Голозоа
| Голозойцы Временной диапазон: | |
|---|---|
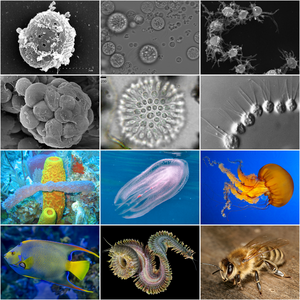 | |
| Голозойное разнообразие. В верхней половине представлены представители голозойных протистов. В нижней половине представлены представители многоклеточных животных. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | Аморфея |
| Клэйд : | Подчиняться |
| (без рейтинга): | Опистоконта |
| (без рейтинга): | Голозоа Ланг и др., 2002 г. [1] |
| Клады [4] | |
Неуверенное сиденье | |
| Синонимы | |
| |
Holozoa (от древнегреческого ὅλος (холос) «целый» и ζῷον (zoion) «животное») — это клад организмов, включающий животных и их ближайших одноклеточных родственников, но исключающий грибы и все другие организмы. Вместе они составляют более 1,5 миллиона видов чисто гетеротрофных организмов, в том числе около 300 одноклеточных видов. Он состоит из различных подгрупп, а именно Metazoa (или животных) и простейших Choanoflagellata , Filasterea , Pluriformea и Ichthyosporea . группами, Holozoa входит в состав Opisthokonta — супергруппы эукариот Наряду с грибами и некоторыми другими . Хоанофила ранее использовалась как название группы, сходной по составу с Holozoa, но сейчас ее использование не рекомендуется, поскольку она исключает животных и, следовательно, является парафилетической .
Голозойные протисты играют решающую роль в понимании эволюционных шагов, ведущих к появлению многоклеточных животных от одноклеточных предков. Недавние геномные исследования пролили свет на эволюционные взаимоотношения между различными линиями голозойных животных , открывая понимание происхождения многоклеточности . Некоторые окаменелости возможных многоклеточных животных были интерпретированы как голозойные протисты.
Характеристики
[ редактировать ]Состав
[ редактировать ]Holozoa — клада , включающая животных и их ближайших родственников, а также их общего предка , но исключающая грибы . Он определяется на основе отраслевого подхода как клада, охватывающая всех родственников Homo sapiens (животное), но не Neurospora crassa ( гриб ). [4] Holozoa, помимо животных, в основном включает в себя одноклеточные линии протистов различной морфологии, такие как хоанофлагелляты , филастерии , ихтиоспории , а также отдельные роды Corallochytrium , Syssomonas и Tunicaraptor . [6] [2]
- Choanoflagellata , насчитывающая около 250 видов. [7] являются ближайшими ныне живущими родственниками животных. Это свободноживущие одноклеточные или колониальные жгутиконосцы , питающиеся бактериями с помощью характерного «воротника» из микроворсинок . Воротник хоанофлагеллят очень напоминает клетки воротника губки . [8] что привело к возникновению теорий с 19 века об их родстве с губками . [9] Загадочная Proterospongia — пример колониальной хоанофлагелляты, которая, как считалось, связана с происхождением губок. [10] Сродство других одноклеточных голозойных начало признаваться только в 1990-х годах. [11]
- Ихтиоспореи , также известные как Mesomycetozoea и насчитывающие около 40 видов, в основном состоят из паразитов или комменсалов . Они взаимодействуют с самыми разными животными: от людей и рыб до морских беспозвоночных . Большинство из них размножаются многоядерными колониями и распространяются в виде жгутиковых или амеб . [7]
- Filasterea — группа из 6 видов амебоидов, принадлежащих к родам Minisisia , Pigoraptor , [6] Капсаспора и Чикиспора , [12] объединены строением своих нитевидных ложноножек . [13]
- Pluriformea — предварительное название клады, состоящей из двух видов Corallochytrium limacisporium и Syssomonas multiformis . Эти организмы имеют различную форму, включая скопления клеток, амебы, жгутиконосцы и амебофлагелляты. [6]
- Tunicaraptor unikontum — новейшая обнаруженная клада, положение которой внутри Holozoa еще предстоит определить. Это жгутиковые со специализированной структурой «рота», отсутствующей у других голозойных. [2]
- Многоклеточные организмы , известные как животные, представляют собой многоклеточные организмы, насчитывающие более 1,5 миллионов живых видов. [14] Для них характерна фаза бластулы в период эмбрионального развития и, за исключением аморфных губок , образование зародышевых листков и дифференцированных тканей . [4]
Генетика
[ редактировать ]Первым секвенированным одноклеточных голозой геномом был геном Monosiga brevicollis , хоанофлагелляты . Его размер составляет около 41,6 мегапар оснований (МБП) и содержит около 9200 кодирующих генов , что делает его сопоставимым по размеру с геномом нитчатых грибов . Геномы животных обычно больше (например, геном человека — 2900 Мбит/с; геном плодовой мухи — 180 Мбит/с), за некоторыми исключениями. [15]
Эволюция
[ редактировать ]Филогения
[ редактировать ]Holozoa, наряду с кладой, содержащей грибы и их родственников- протистов ( Holomycota ), являются частью более крупной супергруппы эукариот, известной как Opisthokonta . Holozoa отделились от своего предка-опистоконта около 1070 миллионов лет назад (млн лет назад). [16] Хоанофлагелляты, животные и филастерии группируются в кладу Filozoa . Внутри Filozoa хоанофлагелляты и животные группируются в кладу Choanozoa . [13] На основе филогенетического и филогеномного анализа кладограмма Holozoa показана ниже: [17] [18] [6] [2]
| Опистоконта |
| ||||||||||||||||||||||||||||||||||
| 1250 млн лет назад |

Неопределенность сохраняется вокруг взаимоотношений двух наиболее базальных групп — Ichthyosporea и Pluriformea . [4] Они могут быть сестрами друг друга, образуя предполагаемую кладу Teretosporea . [19] Альтернативно, Ichthyosporea может быть самым ранним ветвящимся из них, тогда как Pluriformea является сестрой клады Filozoa , состоящей из филастерий, хоанофлагеллят и животных. Этот второй результат получил еще большую поддержку после открытия Syssomonas . [2] [6]
Положение Туникараптора , нового члена голозоа, до сих пор не решено. три различных филогенетических положения Tunicaraptor В результате анализа были получены : как сестринская группа Filasterea , как сестра Filozoa или как самая базальная группа из всех Holozoa. [2] [20]
Экологические исследования ДНК океанов выявили новые разнообразные линии Holozoa. Большинство из них гнездится в пределах известных групп, главным образом Ichthyosporea и Choanoflagellata . Однако одна экологическая клада не гнездится ни в одной известной группе и представляет собой потенциальную новую линию голозоа. Предварительно он получил название МАШОЛ (что означает «морская маленькая голозоа»). [21]
Одноклеточное происхождение животных
[ редактировать ]
Стремление выяснить эволюционное происхождение животных от одноклеточного предка требует изучения перехода к многоклеточности . В отсутствие летописей окаменелостей, документирующих эту эволюцию, понимание одноклеточных предков животных получается благодаря анализу общих генов и генетических путей между животными и их ближайшими ныне живущими одноклеточными родственниками. Генетический состав этих одноклеточных голозойных животных позволил сделать важное открытие: многие генетические характеристики, ранее считавшиеся уникальными для животных, также могут быть обнаружены у этих одноклеточных родственников. Это говорит о том, что происхождение многоклеточных животных произошло не исключительно из-за появления новых генов (т.е. инноваций), а из-за ранее существовавших генов, которые были адаптированы или использованы по-новому (т.е. коопция). [7] [6] Например:
- Белки адгезии необходимы для того, чтобы клетки могли прилипать друг к другу и к внеклеточному матриксу , образуя слои и ткани у животных. Некоторые одноклеточные голозои, такие как хоанофлагелляты и филастерии , обладают генами, которые кодируют белки, участвующие в межклеточной адгезии и адгезии клеточного матрикса (например, кадгерин и интегрин соответственно). Однако другие гены, по-видимому, обнаруживаются исключительно у животных (например, β-катенин ). [7]
- Белки, родственные ЕСМ, участвующие в формировании внеклеточного матрикса , присутствуют и у других голозойных животных (например, ламинины , коллагены и фибронектинами ). [22]
- Белки сигнальной трансдукции являются еще одним требованием для многоклеточности многоклеточных животных. Некоторые цитоплазматические тирозинкиназы животных (такие как киназа фокальной адгезии ) и сигнальный путь Hippo присутствуют у одноклеточных голозойных. Другие сигнальные пути, высококонсервативные у животных (например, Hedgehog , WNT , TGFβ , JAK-STAT и Notch ), отсутствуют у других голозойных, но сходные сигнальные рецепторы развились независимо у хоанофлагеллят , филастерий и ихтиоспорий (например, рецепторы тирозинкиназ ). [7]
- Значительная часть животных транскрипционных факторов (TF) уже присутствует у одноклеточных голозойных, включая некоторые классы TF, которые ранее считались специфичными для животных (например, p53 и T-box ). [7]
Кроме того, многие биологические процессы, наблюдаемые у животных, уже присутствуют у их одноклеточных родственников, такие как половое размножение и гаметогенез у хоанофлагелляты Salpingoeca Rosetta и несколько типов многоклеточной дифференцировки . [7]
Ископаемая запись
[ редактировать ]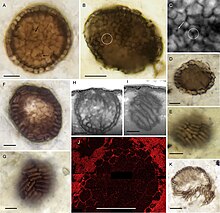
Пресноводное возрастом в миллиард лет возможно , микроскопическое ископаемое под названием Bicellum brasieri, является самым ранним из известных голозойных животных. На нем показаны два дифференцированных типа клеток или стадии жизненного цикла . Он состоит из сферического шара плотно упакованных клеток (стереобластов), заключенных в один слой удлиненных клеток . Существуют также две популяции стереобластов смешанной формы, которые интерпретируются как миграция клеток на периферию, движение, которое можно объяснить дифференциальной межклеточной адгезией . Эти явления согласуются с современными одноклеточными голозойными, которые, как известно, образуют многоклеточные стадии в сложных жизненных циклах. [3]
Предполагаемые эдиакарские ископаемые « эмбрионы » ранних многоклеточных животных , обнаруженные в формации Доушантуо , были переинтерпретированы как неживотные протисты внутри Holozoa. По мнению некоторых авторов, хотя они и представляют возможное эмбриональное дробление , у них отсутствуют синапоморфии многоклеточных животных , такие как дифференциация тканей и близлежащие молодые или взрослые особи. Вместо этого его развитие сравнимо со стадией прорастания голозойных животных неживотных. Возможно, они представляют собой эволюционную ступень , на которой палинтомное дробление (т.е. быстрое деление клеток без цитоплазмы промежуточного роста , характерное для эмбрионального дробления животных ) [23] был метод расселения и распространения . [24]
Таксономия
[ редактировать ]История
[ редактировать ]До 2002 года связь между Choanoflagellata , Ichthyosporea и дивергенцией животных и грибов рассматривалась на основе морфологии и ультраструктуры . Ранние филогенетические анализы дали противоречивые результаты, поскольку количество доступных последовательностей ДНК было недостаточным для получения однозначных результатов. Таксономическая неопределенность была такова, что, например, некоторые ихтиоспории традиционно рассматривались как трихомицетовые грибы. [1]
Holozoa была впервые признана кладой в 2002 году посредством филогеномного анализа, проведенного Францем Берндом Лангом, Чарльзом Дж. О'Келли и другими сотрудниками в рамках статьи , опубликованной в журнале Current Biology . В исследовании использовались полные митохондриальные геномы хоанофлагелляты ( Monosiga brevicollis ) и ихтиоспории ( Amoebidium parasiticum ), чтобы твердо определить положение ихтиоспории как сестринской группы Choanoflagellata+Metazoa. Эта клада была названа Holozoa (от древнегреческого ὅλος (holos) «целое» и ζῷον (zoion) «животное»), что означает «целое животное», ссылаясь на более широкое животное происхождение, которое она содержит. [1]
С тех пор все апостериорные анализы подтверждают, что Holozoa является надежной кладой. [20] даже после открытия большего количества гнездящихся в нем таксонов (а именно Filasterea с 2008 г., [13] и многоформные виды Corallochytrium и Syssomonas с 2014 г. [25] и 2017 год [6] соответственно). По состоянию на 2019 год клада принята Международным обществом протистологов, которое пересматривает классификацию эукариот. [4]
Классификация
[ редактировать ]В классификациях, использующих традиционные таксономические ранги (например, царство, тип, класс), все голозойные протисты относят к подтипу Choanofila (тип Choanozoa , [а] царство Protozoa ), а животных относят к отдельному царству Metazoa или Animalia. [26] Эта классификация исключает животных, хотя они происходят от того же общего предка, что и простейшие хоанофиланы, что делает их парафилетической группой, а не настоящей кладой. Современные кладистические подходы к эукариот классификации отдают предпочтение монофилетическим группировкам над традиционными рангами, которые все чаще воспринимаются как излишние и ненужные. Поскольку Holozoa является кладой, ее использование предпочтительнее парафилетического таксона Choanophila. [4]
- Холозоа Ланг и др. 2002 г.
- Неопределенные места : † Bicellum brazieri Strother & Wellman 2021 [3]
- Tunicaraptor Tikhonenkov, Mikhailov, Hehenberger, Karpov, Prokina, Esaulov, Belyakova, Mazei, Mylnikov, Aleoshin & Keeling 2020 [2]
- Ichthyosporea Cavalier-Smith 1998 [Mesomycetozoea Mendoza et al. 2002 ]
- Дермоцистида Кавальер-Смит 1998 г.
- Ихтиофонида Кавальер-Смит 1998 г.
- Pluriformea Hehenberger et al. 2017 год
- Кораллохитриум Рагу-Кумар 1987 г.
- Сиссомонас Тихоненков, Хехенбергер, Мыльников и Килинг 2017
- Philozoa Shalchian-Tabrizi et al. 2008 год
- Филастерия Шалчиан-Табризи и др. 2008 год
- Капсаспора Хертель, Бэйн и Локер, 2002 г.
- Министерия Паттерсон и др. 1993 год
- Pigoraptor Tikhonenkov et al. 2017
- Чикиспора Уррутия, Файст и Бас 2022 [12]
- Choanozoa Brunet & King 2017 [Choanozoa Cavalier-Smith et al. 1991 (П)] [а]
- Choanoflagellata Kent 1880–1882 [Choanoflagellatea Cavalier-Smith 1997, исправление. Кавалер-Смит 1998 ]
- Краспедида Кавальер-Смит, 1997, исправлено. Ниче и др. 2011 год
- Acanthoecida Cavalier-Smith 1997, исправлено. Ниче и др. 2011 год
- Metazoa Haeckel 1874, исправлено. Аддл и др. 2005 [Животные Линней 1758 ]
- Choanoflagellata Kent 1880–1882 [Choanoflagellatea Cavalier-Smith 1997, исправление. Кавалер-Смит 1998 ]
- Филастерия Шалчиан-Табризи и др. 2008 год
Примечания
[ редактировать ]- ^ Перейти обратно: а б Термин « Choanozoa » использовался с 1991 года Кавальер-Смитом для обозначения парафилетического типа опистоконтных простейших. [27] а термины « Apoikozoa » и «choanoflagellata» были предложены в качестве названий клады Metazoa + Choanoflagellata . Однако эти термины не были официально описаны или приняты и были отвергнуты в пользу переименованного Choanozoa, чтобы соответствовать кладе Metazoa + Choanoflagellata. [4]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Ланг Б.Ф., О'Келли С., Нерад Т., Грей М.В., Бургер Дж. (2002). «Ближайшие одноклеточные родственники животных» . Современная биология . 12 (20): 1773–1778. дои : 10.1016/S0960-9822(02)01187-9 . ПМИД 12401173 .
- ^ Перейти обратно: а б с д и ж г Тихоненков Д.В., Михайлов К.В., Хехенбергер Е., Мыльников А.П., Алеошин В.В., Килинг П.Дж. и др. (2020). «Новая линия микробных хищников усложняет реконструкцию эволюционного происхождения животных» . Современная биология . 30 (22): 4500–4509. дои : 10.1016/j.cub.2020.08.061 . ПМИД 32976804 .
- ^ Перейти обратно: а б с Стротер, Пол К.; Брейзер, Мартин Д.; Уэйси, Дэвид; Тимпе, Лесли; Сондерс, Мартин; Веллман, Чарльз Х. (апрель 2021 г.). «Возможное голозойное животное возрастом в миллиард лет с дифференцированной многоклеточностью» . Современная биология . 31 (12): 2658–2665.e2. дои : 10.1016/j.cub.2021.03.051 . ПМИД 33852871 .
- ^ Перейти обратно: а б с д и ж г Адл С.М., Басс Д., Лейн CE, Лукеш Дж., Шох К.Л., Смирнов А., Агата С., Берни С., Браун М.В., Бурки Ф., Карденас П., Чепичка И., Чистякова Л., дель Кампо Дж., Данторн М., Эдвардсен Б., Эглит. Ю, Гийу Л., Хампл В., Хейсс А.А., Хоппенрат М., Джеймс Тай, Карнковска А., Карпов С., Ким Е., Колиско М., Кудрявцев А., Лар DJG, Лара Е., Ле Галл Л., Линн Д.Х., Манн Д.Г., Массана Р. , Митчелл И.А.Д., Морроу С., Парк Дж.С., Павловски Дж.В., Пауэлл М.Дж., Рихтер Д.Д., Рюкерт С., Шедвик Л., Шимано С., Шпигель Ф.В., Торруэлла Г., Юсеф Н., Златогурский В., Чжан К. (2019). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ПМК 6492006 . ПМИД 30257078 .
- ^ Кавалер-Смит, Томас (2009). «Мегафилогения, планы клеточного тела, адаптивные зоны: причины и время базального излучения эукариот» . Журнал эукариотической микробиологии . 56 : 26–33. дои : 10.1111/j.1550-7408.2008.00373.x .
- ^ Перейти обратно: а б с д и ж г Хехенбергер, Элизабет; Тихоненков Денис Владимирович; Колиско, Мартин; Кампо, Хавьер дель; Есаулов Антон С.; Мыльников Александр П.; Килинг, Патрик Дж. (2017). «Новые хищники меняют филогению голозойных животных и обнаруживают наличие двухкомпонентной сигнальной системы у предков животных» . Современная биология . 27 (13): 2043–2050.e6. дои : 10.1016/j.cub.2017.06.006 . ПМИД 28648822 .
- ^ Перейти обратно: а б с д и ж г Себе-Педрос А, Дегнан Б, Руис-Трилло I (2017). «Происхождение Metazoa: одноклеточный взгляд». Обзоры природы Генетика . 18 (8): 498–512. дои : 10.1038/nrg.2017.21 . ПМИД 28479598 . S2CID 30709486 .
- ^ Стинкамп, Эмма Т.; Райт, Джейн; Балдауф, Сандра Л. (январь 2006 г.). «Протистанское происхождение животных и грибов» . Молекулярная биология и эволюция . 23 (1): 93–106. дои : 10.1093/molbev/msj011 . ПМИД 16151185 .
- ^ Симпсон AGB, Сламовиц CH, Арчибальд Дж. М. (2017). «Глава 1. Разнообразие протистов и филогения эукариот». В Арчибальде Дж. М., Симпсоне А.Г.Б., Сламовице Ч.Х. (ред.). Справочник протистов . Том. 1 (2-е изд.). Международное издательство Спрингер. стр. 1–22. ISBN 978-3-319-28147-6 .
- ^ Брюне Т., Кинг Н. (2022). «Одноклеточные предки животных: история гипотез». В Herron MD, Conlin PL, Ratcliff WC (ред.). Эволюция многоклеточности . Эволюционная клеточная биология. ЦРК Пресс. стр. 251–278. дои : 10.1201/9780429351907-17 . ISBN 9780429351907 .
- ^ Рэган, Марк А.; Гоггин, К. Луиза; Коуторн, Ричард Дж.; Церениус, Лаге; Джеймисон, Анджела ВК; Плурд, Сьюзен М.; Рэнд, Томас Г.; Сёдерхалл, Кеннет; Гутелл, Робин Р. (15 октября 1996 г.). «Новая клада протистанских паразитов, близкая к дивергенции животных и грибов» . ПНАС . 93 (21): 11907–11912. Бибкод : 1996PNAS...9311907R . дои : 10.1073/pnas.93.21.11907 . ПМЦ 38157 . ПМИД 8876236 .
- ^ Перейти обратно: а б Уррутиа А., Мици К., Фостер Р., Росс С., Карр М., Уорд Г.М. и др. (2022). « Txikispora philomaios n. sp., ng, микроэукариотический патоген амфипод, обнаруживает паразитизм и скрытое разнообразие в классе Filasterea» . Журнал эукариотической микробиологии . 69 (2): e12875. дои : 10.1111/jeu.12875 . ПМИД 34726818 . S2CID 240422937 .
- ^ Перейти обратно: а б с Шалчиан-Тебризи, Кямран; Минге, Марианна А.; Эспелунд, Мэри; Орр, Рассел; Руден, Торгейр; Якобсен, Кжетилл С.; Кавальер-Смит, Томас ; Арамайо, Родольфо (7 мая 2008 г.). Арамайо, Родольфо (ред.). «Мультигенная филогения хоанозоа и происхождение животных» . ПЛОС ОДИН . 3 (5): e2098. Бибкод : 2008PLoSO...3.2098S . дои : 10.1371/journal.pone.0002098 . ПМК 2346548 . ПМИД 18461162 .
- ^ Чжан, Чжи-Цян (2013). «Биоразнообразие животных: обновление классификации и разнообразия в 2013+» . Зоотакса . 3703 (1): 5–11. дои : 10.11646/zootaxa.3703.1.3 .
- ^ Кинг Н., Уэстбрук М., Янг С. и др. (2008). «Геном хоанофлагелляты Monosiga brevicollis и происхождение многоклеточных животных» . Природа . 451 : 783–788. дои : 10.1038/nature06617 . hdl : 2027.42/62649 .
- ^ Лавал Х.М., Шильде С., Кин К. и др. (2020). «Адаптация к холодному климату является вероятной причиной эволюции многоклеточных споруляций у Dictyostelia » . Научные отчеты . 10 : 8797. doi : 10.1038/s41598-020-65709-3 . ПМК 7260361 .
- ^ Парфри, Лаура Вегенер ; Лар, Дэниел Дж.Г.; Нолл, Эндрю Х.; Кац, Лаура А. (16 августа 2011 г.). «Оценка времени ранней диверсификации эукариот с помощью мультигенных молекулярных часов» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (33): 13624–13629. Бибкод : 2011PNAS..10813624P . дои : 10.1073/pnas.1110633108 . ПМК 3158185 . ПМИД 21810989 .
- ^ Торруэлла, Гифре; де Мендоса, Алекс; Грау-Бове, Ксавье; Антон, Меритчелл; Чаплин, Марк А.; дель Кампо, Хавьер; Эме, Лаура; Перес-Кордон, Грегорио; Уиппс, Кристофер М. (21 сентября 2015 г.). «Филогеномика раскрывает конвергентную эволюцию образа жизни у близких родственников животных и грибов» . Современная биология . 25 (18): 2404–2410. дои : 10.1016/j.cub.2015.07.053 . ISSN 0960-9822 . ПМИД 26365255 .
- ^ Грау-Бове, Ксавье; Торруэлла, Гифре; Доначи, Стюарт; Суга, Хироши; Леонард, Гай; Ричардс, Томас А; Руис-Трилло, Иньяки (2017). «Динамика геномных инноваций в предках одноклеточных животных» . электронная жизнь . 6 : e26036. дои : 10.7554/eLife.26036 . ПМК 5560861 .
- ^ Перейти обратно: а б Рос-Роше Н., Перес-Посада А., Мишель Л.М., Руис-Трилло I (февраль 2021 г.). «Происхождение животных: предковая реконструкция перехода от одноклеточного к многоклеточному» . Открытая Биол . 11 (2): 200359. doi : 10.1098/rsob.200359 . hdl : 10261/251922 . ПМИД 33622103 .
- ^ Арройо, Алисия С; Ланн, Ромен; Баптест, Эрик; Руис-Трилло, Иньяки (сентябрь 2020 г.). «Сети сходства генов раскрывают потенциальную новую одноклеточную группу, тесно связанную с животными из экспедиции в океаны Тары » . Геномная биология и эволюция . 12 (9): 1664–1678. дои : 10.1093/gbe/evaa117 . ПМК 7533066 .
- ^ Суга Х, Чен З, де Мендоса А, Себе-Педрос А, Браун М.В., Крамер Е, Карр М, Кернер П, Вервут М, Санчес-Понс Н, Торруэлла Г, Дерелле Р., Мэннинг Г, Ланг Б.Ф., Расс С, Хаас Б.Дж., Роджер А.Дж., Нусбаум С., Руис-Трилло I (2013). « Геном Capsaspora раскрывает сложную предысторию одноклеточных животных» . Природные коммуникации . 4 (2325): 2325. Бибкод : 2013NatCo...4.2325S . дои : 10.1038/ncomms3325 . ПМЦ 3753549 . ПМИД 23942320 .
- ^ Чен Л., Сяо С., Пан К., Чжоу С., Юань Икс (сентябрь 2014 г.). «Дифференцировка клеток и разделение зародышей и сомы в окаменелостях, похожих на эмбрионы эдиакарских животных». Природа . 516 (7530): 238–241. Бибкод : 2014Natur.516..238C . дои : 10.1038/nature13766 . ПМИД 25252979 . S2CID 4448316 .
- ^ Хульдтгрен Т., Каннингем Дж.А., Инь С., Стампанони М., Мароне Ф., Донохью PCJ, Бенгтсон С. (2011). «Ископаемые ядра и структуры прорастания идентифицируют эдиакарские «эмбрионы животных» как инцистирующие протисты». Наука . 334 (6063): 1696–1699. Бибкод : 2011Sci...334Q1696H . дои : 10.1126/science.1209537 . ПМИД 22194575 . S2CID 39813961 .
- ^ Торруэлья Дж., де Мендоса А., Грау-Бове Х., Доначи С., Перес-Кордон Г., Ситха-Бобадилья А., Пейли Р., Манохар К.С., Николс К., Эме Л., дель Кампо Дж. (2014). «Филотранскриптомика выявляет древние и конвергентные черты Corallochytrium и Ministryia (Holozoa, Opisthokonta)». Филогения и эволюционные перспективы протистов Opisthokonta (PDF) (кандидатская диссертация). Полет. 75. Университет Барселоны. стр. 1–9.
- ^ Кавалер-Смит Т. (май 2022 г.). «Эволюция ресничной переходной зоны и корень дерева эукариот: значение для происхождения опистоконтов и классификации царств простейших, растений и грибов» . Протоплазма . 259 (3): 487–593. дои : 10.1007/s00709-021-01665-7 . ПМК 9010356 . ПМИД 34940909 .
- ^ Кавалер-Смит Т. (май 2013 г.). «Ранняя эволюция способов питания эукариот, структурное разнообразие клеток и классификация типов простейших Loukozoa, Sulcozoa и Choanozoa». Европейский журнал протистологии . 49 (2): 115–178. дои : 10.1016/j.ejop.2012.06.001 . ПМИД 23085100 .