Горизонтальный перенос генов в эволюции

Эта статья написана как личное размышление, личное эссе или аргументативное эссе , в котором излагаются личные чувства редактора Википедии или представлен оригинальный аргумент по определенной теме. ( Ноябрь 2023 г. ) |
Горизонтальный перенос генов ( HGT ) относится к переносу генов между отдаленными ветвями древа жизни . В ходе эволюции он может шифровать информацию, необходимую для реконструкции филогении организмов и того, как они связаны друг с другом.
HGT также может помочь ученым реконструировать и датировать древо жизни, поскольку перенос генов может использоваться в качестве филогенетического маркера или как доказательство современности организмов-донора и реципиента, а также как след вымершего биоразнообразия.
ГГТ случается очень редко – на уровне отдельного организма такое событие маловероятно. Однако в более широком масштабе эволюционной истории эти события происходят с некоторой регулярностью. С одной стороны, это заставляет биологов отказаться от использования отдельных генов в качестве хороших маркеров истории жизни. С другой стороны, это представляет собой почти неиспользованный большой источник информации о прошлом.
Три сферы жизни
[ редактировать ]Три основные ранние ветви древа жизни интенсивно изучались микробиологами, поскольку первыми организмами были микроорганизмы. Микробиологи (во главе с Карлом Везе ) ввели термин домен для трех основных ветвей этого дерева, где домен — это филогенетический термин, аналогичный по значению биологическому царству . Для реконструкции этого древа жизни оказалась полезной последовательность генов, кодирующая малую субъединицу рибосомальной РНК (SSU рРНК, 16s рРНК ), и дерево (как показано на рисунке) во многом полагается на информацию от этого единственного гена.
Эти три домена жизни представляют собой основные эволюционные линии ранней клеточной жизни и в настоящее время включают бактерии , археи (одноклеточные организмы, внешне похожие на бактерии) и эукарии . К эукариям относятся только организмы, имеющие четко выраженное ядро, такие как грибы , простейшие и все организмы царства растений и животных (см. рисунок).
Геном, наиболее часто используемым для построения филогенетических отношений у микроорганизмов , является ген малой субъединицы рибосомальной РНК, поскольку его последовательности имеют тенденцию сохраняться среди членов с близкими филогенетическими расстояниями, но при этом достаточно изменчивы, чтобы различия можно было измерить. [ 1 ] рРНК SSU как мера эволюционных расстояний была впервые использована Карлом Везе при формулировании первого современного «дерева жизни», и его результаты побудили его предложить архей в качестве третьего домена жизни . Однако недавно появились утверждения, что гены рРНК SSU могут переноситься и горизонтально. [ 2 ] Хотя это может быть редко, такая возможность заставляет тщательно изучить достоверность филогенетических деревьев, основанных на SSU рРНК.
Недавние открытия «безудержного» ГПГ у микроорганизмов и обнаружение горизонтального движения даже генов небольшой субъединицы рибосомальной РНК заставили биологов усомниться в точности, по крайней мере, ранних ветвей дерева, и даже усомниться в достоверности деревья как полезные модели того, как происходит ранняя эволюция. [ 3 ] Фактически, считается, что ранняя эволюция произошла из сообщества предшественников , способных обмениваться большими молекулами, когда HGT был стандартом. Этот латеральный перенос генов произошел и за пределами дарвиновского порога , после того как была установлена наследственность или вертикальный перенос генов. [ 4 ] [ 5 ]
«Сравнение последовательностей предполагает недавний горизонтальный перенос многих генов между различными видами , в том числе через границы филогенетических «доменов». Таким образом, определение филогенетической истории вида не может быть сделано окончательно путем определения эволюционных деревьев для отдельных генов». [ 6 ] Таким образом, HGT является потенциальным искажающим фактором при построении филогенетических деревьев на основе последовательности одного гена . Например, если две отдаленно родственные бактерии обменялись геном, филогенетическое древо, включающее эти виды, покажет, что они тесно связаны, даже если большинство других генов существенно разошлись. По этой причине важно использовать другую информацию для вывода о филогении, например наличие или отсутствие генов, или, что чаще, включать как можно более широкий спектр генов для анализа. [ 6 ]
Считается, что более ранние HGT произошли. Считается, что первый универсальный общий предок (FUCA), самый ранний предок последнего общего предка всего живого (LUCA), имел других потомков, имевших свои собственные линии. [ 7 ] [ 8 ] Считается, что эти ныне вымершие сестринские линии LUCA, происходящие от FUCA, горизонтально перенесли некоторые из своих генов в геном ранних потомков LUCA. [ 8 ]
Выбор метафоры: дерево, сеть, паутина или кольцо.
[ редактировать ]В своей статье «Искоренение Древа Жизни » У. Форд Дулиттл обсуждает Последнего Вселенского Общего Предка – корень Древа Жизни – и проблемы, связанные с этой концепцией, возникающие из-за HGT. [ 9 ] Он описывает микроорганизм Archaeoglobus fulgidus как аномалию по отношению к филогенетическому дереву, основанному на коде фермента HMGCoA-редуктазы : этот организм определенно является археем, со всеми клеточными липидами и механизмом транскрипции, ожидаемыми от архей, но его гены HMGCoA бактериального происхождения. В статье Дулитл говорит, что, хотя в настоящее время широко признано, что митохондрии эукариот произошли от альфа-протеобактериальных клеток, а хлоропласты произошли от проглоченных цианобактерий , [ 9 ]
«... уже нельзя с уверенностью предполагать, что это были единственные латеральные переносы генов, которые произошли после возникновения первых эукариот. Только у более поздних, многоклеточных эукариот мы знаем об определенных ограничениях на горизонтальный обмен генами, таких как появление разделенных ( и защищенные) половые клетки ...
Если бы не было никакого латерального переноса генов, все эти отдельные генные деревья имели бы одинаковую топологию (один и тот же порядок ветвления), и все предковые гены в корне каждого дерева присутствовали бы у последнего универсального общего предка, единственная древняя клетка. Но обширный перенос означает, что ни то, ни другое не так: деревья генов будут различаться (хотя многие из них будут иметь области схожей топологии) и никогда не будет ни одной клетки, которую можно было бы назвать последним универсальным общим предком...» [ 9 ]
Дулиттл предположил, что универсальный общий предок не мог быть одним конкретным организмом, а должен был быть рыхлым, разнообразным скоплением примитивных клеток, которые эволюционировали вместе. Эти ранние клетки, каждая из которых имела относительно небольшое количество генов, во многом различались и свободно меняли свои гены . В конце концов, из этих эклектичных клеток произошли три области жизни, какими мы их знаем сегодня: бактерии , археи и эукариоты . Эти домены теперь заметно различны, поскольку большая часть переноса генов, который все еще происходит, происходит внутри этих доменов, а не между ними. Биолог Питер Гогартен подкрепил эти аргументы и предположил, что метафора дерева не соответствует данным недавних исследований генома и что вместо этого биологам следует использовать «метафору мозаики для описания различных историй, объединенных в отдельные геномы и использование [ ] метафора сети для визуализации богатого обмена и совместных эффектов HGT среди микробов». [ 10 ]
Разрешение неопределенности с помощью филогеномики
[ редактировать ]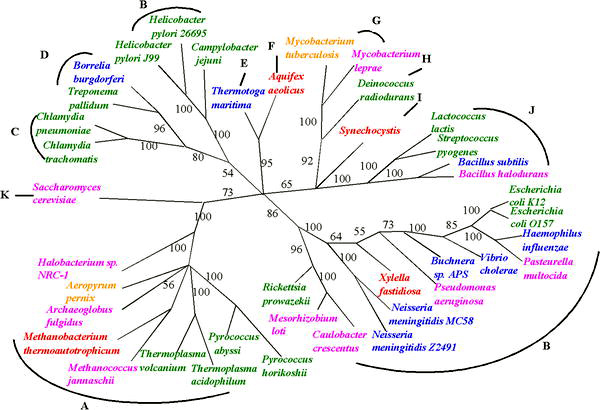
Несмотря на неопределенности в реконструкции филогении до начала жизни, прогресс достигается в реконструкции древа жизни перед лицом неопределенностей, вызванных HGT. Неопределенность любого предполагаемого филогенетического дерева, основанного на одном гене, можно решить, используя несколько общих генов или даже данные целых геномов. [ 12 ] Один из таких подходов, иногда называемый «мультилокусным типированием», использовался для построения филогенетических деревьев организмов, которые обмениваются генами, таких как бактерии менингита. [ 13 ]
Джонатан Эйзен и Клэр Фрейзер отметили, что:
«При построении древа жизни анализ целых геномов начал дополнять, а в некоторых случаях и улучшать исследования, ранее проводившиеся с одним или несколькими генами. Например, недавние исследования полных бактериальных геномов показали, что гипертермофильные виды не являются глубоко ветвящимися; если это правда, то это ставит под сомнение идею о том, что первые формы жизни были термофилами. Анализ генома эукариотического паразита Encephalitozoon cuniculi подтверждает предположения о том, что группа Microsporidia не является глубоковетвистыми протистами. Анализ генома может даже помочь разрешить взаимоотношения внутри видов, например, предоставив новые генетические маркеры для популяционно-генетических исследований бактерий, вызывающих сибирскую язву или туберкулез. Во всех этих исследованиях речь идет о дополнительных данных, предоставляемых полным геномом. Последовательность, которая позволяет отделить филогенетический сигнал от шума. Это не значит, что древо жизни теперь решено — мы выбрали лишь небольшое количество геномов, и многие группы еще не затронуты». [ 14 ]
Эти подходы позволяют оценить относительную частоту ГПГ; наблюдавшиеся относительно низкие значения позволяют предположить, что «дерево» по-прежнему является действенной метафорой эволюции, но дерево украшено «паутиной» горизонтально передаваемых генов. Это основной вывод исследования 2005 года более 40 полных геномных последовательностей микробов, проведенного Фан Ге, Ли-Сан Ван и Чунхён Кимом . Они оценивают частоту событий HGT примерно в 2% основных генов на геном. [ 15 ] Подобные полногеномные подходы к оценке эволюции также позволяют добиться прогресса в идентификации очень ранних событий на древе жизни, например, предположение, что эукариоты возникли в результате слияния двух полных, но очень разных геномов прокариот: одного из бактерии и одного из архейной клетки. . [ 3 ]
слияния организмов Подобную гипотезу о происхождении сложных ядросодержащих клеток выдвинула Линн Маргулис, используя совершенно иные рассуждения о симбиозе бактерии и археи, возникшем в древнем консорциуме микробов. [ 16 ]
Использование в эволюционных исследованиях
[ редактировать ]Хотя HGT часто рассматривается как задача для реконструкции древа жизни, альтернативная точка зрения состоит в том, что, напротив, оно предоставляет дополнительную ценную информацию для его реконструкции.
Во-первых, для организма-реципиента HGT представляет собой мутацию ДНК , как и другие, и поэтому ее можно моделировать и использовать для реконструкции и укоренения деревьев. [ 17 ]
Во-вторых, необходимо, чтобы получатель гена, приобретенного с помощью HGT, жил в то же время или в более позднее время, что и донор. [ 18 ] В последствии появляется информация о сроках диверсификации HGT. [ 19 ] Это тем более примечательно, что основной обычный источник датирования в живом мире — летопись окаменелостей — отсутствует именно там, где HGT в изобилии, в микробном мире.
В-третьих, он предоставляет информацию об исчезнувшем биоразнообразии, поскольку перенос, скорее всего, происходит от вымерших видов. [ 20 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Вёзе С. и др. (1990). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . ПНАС США . 87 (12): 4576–9. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- Вёзе С., Фокс Дж. (1977). «Филогенетическая структура прокариотического домена: первичные царства» . ПНАС США . 74 (11): 5088–90. Бибкод : 1977PNAS...74.5088W . дои : 10.1073/pnas.74.11.5088 . ПМК 432104 . ПМИД 270744 .
- ^ Яп, WH; Чжан, З; Ван, Ю. (1999). «В геноме актиномицета Thermomonospora chromogena существуют различные типы оперонов рРНК, что свидетельствует о горизонтальном переносе всего оперона рРНК» . Журнал бактериологии . 181 (17): 5201–9. дои : 10.1128/JB.181.17.5201-5209.1999 . ПМК 94023 . ПМИД 10464188 .
- ^ Перейти обратно: а б Симонсон, AB; Сервин, Дж.А.; Скопхаммер, Р.Г.; Гербольд, CW; Ривера, MC; Лейк, Дж.А. (2005). «Расшифровка геномного древа жизни» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (Приложение 1): 6608–13. Бибкод : 2005PNAS..102.6608S . дои : 10.1073/pnas.0501996102 . ПМЦ 1131872 . ПМИД 15851667 .
- ^ Вёзе, К. (9 июня 1998 г.). «Всемирный предок» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (12): 6854–6859. Бибкод : 1998PNAS...95.6854W . дои : 10.1073/pnas.95.12.6854 . ISSN 0027-8424 . ПМК 22660 . ПМИД 9618502 .
- ^ Вёзе, Карл Р. (25 июня 2002 г.). «Об эволюции клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (13): 8742–8747. Бибкод : 2002PNAS...99.8742W . дои : 10.1073/pnas.132266999 . ISSN 0027-8424 . ПМЦ 124369 . ПМИД 12077305 .
- ^ Перейти обратно: а б Горизонтальный перенос генов, штат Оклахома
- ^ Просдочими, Франциско; Хосе, Марко В.; де Фариас, Савио Торрес (2019), Понтаротти, Пьер (редактор), «Первый универсальный общий предок (FUCA) как самый ранний предок линии LUCA (последнего UCA)» , «Эволюция, происхождение жизни, концепции и методы» , Чам : Springer International Publishing, стр. 43–54, номер документа : 10.1007/978-3-030-30363-1_3 , ISBN. 978-3-030-30363-1 , S2CID 199534387 , получено 2 ноября 2023 г.
- ^ Перейти обратно: а б Харрис, Хью МБ; Хилл, Колин (2021). «Место вирусам на древе жизни» . Границы микробиологии . 11 . дои : 10.3389/fmicb.2020.604048 . ISSN 1664-302X . ПМЦ 7840587 . ПМИД 33519747 .
- ^ Перейти обратно: а б с Дулитл, WF (2000). «Искоренение древа жизни». Sci Am . 282 (2): 90–5. Бибкод : 2000SciAm.282b..90D . doi : 10.1038/scientificamerican0200-90 . ПМИД 10710791 .
- ^ Гогартен Дж. П. «Горизонтальный перенос генов - новая парадигма биологии». Кандидатская диссертация. Архивировано 21 июля 2012 г. в Wayback Machine.
- Жахыбаева О; Гогартен, JP (2004). «Кладогенез, слияние и эволюция трех областей жизни» (PDF) . Тенденции в генетике . 20 (4): 182–7. дои : 10.1016/j.tig.2004.02.004 . ПМИД 15041172 . Архивировано (PDF) из оригинала 26 марта 2009 г. Проверено 10 июля 2009 г.
- ^ Ге, Ф.; Ван, Л.; Ким, Дж. (2005). «Паутина жизни, раскрываемая с помощью оценок горизонтального переноса генов в масштабе генома» . ПЛОС Биология . 3 (10): е316. дои : 10.1371/journal.pbio.0030316 . ПМЦ 1233574 . ПМИД 16122348 .

- ^ Хенц, С.; Хьюсон, Д.; Ош, А.; Низельт-Струве, К.; Шустер, С. (2005). «Полногеномная филогения прокариот» . Биоинформатика . 21 (10): 2329–2335. doi : 10.1093/биоинформатика/bth324 . ПМИД 15166018 .
- Фитцпатрик, округ Колумбия; Лог, Мэн; Стаич, Дж. Э.; Батлер, Дж. (2006). «Филогения грибов, основанная на 42 полных геномах, полученных в результате супердерева и комбинированного анализа генов» . Эволюционная биология BMC . 6:99 . дои : 10.1186/1471-2148-6-99 . ПМК 1679813 . ПМИД 17121679 .
- Фитцпатрик, округ Колумбия; Лог, Мэн; Стаич, Дж. Э.; Батлер, Дж. (2006). «Филогения грибов, основанная на 42 полных геномах, полученных в результате супердерева и комбинированного анализа генов» . Эволюционная биология BMC . 6:99 . дои : 10.1186/1471-2148-6-99 . ПМК 1679813 . ПМИД 17121679 .
- ^ Урвин, Р; Мейден, MC (2003). «Мультилокусное типирование последовательностей: инструмент глобальной эпидемиологии». Тенденции Микробиол . 11 (10): 479–87. дои : 10.1016/j.tim.2003.08.006 . ПМИД 14557031 .
- Ян, Z (2002). «Оценка вероятности и Байеса размеров предковой популяции гоминоидов с использованием данных из нескольких локусов» . Генетика . 162 (4): 1811–23. дои : 10.1093/генетика/162.4.1811 . ПМЦ 1462394 . ПМИД 12524351 . Архивировано из оригинала 6 января 2009 г. Проверено 10 июля 2009 г.
- Дженнингс, ВБ; Эдвардс, С.В. (2005). «Специальная история австралийских травяных вьюрков (Poephila), выведенная на основе тридцати генных деревьев». Эволюция; Международный журнал органической эволюции . 59 (9): 2033–2047. дои : 10.1554/05-280.1 . ПМИД 16261740 . S2CID 18665436 .
- ^ Эйзен, Дж.; Фрейзер, К. (2003). «Филогеномика: пересечение эволюции и геномики». Наука . 300 (5626): 1706–1707. Бибкод : 2003Sci...300.1706E . дои : 10.1126/science.1086292 . ПМИД 12805538 . S2CID 42394233 .
- ^ Ге, Фан; Ван, Ли-Сан; Ким, Чунхён (2005). «Паутина жизни, раскрытая с помощью оценок горизонтального переноса генов в масштабе генома» . ПЛОС Биология . 3 (10): е316. дои : 10.1371/journal.pbio.0030316 . ПМЦ 1233574 . ПМИД 16122348 .
- ^ Приобретение геномов: теория происхождения видов. Маргулис Л. и Саган Д. (2002) Основные книги ISBN 0-465-04392-5
- ^ Эбби, Софи С.; Танньер, Эрик; Гуи, Маноло; Добен, Винсент (27 марта 2012 г.). «Боковой перенос генов как опора древа жизни» . Труды Национальной академии наук . 109 (13): 4962–4967. Бибкод : 2012PNAS..109.4962A . дои : 10.1073/pnas.1116871109 . ПМК 3323970 . ПМИД 22416123 .
- ^ Хуан, Цзиньлин; Гогартен, Иоганн Петер (2009). Горизонтальный перенос генов . Методы молекулярной биологии. Том. 532. Хумана Пресс. стр. 127–139. дои : 10.1007/978-1-60327-853-9_7 . ISBN 9781603278522 . ПМИД 19271182 .
- ^ Давин, Адриан А.; Танньер, Эрик; Уильямс, Том А.; Буссау, Бастьен; Добен, Винсент; Сёллёси, Гергеи Дж. (2 апреля 2018 г.). «Перенос генов позволяет датировать древо жизни» . Экология и эволюция природы . 2 (5): 904–909. дои : 10.1038/s41559-018-0525-3 . ISSN 2397-334X . ПМЦ 5912509 . ПМИД 29610471 .
- ^ Сёллёси, Гергеи Дж.; Танньер, Эрик; Лартильо, Николя; Добен, Винсент (01 мая 2013 г.). «Боковой перенос генов от мертвых» . Систематическая биология . 62 (3): 386–397. arXiv : 1211.4606 . дои : 10.1093/sysbio/syt003 . ISSN 1063-5157 . ПМЦ 3622898 . ПМИД 23355531 .
- В эту статью включены материалы из статьи Citizendium « Филогения и эволюция прокариот », которая распространяется по лицензии Creative Commons Attribution-ShareAlike 3.0 Unported License , но не по GFDL .
Дальнейшее чтение
[ редактировать ]- Гогартен, JP; Таунсенд, JP (2005). «Горизонтальный перенос генов, инновации и эволюция генома». Обзоры природы. Микробиология . 3 (9): 679–87. дои : 10.1038/nrmicro1204 . ПМИД 16138096 . S2CID 8092665 . . Одна статья в целом выпуске журнала Nature Reviews Microbiology в основном посвящена ГПГ.
- Добен, В; Моран, Северная Каролина; Охман, Х. (2003). «Филогенетика и сплоченность бактериальных геномов». Наука . 301 (5634): 829–32. Бибкод : 2003Sci...301..829D . дои : 10.1126/science.1086568 . ПМИД 12907801 . S2CID 11268678 .
- Эйзен, Дж. А.; Фрейзер, CM (2003). «Точка зрения филогеномики: пересечение эволюции и геномики». Наука . 300 (5626): 1706–7. Бибкод : 2003Sci...300.1706E . дои : 10.1126/science.1086292 . ПМИД 12805538 . S2CID 42394233 .
- Ге, Ф; Ван, Л.С.; Ким, Дж (2005). «Паутина жизни, выявленная с помощью оценок горизонтального переноса генов в масштабе генома» . ПЛОС Биология . 3 (10): е316. дои : 10.1371/journal.pbio.0030316 . ПМЦ 1233574 . ПМИД 16122348 .
- Хенц, СР; Хьюсон, Д.Х.; Кроме того, А.Ф.; Низельт-Штруве, К; Шустер, Южная Каролина (2005). «Полногеномная филогения прокариот» . Биоинформатика . 21 (10): 2329–35. doi : 10.1093/биоинформатика/bth324 . ПМИД 15166018 .
- Лерат, Э; Добен, В; Охман, Х; Моран, Н.А. (2005). «Эволюционное происхождение геномного репертуара бактерий» . ПЛОС Биология . 3 (5): е130. doi : 10.1371/journal.pbio.0030130 . ПМЦ 1073693 . ПМИД 15799709 .
- Урвин, Р; Мейден, MC (2003). «Мультилокусное типирование последовательностей: инструмент глобальной эпидемиологии». Тенденции Микробиол . 11 (10): 479–87. дои : 10.1016/j.tim.2003.08.006 . ПМИД 14557031 .
- Адутте, А; Балавуан, Г; Лартильо, Н.; Леспине, О; Прюдом, Б; Де Роза, Р. (2000). «Новая филогения животных: надежность и последствия» . Proc Natl Acad Sci США . 97 (9): 4453–6. Бибкод : 2000PNAS...97.4453A . дои : 10.1073/pnas.97.9.4453 . ПМК 34321 . ПМИД 10781043 .
- Дельсюк, Ф; Бринкманн, Х; Филипп, Х (2005). «и др. (2005) Филогеномика и реконструкция древа жизни». Нат преподобный Жене . 6 (5): 361–75. CiteSeerX 10.1.1.333.1615 . дои : 10.1038/nrg1603 . ПМИД 15861208 . S2CID 16379422 .
- Стенкамп, ET; Райт, Дж; Балдауф, СЛ (2006). «Протистанское происхождение животных и грибов» . Мол Биол Эвол . 23 (1): 93–106. дои : 10.1093/molbev/msj011 . ПМИД 16151185 .
- Томас, CM; Нильсен, КМ. (2005). «Механизмы и барьеры горизонтального переноса генов между бактериями». Nat Rev Микробиол . 3 (9): 711–21. дои : 10.1038/nrmicro1234 . ПМИД 16138099 . S2CID 1231127 .