Пилус
Пилюс лат ( . «волосы»; мн.: пили ) — это волосоподобный придаток, обнаруженный на поверхности многих бактерий и архей . [1] Термины пилус и фимбрия (лат. «бахрома»; множественное число: фимбрии ) могут использоваться как взаимозаменяемые, хотя некоторые исследователи оставляют термин пилус для придатка, необходимого для бактериальной конъюгации . Все конъюгативные пили состоят в основном из пилинов – волокнистых белков , которые являются олигомерными .
На поверхности бактерий и архей могут существовать десятки таких структур. Некоторые бактерии, вирусы или бактериофаги прикрепляются к рецепторам пилей в начале репродуктивного цикла.
Пили антигенны . Они также хрупкие и постоянно заменяются пилями различного состава, что приводит к изменению антигенности. Специфические реакции хозяина на старые структуры пилей не эффективны для новой структуры. Рекомбинация между генами некоторых (но не всех) пилей кодирует вариабельные (V) и константные (C) области пилей (аналогично разнообразию иммуноглобулинов ). В качестве первичных антигенных детерминант, факторов вирулентности и факторов безнаказанности на клеточной поверхности ряда видов грамотрицательных и некоторых грамположительных бактерий, включая Enterobacteriaceae , Pseudomonadaceae и Neisseriaceae , большой интерес вызвало изучение пилей как органеллы адгезию и в качестве компонентов вакцин. Первое детальное исследование пилей было проведено Бринтоном и его сотрудниками, которые продемонстрировали существование двух различных фаз внутри одного бактериального штамма: пилей (p+) и непилейных). [2]
Типы по функциям
[ редактировать ]Разным типам пилей дано несколько названий в зависимости от их функции. Классификация не всегда пересекается со структурными или эволюционными типами, поскольку происходит конвергентная эволюция . [3]
Конъюгативные пили
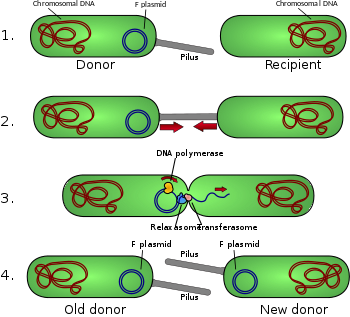
[ редактировать ]Конъюгативные пили позволяют передавать ДНК между бактериями в процессе бактериальной конъюгации . Их иногда называют «секс-пили» по аналогии с половым размножением , поскольку они позволяют обмениваться генами посредством образования «брачных пар». Пожалуй, наиболее хорошо изученным является F-пилус Escherichia coli , кодируемый F. половым фактором
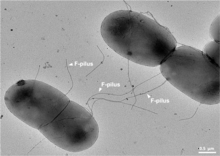
Половой пилус обычно от 6 до 7 нм имеет диаметр . Во время конъюгации пилус, выходящий из бактерии-донора, захватывает бактерию-реципиент, притягивает ее к себе и в конечном итоге запускает образование мостика спаривания , который устанавливает прямой контакт и образование контролируемой поры, позволяющей перенести ДНК от донора к получатель. Обычно передаваемая ДНК состоит из генов, необходимых для создания и переноса пилей (часто закодированных на плазмиде ), и поэтому является своего рода эгоистичной ДНК ; однако часто передаются и другие фрагменты ДНК, и это может привести к распространению генетических признаков по всей бактериальной популяции, таких как устойчивость к антибиотикам . Соединение, установленное F-pilus, чрезвычайно устойчиво механически и термохимически благодаря прочным свойствам F-pilus, что обеспечивает успешный перенос генов в различных средах. [5] Не все бактерии способны образовывать конъюгативные пили, но конъюгация может происходить между бактериями разных видов. [6] [7]
Гипертермофильные археи кодируют пили, структурно сходные с бактериальными конъюгативными пилями. [8] Однако, в отличие от бактерий, где аппарат конъюгации обычно опосредует перенос мобильных генетических элементов, таких как плазмиды или транспозоны, у гипертермофильных архей используется конъюгативный аппарат, называемый Ced (кренархейная система обмена ДНК). [9] и Тед (система Thermoproteales для обмена ДНК), [8] по-видимому, отвечает за передачу клеточной ДНК между представителями одного и того же вида. Было высказано предположение, что у этих архей полностью приручен механизм конъюгации, способствующий репарации ДНК посредством гомологичной рекомбинации, а не распространения мобильных генетических элементов. [8]
Фимбрии
[ редактировать ]
Фимбрия ( лат. «бахрома», мн.: fimbriae ) — это термин, обозначающий короткий пилус, который используется для прикрепления бактерии к поверхности, иногда также называемый «пилусом прикрепления». [10] или клейкий пилус . Термин «фимбрии» может относиться ко многим различным (структурным) типам пилуса. Действительно, для адгезии использовалось много различных типов пилей, что является примером конвергентной эволюции . [3] Система Gene Ontology не рассматривает фимбрии как отдельный тип придатков, вместо этого использует общий тип пилуса (GO:0009289).
Диаметр этого придатка составляет 3–10 нанометров, а длина может достигать нескольких микрометров. Фимбрии используются бактериями для прикрепления друг к другу, а также к клеткам животных и некоторым неодушевленным предметам. Бактерия может иметь до 1000 фимбрии. Фимбрии видны только при использовании электронного микроскопа . Они могут быть прямыми или гибкими.
Фимбрии обладают адгезинами , которые прикрепляют их к определенному субстрату, благодаря чему бактерии могут противостоять силам сдвига и получать питательные вещества. Например, E. coli использует их для прикрепления к маннозным рецепторам .
Некоторые аэробные бактерии образуют очень тонкий слой на поверхности бульонной культуры . Этот слой, называемый пелликулой , состоит из множества аэробных бактерий, которые прикрепляются к поверхности своими бахромками. Таким образом, фимбрии позволяют аэробным бактериям сохраняться как в бульоне, из которого они берут питательные вещества, так и вблизи воздуха.
Фимбрии необходимы для образования биопленки , поскольку они прикрепляют бактерии к поверхности хозяина для колонизации во время инфекции. Фимбрии либо располагаются у полюсов клетки, либо равномерно распределены по всей ее поверхности.
Этот термин также использовался в широком смысле для обозначения всех пилей теми, кто использует слово «пилус» конкретно для обозначения половых пилей. [11]
Типы по сборке системы или конструкции
[ редактировать ]В этом разделе отсутствует информация об упоминании других типов: различные шаперон-ушеровские фимбрии , построенные T7SS, внеклеточные пили нуклеации-преципитации, построенные T8SS (включая курли ), LPXTG, включая пилусы 3-го типа (T3P; spaHIG). [3] . ( декабрь 2020 г. ) |
Передача
[ редактировать ]Семейство Тра (переносное) включает все известные половые пили (по состоянию на 2010 год). Они связаны с системой секреции IV типа (T4SS). [3] Их можно разделить на F-подобный тип (после F-пилуса) и P-подобный тип. Как и их секреторные аналоги, пилусы вводят материал, в данном случае ДНК, в другую клетку. [12]
Пилы типа IV
[ редактировать ]
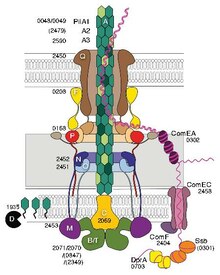
Некоторые пили, называемые пилями IV типа (T4P), генерируют подвижные силы. [14] Внешние концы пилей прикрепляются к твердой подложке — либо к поверхности, к которой прикреплена бактерия, либо к другим бактериям. Затем, когда пили сокращаются, они тянут бактерию вперед, как крюк. Движения, производимые пилями IV типа, обычно резкие, поэтому их называют подергивающейся подвижностью , в отличие от других форм бактериальной подвижности, таких как подвижность жгутиков . Однако некоторые бактерии, например Myxococcus xanthus , обладают скользящей подвижностью . Бактериальные пили IV типа по структуре сходны с компонентными белками архей (архейных жгутиков), и оба относятся к системе секреции типа II (T2SS); [15] они объединены группой нитевых систем типа IV . Помимо архей, многие археи производят адгезивные пили 4-го типа, которые позволяют клеткам архей прикрепляться к различным субстратам. N-концевые альфа-спиральные части пилинов и архееллинов архейного типа 4 гомологичны соответствующим областям бактериального T4P; однако С-концевые домены, богатые бета-цепями, по-видимому, не связаны между собой у бактериальных и архейных пилинов. [16]
Генетическая трансформация — это процесс, посредством которого бактериальная клетка-реципиент поглощает ДНК из соседней клетки и интегрирует эту ДНК в свой геном путем гомологичной рекомбинации . У Neisseria meningitidis (также называемого менингококком) трансформация ДНК требует присутствия коротких последовательностей поглощения ДНК (DUS), которые представляют собой 9-10 мономеров, находящихся в кодирующих областях донорской ДНК. Специфическое распознавание DUS опосредуется пилином IV типа . [17] Пили меннингококка IV типа связываются с ДНК через минорный пилин ComP через электроположительную полосу, которая, как ожидается, будет обнажена на поверхности нити. ComP демонстрирует изысканные предпочтения привязки для выборочных DUS. Распределение DUS в геноме N. meningitides благоприятствует определенным генам, что позволяет предположить, что существует предвзятость в отношении генов, участвующих в поддержании и восстановлении генома. [18] [19]
Первоначально это семейство было идентифицировано как «бахромки IV типа» по их внешнему виду под микроскопом. Эта классификация сохранилась, поскольку соответствует кладе. [20] Было показано, что некоторые пилины архейного типа IV могут существовать в 4 различных конформациях, образуя два пилина с совершенно разной структурой. [21] Примечательно, что оба пили были произведены одним и тем же механизмом секреции. Однако то, какой из двух пилей образуется, по-видимому, зависит от условий роста, что позволяет предположить, что два пили функционально различны. [21]
Фимбрии 1 типа
[ редактировать ]Другой тип называется фимбриями 1 типа. [22] На «кончиках» они содержат адгезины FimH. Путь шаперон-ушер отвечает за перемещение многих типов фимбрий из клетки, включая фимбрии 1 типа. [23] и P бахромки . [24]
Кудрявый
[ редактировать ]
«Грам-отрицательные бактерии собирают функциональные амилоидные поверхностные волокна, называемые курли ». [26] Курли представляют собой разновидность фимбрий. [22] Керли состоят из белков, называемых керлинами. [26] Некоторые из задействованных генов: CsgA , CsgB , CsgC , CsgD , CsgE , CsgF и CsgG . [26]
Вирулентность
[ редактировать ]Пили ответственны за вирулентность патогенных штаммов многих бактерий, включая кишечную палочку , холерный вибрион и многие штаммы стрептококков . [27] [28] Это связано с тем, что наличие пилей значительно увеличивает способность бактерий связываться с тканями организма, что затем увеличивает скорость репликации и способность взаимодействовать с организмом-хозяином. [27] Если у вида бактерий есть несколько штаммов, но только некоторые из них являются патогенными, вполне вероятно, что патогенные штаммы будут иметь пили, а непатогенные штаммы - нет. [29] [30]
Развитие прикрепленных пилей может затем привести к развитию дальнейших признаков вирулентности. Фимбрии являются одним из основных механизмов вирулентности бактерий E. coli , Bordetella pertussis , стафилококков и стрептококков . Их присутствие значительно увеличивает способность бактерий прикрепляться к хозяину и вызывать заболевание. [31] Непатогенные штаммы V. cholerae сначала развили пили, что позволило им связываться с тканями человека и образовывать микроколонии . [27] [30] Эти пили затем служили сайтами связывания для лизогенного бактериофага , несущего болезнетворный токсин . [27] [30] Ген этого токсина, однажды включенный в геном бактерии, экспрессируется, когда экспрессируется ген, кодирующий пилус (отсюда и название «пилус, опосредованный токсином»). [27]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «пилус» в Медицинском словаре Дорланда.
- ^ Бринтон, Чарльз (1954). «Исследование электрофореза и чувствительности к фагам варианта бактерии E. coli, продуцирующего нити». Биохимика и биофизика Acta . 15 (4): 533–542. дои : 10.1016/0006-3002(54)90011-6 . ПМИД 13230101 .
- ^ Перейти обратно: а б с д Шагно, К; Зоргани, Массачусетс; Астрюк, Т; Дево, М. (14 октября 2013 г.). «Белковые детерминанты поверхностной колонизации бактерий: бактериальная адгезия и образование биопленок с точки зрения секреции белка» . Границы микробиологии . 4 : 303. дои : 10.3389/fmicb.2013.00303 . ПМК 3796261 . ПМИД 24133488 .
- ^ «Кишечные бактерии используют суперполимеры, чтобы уклониться от антибиотиков | Imperial News | Имперский колледж Лондона» . Имперские новости .
- ^ Патковски, Йонаш Б.; Дальберг, Тобиас; Амин, Химани; Галот, Дхармендер К.; Виджайраджратнам, Сухитасри; Фогель, Джозеф П.; Фрэнсис, Мэтью С.; Бейкер, Джозеф Л.; Андерссон, Магнус; Коста, Тьяго РД (5 апреля 2023 г.). «Биомеханическая адаптируемость F-pilus ускоряет конъюгативное распространение резистентности к противомикробным препаратам и образование биопленок» . Природные коммуникации . 14 (1): 1879. doi : 10.1038/s41467-023-37600-y . ПМЦ 10076315 . ПМИД 37019921 .
- ^ Петижан, Мари; Кондамин, Бенедикт; Бурде, Чарльз; Денамур, Эрик; Руппе, Этьен (2021). «Филумный барьер и внутривидовая филогения Escherichia coli способствуют приобретению генов устойчивости к антибиотикам» . Микробная геномика . 7 (8): 000489. doi : 10.1099/mgen.0.000489 . ISSN 2057-5858 . ПМЦ 8549366 . ПМИД 34435947 .
- ^ Гшвинд, Реми; Петижан, Мари; Фурнье, Клодин; Лао, Джули; Клермон, Оливье; Нордманн, Патрис; Мельманн, Александр; Денамур, Эрик; Пуарель, Лоран; Руппе, Этьен (3 апреля 2024 г.). Улеманн, Анн-Катрин (ред.). «Межтиповая циркуляция гена, кодирующего бета-лактамазу: редкое, но наблюдаемое событие» . Антимикробные средства и химиотерапия . 68 (4): e0145923. дои : 10.1128/aac.01459-23 . ISSN 0066-4804 . ПМЦ 10989005 . ПМИД 38441061 .
- ^ Перейти обратно: а б с д Бельтран, Летисия К.; Цвиркайте-Крупович, Вирджиния; Миллер, Джессалин; Ван, Фэнбинь; Крейцбергер, Марк А.Б.; Патковски, Йонаш Б.; Коста, Тьяго Р.Д.; Схоутен, Стефан; Левенталь, Илья; Контичелло, Винсент П.; Эгельман, Эдвард Х.; Крупович, Март (07 февраля 2023 г.). «Аппарат для импорта ДНК архей гомологичен бактериальному аппарату конъюгации» . Природные коммуникации . 14 (1): 666. Бибкод : 2023NatCo..14..666B . дои : 10.1038/s41467-023-36349-8 . ISSN 2041-1723 . ПМЦ 9905601 . ПМИД 36750723 .
- ^ ван Вулферен, Марлин; Вагнер, Александр; ван дер Дос, Крис; Альберс, Соня-Верена (01 марта 2016 г.). «Архейная система Ced импортирует ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (9): 2496–2501. Бибкод : 2016PNAS..113.2496V . дои : 10.1073/pnas.1513740113 . ISSN 1091-6490 . ПМЦ 4780597 . ПМИД 26884154 .
- ^ Профт, Т.; Бейкер, EN (февраль 2009 г.). «Пили у грамотрицательных и грамположительных бактерий — строение, сборка и их роль в заболевании» . Клеточные и молекулярные науки о жизни . 66 (4): 613–635. дои : 10.1007/s00018-008-8477-4 . ПМЦ 11131518 . ПМИД 18953686 . S2CID 860681 .
- ^ Оттоу, Дж. К. (1975). «Экология, физиология и генетика бахромок и пилей». Ежегодный обзор микробиологии . 29 : 79–108. дои : 10.1146/annurev.mi.29.100175.000455 . ПМИД 1180526 .
- ^ Филлу, А. (июль 2010 г.). «Разновидность бактериальных пилей, участвующих в горизонтальном переносе генов» . Журнал бактериологии . 192 (13): 3243–5. дои : 10.1128/JB.00424-10 . ПМЦ 2897649 . ПМИД 20418394 .
- ^ Джоан, Слончевски (2017). Микробиология: развивающаяся наука . Фостер, Джон Уоткинс (Четвертое изд.). Нью-Йорк: WW Norton & Company. стр. 1000–1002. ISBN 9780393614039 . OCLC 951925510 .
- ^ Мэттик Дж.С. (2002). «Пили IV типа и подергивания моторики». Анну. Преподобный Микробиол . 56 (1): 289–314. дои : 10.1146/annurev.micro.56.012302.160938 . ПМИД 12142488 .
- ^ Джаррелл; и др. (2009). «Археальные жгутики и пили». Пили и жгутики: текущие исследования и будущие тенденции . Кайстер Академик Пресс. ISBN 978-1-904455-48-6 .
- ^ Ван, Ф; Цвиркайте-Крупович, В; Крейцбергер, МАБ; Су, З; де Оливейра, GAP; Осинский, Т; Шерман, Н; ДиМайо, Ф; Уолл, Дж. С.; Прангишвили, Д; Крупович, М; Эгельман, Э.Х. (2019). «Обильно гликозилированные архейные пилюсы выживают в экстремальных условиях» . Природная микробиология . 4 (8): 1401–1410. дои : 10.1038/s41564-019-0458-x . ПМК 6656605 . ПМИД 31110358 .
- ^ Чеховин А., Симпсон П.Дж., Макдауэлл М.А., Браун Д.Р., Ношезе Р., Паллетт М., Брэйди Дж., Болдуин Г.С., Леа С.М., Мэтьюз С.Дж., Пеличич В. (2013). «Специфическое распознавание ДНК, опосредованное пилином IV типа» . Учеб. Натл. акад. наук. США . 110 (8): 3065–70. Бибкод : 2013PNAS..110.3065C . дои : 10.1073/pnas.1218832110 . ПМК 3581936 . ПМИД 23386723 .
- ^ Дэвидсен Т., Родланд Э.А., Лагесен К., Сиберг Э., Рогнес Т., Тоньюм Т. (2004). «Смещенное распределение последовательностей поглощения ДНК в сторону генов поддержания генома» . Нуклеиновые кислоты Рез . 32 (3): 1050–8. дои : 10.1093/nar/gkh255 . ПМЦ 373393 . ПМИД 14960717 .
- ^ Каугант Д.А., Maiden MC (2009). «Менингококковое носительство и болезни – популяционная биология и эволюция» . Вакцина . 27 Приложение 2 (4): B64–70. doi : 10.1016/j.vaccine.2009.04.061 . ПМЦ 2719693 . ПМИД 19464092 .
- ^ Нуччио С.П. и др. (2007). «Эволюция пути сборки шаперона / помощника: фимбриальная классификация идет по-гречески» . Обзоры микробиологии и молекулярной биологии . 71 (4): 551–575. дои : 10.1128/MMBR.00014-07 . ПМК 2168650 . ПМИД 18063717 .
- ^ Перейти обратно: а б Лю, Дж; Истеп, GN; Цвиркайте-Крупович, В; Рич-Нью, Северная Каролина; Крейцбергер, МАБ; Эгельман, Э.Х.; Крупович, М; Ван, Ф (2024). «Две различные структуры пилей архейного типа IV, образованные белками с идентичной последовательностью» . Природные коммуникации . 15 (1): 5049. doi : 10.1038/s41467-024-45062-z . ПМЦ 11178852 . ПМИД 38877064 .
- ^ Перейти обратно: а б Куксон, Алабама; Кули, Вашингтон; Вудворд, MJ (2002), «Роль типа 1 и завитковых бахромок Escherichia coli, продуцирующих шига-токсин, в прилипании к абиотическим поверхностям», Int J Med Microbiol , 292 (3–4): 195–205, doi : 10.1078/ 1438-4221-00203 , ПМИД 12398210 .
- ^ Коленда, Рафаль; Угорский, Мацей; Гжимайло, Кшиштоф (14 мая 2019 г.). «Все, что вы всегда хотели знать о фимбриях сальмонеллы 1 типа, но боялись спросить» . Границы микробиологии . 10 :1017. дои : 10.3389/fmicb.2019.01017 . ПМК 6527747 . ПМИД 31139165 .
- ^ Райс Дж.К., Пэн Т., Спенс Дж.С., штаб-квартира Вана, Голдблюм Р.М., Кортези Б., Новицки Б.Дж. (декабрь 2005 г.). «Пиелонефритическая Escherichia coli, экспрессирующая P-фимбрии, снижает иммунный ответ почек мыши» . Журнал Американского общества нефрологов . 16 (12): 3583–91. дои : 10.1681/ASN.2005030243 . ПМИД 16236807 .
- ^ Висконсин, Кеннет Тодар, Мэдисон. «Колонизация и инвазия бактериальных патогенов» . www.textbookofbacteriology.net . Проверено 3 декабря 2016 г.
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б с Эпштейн, Э.А.; Рейзиан, Массачусетс; Чепмен, М.Р. (2009), «Пространственная кластеризация липопротеина секреции курлина требует сборки волокон курли». J Bacteriol , 191 (2): 608–615, doi : 10.1128/JB.01244-08 , PMC 2620823 , PMID 19011034 .
- ^ Перейти обратно: а б с д и Крейг, Лиза; Тейлор, Рональд (2014). «Глава 1: Пилус, регулируемый токсином холерного вибриона: структура, сборка и функции, имеющие значение для разработки вакцин». В Барокки, Мишель; Телфорд, Джон (ред.). Бактериальные пили: структура, синтез и роль в заболевании . КАБ Интернешнл. стр. 1–16. ISBN 978-1-78064-255-0 .
- ^ Ринаудо, Даниэла; Москиони, Моника (2014). «Глава 13: Разработка вакцины против стрептококков на основе пилуса: изменчивость, разнообразие и иммунологический ответ». В Барокки, Мишель; Телфорд, Джон (ред.). Бактериальные пили: структура, синтез и роль в заболевании . КАБ Интернешнл. стр. 182–202. ISBN 978-1-78064-255-0 .
- ^ Тодар, Кеннет. «Учебник бактериологии: Структура бактерий в связи с патогенностью» . Учебник бактериологии . Проверено 24 ноября 2017 г.
- ^ Перейти обратно: а б с Джорджиаду, Микаэлла; Пеличич, Владимир (2014). «Глава 5: Пили типа IV: функции и биогенез». В Барокки, Мишель; Телфорд, Джон (ред.). Бактериальные пили: структура, синтез и роль в заболевании . КАБ Интернешнл. стр. 71–84. ISBN 978-1-78064-255-0 .
- ^ Коннелл И., Агаче В., Клемм П., Шембри М., Мэрилд С., Сванборг К. (сентябрь 1996 г.). «Экспрессия фимбрионов типа 1 повышает вирулентность Escherichia coli для мочевыводящих путей» . Учеб. Натл. акад. наук. США . 93 (18): 9827–32. Бибкод : 1996PNAS...93.9827C . дои : 10.1073/pnas.93.18.9827 . ПМК 38514 . ПМИД 8790416 .
Внешние ссылки
[ редактировать ]- Sex + Pilus в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Bacterial+Pilus в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Фимбрии + белки Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
| Медицинский микробиология | |||||||
|---|---|---|---|---|---|---|---|
| Биохимия и экология |
| ||||||
| Форма | |||||||
| Структура |
| ||||||
| Таксономия и эволюция | |||||||