Шапероны-проводники бахромки
Шаперон-ушерные фимбрии (CU) представляют собой линейные неразветвленные на внешней мембране пили , секретируемые грамотрицательными бактериями через систему шаперон-ушер, а не через секрецию типа IV или системы внеклеточной нуклеации. Эти фимбрии построены из модульных субъединиц ворсинок, которые транспортируются в периплазму Sec- зависимым образом. Фимбрии, секретируемые шаперонами-ушерами, являются важными патогенности, факторами способствующими колонизации хозяина , локализации и образованию биопленок у клинически важных видов, таких как уропатогенные Escherichia coli и Pseudomonas aeruginosa .
Структура
[ редактировать ]Общий
[ редактировать ]Все системы шаперон/ушер находятся в кластерах генов, состоящих по крайней мере из шаперона , шаперона и одной или нескольких субъединиц фимбрий. [ 1 ] В целом система включает периплазматические шапероны, периплазматические и внеклеточные субъединицы ворсинок , димерную пору наружной мембраны и связанный с ней аппарат Sec. Субъединицы пилуса полимеризуются посредством нековалентного взаимодействия с образованием зрелого пилуса, который состоит из адгезивного кончика, спирального тела и связанного с выступом основания. Система P-pilus (pap) является одной из наиболее изученных и показана ниже. [ 2 ]
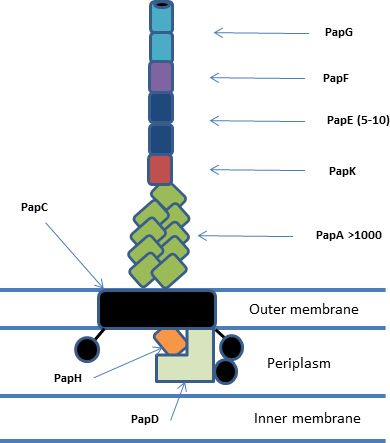
Структура и функции Ашера
[ редактировать ]Ашер образует пору внешней мембраны и функционирует in vivo как димер , хотя в любой момент времени активен только один из ашеров. [ 3 ] Входная пора (PapC) образована 24-нитевым бета-цилиндром с 4 дополнительными доменами: N-концевым доменом (NTD), Plug-доменом и двумя C-концевыми доменами (CTD1 и CTD2 соответственно). Функция NTD и CTDs заключается в доставке комплексов шаперон/субъединица к растущему ворсу и облегчении транслокации через пору. [ 4 ] Домен пробки предотвращает преждевременное образование ворсинок и может располагаться либо внутри поры, либо на периплазматической стороне мембраны. В активном димерном ашере у одного из ашеров пробка расположена периплазматически, а у другого - внутри транслокационной поры. [ 5 ]
Структура и функции шаперона
[ редактировать ]Периплазматический шаперон (PapD) имеет структуру «бумеранга», образованную складкой, подобной иммуноглобулину (Ig), с существенным удлинением С-конца (G1). Эта складка образована 13 β-тяжами (A1-G1) и 4 короткими α-спиралями. Шапероны принадлежат к одному из двух семейств в зависимости от длины петли, соединяющей бета-цепи F1 и G1. Шапероны с длинной петлей — это FGL, а шапероны с короткой петлей — FGS. Функция шаперона останавливает агрегацию и деградацию субъединиц ворсинок, связывая их сопрягающие области и способствуя правильному сворачиванию белков перед их переносом в комплекс ушеров/фимбрий. [ 6 ]
Структура и функции субъединицы пилина
[ редактировать ]
Структура субъединицы пилуса также имеет Ig-подобную складку, но с существенным N-концевым расширением, а не C-концевым расширением. Поскольку это N-концевое удлинение представляет собой ту же β-цепь, которая в норме завершает складку Ig, в зрелой субъединице образуется большая гидрофобная щель (P1-P5). Эта щель не только важна для биогенеза пилуса, но также требует шаперона для стабилизации структура. [ 7 ] Отдельные субъединицы могут иметь дополнительные домены, например, papG имеет адгезивный лектиновый домен. In vivo субъединицы ворсинок формируют основную часть спирального ворса (приблизительно 3,28 субъединицы на виток) и адгезивную головку.
Биогенез
[ редактировать ]Биогенез ворсинок протекает по механизму комплементации цепей, причем этот механизм функционирует как прямое следствие структуры как шаперонов, так и субъединиц ворсинок. Как G1-удлинение шаперона, так и N-концевое удлинение субъединиц ворсинок могут связываться с гидрофобной щелью P1-P5. Когда G1 связан, он связывается в параллельной конформации, стабилизируя структуру субъединицы, но не образует настоящую складку Ig (которая является антипараллельной). Он также связывает расщелину только на участках P1-P4, оставляя открытым карман P5. Когда другая субъединица связывается с гидрофобной щелью, она связывается антипараллельно P5-P2, создавая наиболее энергетически выгодную конформацию истинной Ig-складки. [ 1 ]
Когда комплекс шаперон/субъединица переносится к растущему комплексу шаперон/субъединица (обратите внимание, что комплекс шаперон/субъединица образует основу этого), NTD и CTD димера помощника переносят и удерживают комплекс шаперон/субъединица на основании пилуса. Здесь N-концевой отросток субъединицы взаимодействует со свободным карманом P5 пилуса. Это связывание вызывает «расстегивание» остатка за остатком цепи G1 шаперона с одновременным «застегиванием» N-концевого удлинения комплекса шаперон/субъединица, что известно как механизм «zip-in-zip-out». [ 2 ]
Как только шаперон вытесняется, он диффундирует обратно в механизм sec, чтобы связать другую возникающую субъединицу пилуса. Затем ашер, использующий CTD, перемещает более длинный пилус через пору транслокации на один остаток до тех пор, пока шаперон не окажется в тесной связи с ашером. Эта транслокация сопровождается небольшим вращением, придающим пилусу спиральную структуру. Этот рост продолжается до тех пор, пока терминирующая субъединица не свяжет пилус (если он существует), терминирующие субъединицы лишены кармана P5 и, таким образом, почти необратимо связывают свой шаперон. Поскольку ашер может перемещать только субъединицы пилуса, но не комплексы шаперон/субъединица, а также тот факт, что шаперон не может быть энергетически превзойден другой субъединицей (из-за отсутствия P5), эта терминирующая субъединица закрепляет пилус на внешней мембране и остановить дальнейший рост пилуса. [ 1 ]
Номенклатура и классификация
[ редактировать ]Исторически классификация фимбрий проводилась на основе их внешнего вида под микроскопом, в результате чего были выделены 4 класса: афимбриальные, фимбрии 1-го типа, пучковообразующие (тип IV) ворсинки и завитки . Однако этот тип классификации не дает никакой меры родства и поэтому был заменен филогенетической системой. [ 8 ]
В фимбриях шаперона / помощника в целом существует два типа классификации: на основе типа присутствующего шаперона или на основе типа присутствующего шаперона. Использование ашера в качестве основы классификации формирует 5 клад: альфа, бета, гамма, пи и сигма, а также четыре субклада: гамма 1–4. Названия бета- и гамма-клад просто в алфавитном порядке, тогда как остальные названы по определяющему признаку: альфа для альтернативного семейства шаперонов/помощников; pi для P-пилуса (связанного с пиелонефритом); и сигма для пилуса споровой оболочки Myxococcus xanthus . [ 8 ]
Как указывалось ранее, существует два типа сопровождающих; FGL и FGS, и это основа классификации шаперонов. Все шапероны FGL связаны с афимбриальной секрецией, а все шапероны FGS связаны с фимбриальной секрецией. Все шапероны FGL обнаружены в субкладе гамма-3, все остальные клады и субклады представляют собой FGS. [ 8 ]
Патогенность
[ редактировать ]Шапероны/проводящие фимбрии играют множество ролей на многих стадиях патогенности у нескольких видов. Наиболее хорошо изучены роли в адгезии, уклонении от врожденного иммунитета и локализации. Адгезии способствует лектиновый домен на кончике шаперонных/ушерных фимбрий, этот домен FimH хорошо изучен. FimH является первой субъединицей пилуса типа I (клада γ-1) у таких видов, как UPEC, и связывается с D-маннозилированными остатками в мочевом пузыре, обеспечивая адгезию и колонизацию патогена. Ускользанию от иммунитета также способствует кончиковый адгезин, например, связывание фактора ускорения распада (DAF) фимбриями клады гамма3 ингибирует эффективную активацию комплемента. Локализация может осуществляться путем последовательной экспрессии различных систем шаперонов/помощников. Большинство видов патогенных бактерий экспрессируют более одного типа системы шаперон/проводник, например, у Pseudomonas aeruginosa имеется пять различных систем. UPEC экспрессирует как фимбрии типа 1, так и P-пилусы (pap), которые он экспрессирует последовательно, возможно, для облегчения миграции из мочевого пузыря (фимбрии I типа) в почки (pap). [ 9 ]
Такая важность патогенности делает систему шаперон/проводник привлекательной мишенью для новых лекарственных методов лечения. Путем воздействия на патогенность, а не на организм (в случае обычных антибиотиков) устраняется сильное селективное давление, способствующее развитию резистентности. В случае с УПЭК были достигнуты определенные успехи в прекращении образования ворсинок в клинических испытаниях и разработке эффективной вакцины на мышах и яванских макак на основе комплекса FimCH. [ 9 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Буш А. и Ваксман Г (2012). «Пути шаперона-проводника: разнообразие и механизм сборки пилуса» . Философские труды Лондонского королевского общества Б. 367 (1592): 1112–1122. дои : 10.1098/rstb.2011.0206 . ПМЦ 3297437 . ПМИД 22411982 .
- ^ Перейти обратно: а б Хультгрен С.Дж. и Ваксман Г. (2009). «Структурная биология шаперон-проводника биогенеза пилуса» . Обзоры природы Микробиология . 7 (11): 765–774. дои : 10.1038/nrmicro2220 . ПМК 3790644 . ПМИД 19820722 .
- ^ Вернебург, Гленн Т.; Хендерсон, Надин С.; Портной, Эрика Б.; Саровар, Самема; Халтгрен, Скотт Дж.; Ли, Хуэйлинь; Танасси, Дэвид Г. (июль 2015 г.). «Пилус-ушер контролирует взаимодействие белков посредством маскировки доменов и функционирует как олигомер» . Структурная и молекулярная биология природы . 22 (7): 540–546. дои : 10.1038/nsmb.3044 . ISSN 1545-9985 . ПМЦ 4496297 . ПМИД 26052892 .
- ^ Ветш М. и др. (2006). «Механизм сборки волокон через путь шаперон-проводник» . Отчеты ЭМБО . 7 (7): 734–738. дои : 10.1038/sj.embor.7400722 . ПМК 1500831 . ПМИД 16767077 .
- ^ Фан Дж. и др. (2011). «Кристаллическая структура ашера FimD, связанного с родственным ему субстратом FimC-FimH» . Природа . 474 (7349): 49–53. дои : 10.1038/nature10109 . ПМК 3162478 . ПМИД 21637253 .
- ^ Зауэр Ф. и др. (1999). «Структурные основы функции шаперона и биогенеза пилуса». Наука . 285 (5430): 1058–1061. дои : 10.1126/science.285.5430.1058 . ПМИД 10446050 .
- ^ Вергер Д. и др. (2007). «Кристаллическая структура субъединицы PapA P-pilus rob» . ПЛОС ОДИН . 3 (5): е73. дои : 10.1371/journal.ppat.0030073 . ПМК 1868955 . ПМИД 17511517 .
- ^ Перейти обратно: а б с Нуччио С.П. и др. (2007). «Эволюция пути сборки шаперона / помощника: фимбриальная классификация идет по-гречески» . Обзоры микробиологии и молекулярной биологии . 71 (4): 551–575. дои : 10.1128/MMBR.00014-07 . ПМК 2168650 . ПМИД 18063717 .
- ^ Перейти обратно: а б Беата М. и др. (2011). «Инфекции мочевыводящих путей штаммов Escherichia coli шаперонно-проводниковой системы» . Польский журнал микробиологии . 60 (4): 279–285. дои : 10.33073/pjm-2011-040 . ПМИД 22390061 .