): 311–324. дои : 10.1385/IR:24:3:311 . ПМИД 11817328 . S2CID 19537892 .
- Ченг Г., Шенбергер С.П. (2002). «Передача сигналов CD40 и аутоиммунитет». Пути передачи сигнала при аутоиммунитете . Современные направления в области аутоиммунитета. Том. 5. С. 51–61. дои : 10.1159/000060547 . ISBN 978-3-8055-7308-5 . ПМИД 11826760 .
- Даллман С., Джонсон П.В., Пакхэм Дж. (январь 2003 г.). «Дифференциальная регуляция выживания клеток с помощью CD40». Апоптоз . 8 (1): 45–53. дои : 10.1023/А:1021696902187 . ПМИД 12510151 . S2CID 22461134 .
- О'Салливан Б., Томас Р. (июль 2003 г.). «Последние достижения в области роли CD40 и дендритных клеток в иммунитете и толерантности». Современное мнение в гематологии . 10 (4): 272–278. дои : 10.1097/00062752-200307000-00004 . ПМИД 12799532 . S2CID 43043879 .
- Бенвенист Э.Н., Нгуен В.Т., Весеманн Д.Р. (январь 2004 г.). «Молекулярная регуляция экспрессии гена CD40 в макрофагах и микроглии». Мозг, поведение и иммунитет . 18 (1): 7–12. дои : 10.1016/j.bbi.2003.09.001 . ПМИД 14651941 . S2CID 8081107 .
- Сюй Ю, Сун Г (2005). «Роль взаимодействия CD40-CD154 в клеточной иммунорегуляции». Журнал биомедицинской науки . 11 (4): 426–438. дои : 10.1159/000077892 . ПМИД 15153777 . S2CID 202658036 .
- Контин С., Кузи Л., Моро Дж. Ф., Дешане-Мервиль Дж., Мервилл П. (2004). «[Иммунная дисфункция пациентов с уремией: потенциальная роль растворимой формы CD40]». Нефрология . 25 (4): 119–126. ПМИД 15291139 .