Радиолярии
| Радиолярии Временной диапазон: | |
|---|---|
 | |
| Иллюстрация радиолярий из Челленджера экспедиции 1873–1876 гг. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | потогонные средства |
| Клэйд : | САР |
| Тип: | Ретария |
| Подтип: | Радиолярии Кавальер-Смит , 1987 год. |
| Классы | |
| |
| Часть серии о |
| Планктон |
|---|
 |
Радиолярии Radiozoa , также называемые , обычно , представляют собой простейшие диаметром 0,1–0,2 мм, образующие сложные минеральные скелеты с центральной капсулой, разделяющей клетку на внутреннюю и внешнюю части эндоплазмы и эктоплазмы . Сложный минеральный скелет обычно состоит из кремнезема . [1] Они встречаются в виде зоопланктона по всему Мировому океану. Как и зоопланктон, радиолярии преимущественно гетеротрофны , но многие из них имеют фотосинтезирующие эндосимбионты и поэтому считаются миксотрофами . Скелетные остатки некоторых видов радиолярий составляют большую часть покрова дна океана в виде кремнистого ила . Из-за быстрого изменения видов и сложного скелета радиолярии представляют собой важную диагностическую окаменелость, обнаруженную начиная с кембрия .
Описание
[ редактировать ]Радиолярии имеют множество игольчатых ложноножек, поддерживаемых пучками микротрубочек , которые обеспечивают плавучесть радиолярий. Ядро клетки и большинство других органелл находятся в эндоплазме, тогда как эктоплазма заполнена пенистыми вакуолями и каплями липидов , поддерживающими их плавучесть. Радиолярии часто могут содержать симбиотические водоросли, особенно зооксантеллы , которые обеспечивают большую часть энергии клетки. Частично эта организация встречается у солнечников , но у них отсутствуют центральные капсулы, и они образуют только простые чешуйки и шипы.
Некоторые радиолярии известны своим сходством с правильными многогранниками , например, икосаэдра, в форме икосаэдры Circogonia изображенные ниже.
Таксономия
[ редактировать ]Радиолярии относятся к надгруппе Rhizaria вместе с ( амебоидными или жгутиковыми ) Cercozoa и (раковинными амебоидными) Foraminifera . [2] Традиционно радиолярии делят на четыре группы — Acantharea , Nassellaria , Spumellaria и Phaeodarea . Однако сейчас феодарию считают церкозоем. [3] [4] Nassellaria и Spumellaria образуют кремнистый скелет и поэтому были сгруппированы в группу Polycystina . Несмотря на некоторые первоначальные предположения об обратном, это также подтверждается молекулярной филогенией. Acantharea производит скелеты из сульфата стронция и тесно связана с своеобразным родом Sticholonche ( Taxopodida ), у которого отсутствует внутренний скелет и который долгое время считался солнечником . Таким образом, радиолярии можно разделить на две основные линии: Polycystina (Spumellaria + Nassellaria) и Spasmaria (Acantharia + Taxopodida). [5] [6]
Существует несколько групп высшего порядка, которые были обнаружены при молекулярном анализе данных окружающей среды. В частности, группы, связанные с Акантарией. [7] и Спумеллария. [8] Эти группы пока совершенно неизвестны с точки зрения морфологии и физиологии, и поэтому разнообразие радиолярий, вероятно, будет намного выше, чем известно в настоящее время.
Отношения между фораминиферами и радиоляриями также обсуждаются. Молекулярные деревья поддерживают их тесную связь — группу, названную Ретарией. [9] Но являются ли они сестринскими линиями или следует включить Foraminifera в состав Radiolaria, неизвестно.
| Сорт | Заказ | Изображение | Семьи | Роды | Разновидность | Описание |
|---|---|---|---|---|---|---|
| Полицистея | Насселлярия |  | ... | |||
| Спумеллария | 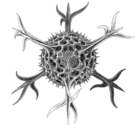 | ... | ||||
| Коллодария |  | ... | ||||
| Акантарея |  | ... | ||||
| Стихолонхея | У таксопода |  | 1 | 1 | 1 | ... |
Биогеография
[ редактировать ]
На диаграмме справа а показаны генерализованные провинции радиолярий. [10] [11] и их связь с температурой водных масс (оттенки теплых и холодных цветов) и циркуляцией (серые стрелки). Из-за погружения водной массы в высоких широтах под теплые стратифицированные воды в более низких широтах виды радиолярий занимают места обитания на разных широтах и глубинах мирового океана. Таким образом, морские отложения из тропиков представляют собой совокупность нескольких вертикально расположенных комплексов фауны, некоторые из которых соприкасаются с поверхностными комплексами более высоких широт. Отложения под полярными водами включают космополитные глубоководные радиолярии, а также эндемичные для высокоширотных поверхностных вод виды. Звездочки в ( а ) обозначают широты, в которых были отобраны пробы, а серые столбцы обозначают комплексы радиолярий, включенные в каждый осадочный композит. Горизонтальные фиолетовые полосы обозначают широты, известные хорошей сохранностью радиолярий (кремнезема), судя по составу поверхностных отложений. [12] [13]
Данные показывают, что некоторые виды были истреблены из высоких широт, но сохранились в тропиках в позднем неогене либо в результате миграции, либо в результате ограничения ареала ( б ). Учитывая прогнозируемое глобальное потепление, современные виды Южного океана не смогут использовать миграцию или сокращение ареала, чтобы избежать стрессовых факторов окружающей среды, поскольку их предпочтительные холодноводные места обитания исчезают с земного шара ( c ). Однако тропические эндемичные виды могут расширять свой ареал в сторону средних широт. Цветные полигоны на всех трех панелях представляют собой обобщенные биогеографические провинции радиолярий, а также их относительные температуры водных масс (более холодные цвета указывают на более низкие температуры, и наоборот). [13]
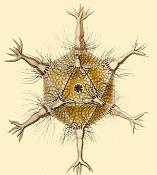 Циркогонии икосаэдры , виды радиолярий, имеющие форму правильного икосаэдра.
Циркогонии икосаэдры , виды радиолярий, имеющие форму правильного икосаэдра. Anthocyrtium hispidum Геккеля
Anthocyrtium hispidum Геккеля


Раковины радиолярий
[ редактировать ]

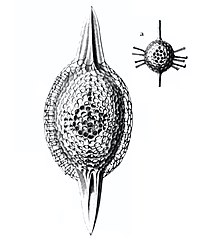
Радиолярии — это одноклеточные хищные протисты , заключенные в сложные шаровидные оболочки, обычно сделанные из кремнезема и пронизанные отверстиями. Их название происходит от латинского слова «радиус». Они ловят добычу, вытягивая части своего тела через отверстия. Как и в случае с кремнеземными панцирями диатомей, раковины радиолярий могут опускаться на дно океана, когда радиолярии умирают, и сохраняются как часть океанских отложений. Эти останки в виде микрокаменелостей предоставляют ценную информацию о прошлых условиях океана. [14]
 Как и диатомеи, радиолярии бывают разных форм.
Как и диатомеи, радиолярии бывают разных форм. Раковины радиолярий, как и диатомеи, обычно состоят из силиката.
Раковины радиолярий, как и диатомеи, обычно состоят из силиката. Однако акантарианские радиолярии имеют раковины из сульфата стронция. кристаллов
Однако акантарианские радиолярии имеют раковины из сульфата стронция. кристаллов




 Схематическая диаграмма сферической раковины радиолярий в разрезе
Схематическая диаграмма сферической раковины радиолярий в разрезе Кладококк абиетинус
Кладококк абиетинус


Поэтому я приступил к поиску решения уравнений морфогенеза на сфере. Теория заключалась в том, что сферический организм подвергался диффузии через свою поверхностную мембрану инородного вещества, например морской воды. Уравнения были:
Функция Утверждалось, что , принятый за радиус-вектор от центра до любой точки на поверхности мембраны, можно представить в виде ряда нормализованных функций Лежандра . Алгебраическое решение приведенных выше уравнений заняло в моей диссертации около 30 страниц и поэтому здесь не воспроизводится. Полностью они описаны в книге под названием «Морфогенез», посвященной Тьюрингу, под редакцией П.Т. Сондерса, опубликованной издательством North Holland в 1992 году. [16]
Алгебраическое решение уравнений выявило семейство решений, соответствующее параметру n, принимающему значения 2, 4, 6.
Решив алгебраические уравнения, я воспользовался компьютером, чтобы построить форму получившихся организмов. Тьюринг сказал мне, что существуют реальные организмы, соответствующие тем, что я создал. Он рассказал, что они были описаны и изображены в записях о путешествиях HMS Challenger в XIX веке.
Я решил уравнения и получил ряд решений, которые соответствовали реальным видам радиолярий, обнаруженным на корабле HMS Challenger в 19 веке. Эта экспедиция в Тихий океан обнаружила восемь вариантов моделей роста. Они показаны на следующих рисунках. Существенной особенностью роста является появление удлиненных «шипов», торчащих из сферы в правильных положениях. Таким образом, вид включал две, шесть, двенадцать и двадцать разновидностей шипов.



Бернард Ричардс , 2006 г. [17]
Разнообразие и морфогенез
[ редактировать ]Бернард Ричардс под руководством Алана Тьюринга (1912–1954) в Манчестере в качестве одного из последних учеников Тьюринга, помогая подтвердить Тьюринга теорию морфогенеза работал . [18] [19] [20] [21]
«Тьюринг стремился продолжить работу, которую Д'Арси Томпсон опубликовал в книге «О росте и форме» в 1917 году». [20]

точно повторяют некоторые образцы раковин радиолярий [22]
| Внешние видео | |
|---|---|
- Вариации позвоночника радиолярий, обнаруженные кораблем HMS Challenger в XIX веке и нарисованные Эрнстом Геккелем.
 Cromyactus тетрацелифус с 2 шипами.
Cromyactus тетрацелифус с 2 шипами.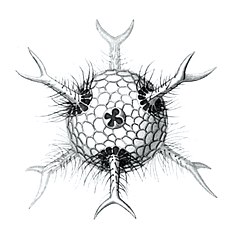 Circopus sexfurcus с 6 шипами.
Circopus sexfurcus с 6 шипами. Circopurus октаэдр с 6 шипами и 8 гранями.
Circopurus октаэдр с 6 шипами и 8 гранями.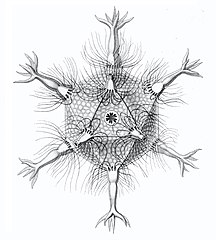 Икосаэдры циркогонии с 12 шипами и 20 гранями.
Икосаэдры циркогонии с 12 шипами и 20 гранями.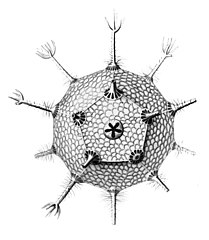 Циркоррегма додекаэдры с 20 (неполностью прорисованными) шипами и 12 гранями.
Циркоррегма додекаэдры с 20 (неполностью прорисованными) шипами и 12 гранями. Cannocapsa stethoscopium с 20 шипами.
Cannocapsa stethoscopium с 20 шипами.






В галерее представлены изображения радиолярий, извлеченные из рисунков немецкого зоолога и эрудита Эрнста Геккеля, сделанных в 1887 году.
- Тьюринг, Алан (1952). «Химические основы морфогенеза» (PDF) . Философские труды Лондонского королевского общества Б. 237 (641): 37–72. Бибкод : 1952РСПТБ.237...37Т . дои : 10.1098/rstb.1952.0012 . JSTOR 92463 . S2CID 120437796 .
- Ричардс, Бернард (2005–2006) «Тьюринг, Ричардс и морфогенез» , The Rutherford Journal , Том 1.
Ископаемая запись
[ редактировать ]
Самые ранние из известных радиолярий относятся к самому началу кембрийского периода и появляются в тех же слоях, что и первая мелкая ракушечная фауна ; они могут быть даже конечными докембрийскими по возрасту. [24] [25] [26] [27] Они имеют существенные отличия от более поздних радиолярий, с другой структурой решетки кремнезема и небольшим количеством шипов на тесте , если таковые имеются . [26] Около девяноста процентов известных видов радиолярий вымерли. Скелеты или тесты древних радиолярий используются в геологическом датировании , в том числе для разведки нефти и определения древнего климата . [28]
Некоторые распространенные окаменелости радиолярий включают Actinomma , Heliosphaera и Hexadoridium .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Смолли, Эй Джей (1963). «Радиолярии: построение шаровидного скелета». Наука . 140 (3565): 396–397. Бибкод : 1963Sci...140..396S . дои : 10.1126/science.140.3565.396 . ПМИД 17815802 . S2CID 28616246 .
- ^ Павловски Дж., Бурки Ф. (2009). «Распутывание филогении амебоидных простейших» . Дж. Эукариот. Микробиол . 56 (1): 16–25. дои : 10.1111/j.1550-7408.2008.00379.x . ПМИД 19335771 .
- ^ Юаса Т., Такахаши О., Хонда Д., Маяма С. (2005). «Филогенетический анализ полицистиновых радиолярий на основе последовательностей 18s рДНК Spumellarida и Nassellarida». Европейский журнал протистологии . 41 (4): 287–298. дои : 10.1016/j.ejop.2005.06.001 .
- ^ Николаев С.И., Берни С., Фарни Дж.Ф. и др. (май 2004 г.). «Закат Heliozoa и появление Rhizaria, новой супергруппы амебоидных эукариот» . Учеб. Натл. акад. наук. США . 101 (21): 8066–71. дои : 10.1073/pnas.0308602101 . ПМК 419558 . ПМИД 15148395 .
- ^ Крабберод А.К., Броте Дж., Долвен Дж.К. и др. (2011). «Радиолярии разделены на Polycystina и Spasmaria в комбинированной филогении 18S и 28S рДНК» . ПЛОС ОДИН . 6 (8): e23526. Бибкод : 2011PLoSO...623526K . дои : 10.1371/journal.pone.0023526 . ПМК 3154480 . ПМИД 21853146 .
- ^ Кавалер-Смит Т. (декабрь 1993 г.). «Царство простейших и его 18 типов» . Микробиол. Преподобный . 57 (4): 953–94. doi : 10.1128/mmbr.57.4.953-994.1993 . ПМК 372943 . ПМИД 8302218 .
- ^ Десель Дж., Сузуки Н., Маэ Ф., де Варгас К., Нот Ф. (май 2012 г.). «Молекулярная филогения и морфологическая эволюция акантарии (радиолярии)». Протист . 163 (3): 435–50. дои : 10.1016/j.protis.2011.10.002 . ПМИД 22154393 .
- ^ Не Ф., Гауслинг Р., Азам Ф., Гейдельберг Дж. Ф., Уорден А. З. (май 2007 г.). «Вертикальное распределение пикоэукариотического разнообразия в Саргассовом море». Окружающая среда. Микробиол . 9 (5): 1233–52. Бибкод : 2007EnvMi...9.1233N . дои : 10.1111/j.1462-2920.2007.01247.x . ПМИД 17472637 .
- ^ Кавалер-Смит Т. (июль 1999 г.). «Принципы нацеливания белков и липидов во вторичном симбиогенезе: происхождение эвгленоидных, динофлагеллятных и споровидных пластид и генеалогическое древо эукариот». Дж. Эукариот. Микробиол . 46 (4): 347–66. дои : 10.1111/j.1550-7408.1999.tb04614.x . ПМИД 18092388 . S2CID 22759799 .
- ^ Болтовской Д., Клинг С.А., Такахаши К. и Бьёрклунд К. (2010) «Мировой атлас распространения современных Polycystina (Radiolaria)». Электронная палеонтология , 13 : 1–230.
- ^ Кейси, Р.Э., Споу, Дж.М., и Кунце, Ф.Р. (1982) «Распределение полицистиновых радиолярий и улучшения, связанные с океанографическими условиями в гипотетическом океане». Являюсь. доц. Домашний питомец. геол. Бык. , 66 : 319–332.
- ^ Лазарь, Дэвид Б. (2011). «Глубоководные микроископаемые записи макроэволюционных изменений планктона и их изучение». Геологическое общество, Лондон, специальные публикации . 358 (1): 141–166. Бибкод : 2011ГСЛСП.358..141Л . дои : 10.1144/SP358.10 . S2CID 128826639 .
- ↑ Перейти обратно: Перейти обратно: а б Трубовиц, Сара; Лазарь, Дэвид; Реноди, Йохан; Ноубл, Паула Дж. (2020). «Морской планктон демонстрирует пороговое вымирание в ответ на изменение климата в неогеновом периоде» . Природные коммуникации . 11 (1): 5069. Бибкод : 2020NatCo..11.5069T . дои : 10.1038/s41467-020-18879-7 . ПМЦ 7582175 . ПМИД 33093493 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Васильев, Мэгги (2006) «Планктон — животный планктон» , Те Ара — Энциклопедия Новой Зеландии . Доступ: 2 ноября 2019 г.
- ^ Качович, Сара (2018) «Разум важнее методов: связь микрокаменелостей с тектоникой» Блог Отдела тектоники и структурной геологии Европейского союза геонаук.
- ^ Тьюринг, Алан; Сондерс, ПТ (1992). Морфогенез (на эсперанто). Амстердам: Северная Голландия. ISBN 978-0-08-093405-1 . OCLC 680063781 .
- ^ Ричардс, Бернард (2006) «Тьюринг, Ричардс и морфогенез» , The Rutherford Journal , Том 1.
- ^ Ричардс, Бернард (1954), «Морфогенез радиолярий», магистерская диссертация , Манчестер, Великобритания: Манчестерский университет .
- ^ Ричардс, Бернард (2005). «Тьюринг, Ричардс и морфогенез» . Резерфордский журнал . 1 .
- ↑ Перейти обратно: Перейти обратно: а б Ричардс, Бернард (2017). «Глава 35 – Радиолярии: проверка теории Тьюринга». В Коупленде, Джек ; и др. (ред.). Руководство Тьюринга . стр. 383–388.
- ^ Коупленд, Джек ; Боуэн, Джонатан ; Спревак, Марк; Уилсон, Робин ; и др. (2017). «Заметки об участниках». Руководство Тьюринга . Издательство Оксфордского университета . п. 478. ИСБН 978-0198747833 .
- ^ Варя, К.; Арагон, JL; Баррио, РА (1999). «Узоры Тьюринга на сфере». Физический обзор E . 60 (4): 4588–92. Бибкод : 1999PhRvE..60.4588V . дои : 10.1103/PhysRevE.60.4588 . ПМИД 11970318 .
- ^ Качович С., Шэн Дж. и Эйчисон Дж. К., 2019. Добавление нового измерения к исследованиям ранней эволюции радиолярий. Научные отчеты, 9(1), стр.1-10. два : 10.1038/s41598-019-42771-0 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Чанг, Шан; Фэн, Цинлай; Чжан, Лэй (14 августа 2018 г.). «Новые кремнистые микрофоссилии из терренёвской формации Яньцзяхэ, Южный Китай: возможные самые ранние ископаемые останки радиолярий». Журнал наук о Земле . 29 (4): 912–919. Бибкод : 2018JEaSc..29..912C . дои : 10.1007/s12583-017-0960-0 . S2CID 134890245 .
- ^ Чжан, Кэ; Фэн, Цин-Лай (сентябрь 2019 г.). «Раннекембрийские радиолярии и спикулы губок из формации Нюцзяохэ в Южном Китае». Палеомир . 28 (3): 234–242. дои : 10.1016/j.palwor.2019.04.001 . S2CID 146452469 .
- ↑ Перейти обратно: Перейти обратно: а б А. Браун; Дж. Чен; Д. Валошек; А. Маас (2007), «Первые раннекембрийские радиолярии», в Vickers-Rich, Patricia; Комарауэр, Патрисия (ред.), Взлет и падение эдиакарской биоты , Специальные публикации, том. 286, Лондон: Геологическое общество, стр. 143–149, номер документа : 10.1144/SP286.10 , ISBN. 978-1-86239-233-5 , OCLC 156823511
- ^ Малец, Йорг (июнь 2017 г.). «Идентификация предполагаемых нижнекембрийских радиолярий». Ревю микропалеонтологии . 60 (2): 233–240. Бибкод : 2017RvMic..60..233M . дои : 10.1016/j.revmic.2017.04.001 .
- ↑ Цукерман, Л.Д., Феллерс, Т.Дж., Альварадо, О. и Дэвидсон, М.В. «Радиолярии» , «Молекулярные выражения», Университет штата Флорида, 4 февраля 2004 г.
- Зеттлер, Линда А.; Согин, М.Л.; Кэрон, Д.А. (1997). «Филогенетические отношения между Acantharea и Polycystinea: молекулярный взгляд на радиолярии Геккеля» . Учеб. Натл. акад. наук. США . 94 (21): 11411–6. Бибкод : 1997PNAS...9411411A . дои : 10.1073/pnas.94.21.11411 . ПМК 23483 . ПМИД 9326623 .
- Лопес-Гарсия П., Родригес-Валера Ф., Морейра Д. (январь 2002 г.). «На пути к монофилии радиолярий Геккеля: данные об окружающей среде 18S рРНК подтверждают сестринство полицистинеи и акантарея». Мол. Биол. Эвол . 19 (1): 118–121. doi : 10.1093/oxfordjournals.molbev.a003976 . ПМИД 11752197 .
- Адл С.М., Симпсон А.Г., Фармер М.А. и др. (2005). «Новая классификация эукариот более высокого уровня с упором на таксономию протистов» . Дж. Эукариот. Микробиол . 52 (5): 399–451. дои : 10.1111/j.1550-7408.2005.00053.x . ПМИД 16248873 .
- Геккель, Эрнст (2005). Формы искусства из океана: Атлас радиолярий 1862 года . Мюнхен; Лондон: Престель Верлаг. ISBN 978-3-7913-3327-4 .
Внешние ссылки
[ редактировать ]- [1] Радиолярии
- Броди, К. (февраль 2005 г.). «Геометрия и закономерности в природе 3: дыры в тестах радиолярий и диатомей» . Микскейп (112). ISSN 1365-070X .
- Radiolaria.org
- Геккель, Эрнст (1862). Die Radiolarien ( Rhizopoda radiaria ) . Берлин. Архивировано из оригинала 19 июня 2009 г. Проверено 7 сентября 2007 г.
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - Радиолярии — капли
- Древо жизни — радиолярия
- ^ Болтовской, Деметрио; Андерсон, О. Роджер; Корреа, Нэнси М. (2016). «Радиолярия и Феодария». В Арчибальде, Джон М.; Симпсон, Аластер ГБ; Сламовиц, Клаудио Х.; Маргулис, Линн ; Мелконян, Михаил; Чепмен, Дэвид Дж.; Корлисс, Джон О. (ред.). Справочник протистов . Международное издательство Спрингер. стр. 1–33. дои : 10.1007/978-3-319-32669-6_19-1 . ISBN 978-3-319-32669-6 .