Хоанофлагеллята
| Хоанофлагелляты Временной диапазон: известны только возможные окаменелости . Из мелового периода ( сеномана / турона ) [1] [2] | |
|---|---|
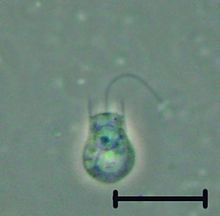 | |
| Кодосига сп. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | Аморфея |
| Клэйд : | Подчиняться |
| (без рейтинга): | Опистоконта |
| (без рейтинга): | Голозоа |
| (без рейтинга): | Философия |
| Клэйд : | Чоанозоа |
| Сорт: | Хоанофлагеллята Кент , 1880–1882 гг. [3] [4] |
| Типовой вид | |
| Моносига бревиколлис [5] | |
| Ордена и семьи | |
| Синонимы | |
| |
Хоанофлагелляты которые — группа свободноживущих одноклеточных и колониальных жгутиковых эукариот, считаются ближайшими ныне живущими родственниками животных . Хоанофлагелляты — это жгутики с воротником, имеющие воронкообразный воротник из взаимосвязанных микроворсинок у основания жгутика . Хоанофлагелляты способны как к бесполому , так и к половому размножению. [9] Они имеют характерную клеточную морфологию, характеризующуюся яйцевидным или сферическим телом клетки диаметром 3–10 мкм с единственным апикальным жгутиком, окруженным воротником из 30–40 микроворсинок (см. Рисунок). Движение жгутика создает потоки воды, которые могут продвигать свободно плавающие хоанофлагелляты через толщу воды и задерживать бактерии и детрит на воротнике микроворсинок, где эти пищевые продукты поглощаются. Такое питание обеспечивает важнейшее звено в глобальном углеродном цикле , связывающее трофические уровни . Помимо своей важной экологической роли, хоанофлагелляты представляют особый интерес для биологов-эволюционистов, изучающих происхождение многоклеточности у животных. Как ближайшие ныне живущие родственники животных, хоанофлагелляты служат полезной моделью для реконструкции последнего одноклеточного предка животных.
Этимология
[ редактировать ]Хоанофлагеллята — гибридное слово от греческого χοάνη khoánē, означающего « воронка » (из-за формы воротника) и латинского слова flagellum (отсюда английское flagellum ). [ нужна ссылка ]
Появление
[ редактировать ]
Каждая хоанофлагеллята имеет единственный жгутик , окруженный кольцом заполненных актином выступов, называемых микроворсинками , образующих цилиндрический или конический воротник ( хоанос по-гречески). Движение жгутика вытягивает воду через воротник, а бактерии и детрит захватываются микроворсинками и проглатываются. [10] Потоки воды, генерируемые жгутиком, также толкают свободно плавающие клетки, как и в животных сперме . Напротив, большинство других жгутиконосцев тянутся за свои жгутики. [ нужна ссылка ]
Помимо характерного для хоанофлагеллят единственного апикального жгутика, окруженного заполненными актином микроворсинками, внутренняя организация органелл в цитоплазме постоянна. [11] жгутика Базальное тельце расположено у основания апикального жгутика, а второе, нежгутиковое базальное тельце лежит под прямым углом к основанию жгутика. Ядро . занимает в клетке апикально-центральное положение, а пищевые вакуоли располагаются в базальной области цитоплазмы [11] [12] Кроме того, тело клетки многих хоанофлагеллят окружено отличительным внеклеточным матриксом или перипластом . Эти клеточные покрытия сильно различаются по структуре и составу и используются систематиками для целей классификации. Многие хоанофлагелляты строят сложные корзинчатые «дома», называемые лориками , из нескольких склеенных вместе полосок кремнезема. [11] Функциональное значение перипласта неизвестно, но считается, что у сидячих организмов он способствует прикреплению к субстрату. Есть предположение, что у планктонных организмов перипласт увеличивает сопротивление, тем самым противодействуя силе, создаваемой жгутиком, и повышая эффективность питания. [13]
Хоанофлагелляты либо свободно плавают в толще воды, либо сидячие , прикрепляясь к субстрату непосредственно или через перипласт или тонкую ножку. [14] Хотя хоанофлагелляты считаются строго свободноживущими и гетеротрофными , ряд родственников хоанофлагеллят, таких как представители Ichthyosporea или Mesomycetozoa , ведут паразитический или патогенный образ жизни. [15] История жизни хоанофлагеллят мало изучена. Считается, что многие виды ведут одиночный образ жизни; однако колониальность, по-видимому, несколько раз возникала независимо внутри группы, и колониальные виды все еще сохраняют одиночную стадию. [14]
Экология
[ редактировать ]
Более 125 современных видов хоанофлагеллят. [10] известны, распространены по всему миру в морских , солоноватых и пресноводных средах от Арктики до тропиков, занимая как пелагическую , так и донную зоны. Хотя большая часть выборок хоанофлагеллят была собрана на глубине от 0 до 25 м (от 0 до 82 футов), они были обнаружены на глубине до 300 м (980 футов) в открытой воде. [16] и 100 м (330 футов) под ледяными щитами Антарктики. [17] Предполагается, что многие виды являются космополитами в глобальном масштабе [например, Diaphanoeca grandis зарегистрирована в Северной Америке , Европе и Австралии (OBIS)], в то время как другие виды, как сообщается, имеют ограниченное региональное распространение. [18] Совместно распространенные виды хоанофлагеллят могут занимать совершенно разные микроокружения, но в целом факторы, влияющие на распределение и дисперсию хоанофлагеллят, еще предстоит выяснить. [ нужна ссылка ]
Ряд видов , например представители рода Proterospongia , образуют простые колонии . [10] планктонные комки, напоминающие миниатюрную гроздь винограда, в которой каждая клетка колонии снабжена жгутиками, или скопления клеток на одном стебле. [11] [19] В октябре 2019 года ученые обнаружили новое полосовое поведение хоанофлагеллят: они, по-видимому, могут координироваться, чтобы реагировать на свет. [20]
Хоанофлагелляты питаются бактериями и связывают недоступные иначе формы углерода с организмами, стоящими выше в трофической цепи. [21] Даже сегодня они играют важную роль в углеродном цикле и микробной пищевой сети . [10] Есть некоторые свидетельства того, что хоанофлагелляты также питаются вирусами. [22]
Жизненный цикл
[ редактировать ]
Хоанофлагелляты растут вегетативно, при этом многие виды подвергаются продольному делению; [12] однако репродуктивный жизненный цикл хоанофлагеллят еще предстоит выяснить. В статье, опубликованной в августе 2017 года, показано, что изменения окружающей среды, включая присутствие определенных бактерий, вызывают роение и последующее половое размножение хоанофлагеллят. [9] Уровень плоидности неизвестен; [24] однако открытие как ретротранспозонов, так и ключевых генов, участвующих в мейозе [25] ранее предположили, что они использовали половое размножение как часть своего жизненного цикла. Некоторые хоанофлагелляты могут подвергаться инцистации, которая включает втягивание жгутика и воротника и заключение в электронно-плотную фибриллярную стенку. При переносе на свежие среды происходит эксцистирование; хотя это еще предстоит наблюдать непосредственно. [26]
Доказательства полового размножения были зарегистрированы у видов хоанофлагеллят Salpingoeca Rosetta . [27] [28] Также сообщалось о наличии консервативных мейотических генов у хоанофлагеллят Monosiga brevicollis и Monosiga ovata . [29]
Биоминерализация кремния
[ редактировать ]Хоанофлагелляты Acanthoecid образуют внеклеточную корзинчатую структуру, известную как лорика. Лорика состоит из отдельных реберных полосок, состоящих из кремнеземно-белкового биокомпозита. Каждая реберная полоска формируется внутри клетки хоанофлагелляты и затем секретируется на поверхность клетки. У голых хоанофлагеллят сборка лорики происходит с использованием нескольких щупалец после того, как образуется достаточное количество реберных полосок, чтобы составить полную лорику. У тектиформных хоанофлагеллят реберные полоски собраны в ряд под воротником. Во время клеточного деления новая клетка берет эти реберные полоски в ходе цитокинеза и собирает свою собственную лорику, используя только эти ранее произведенные полоски. [30]
Биосиликатизация хоанофлагеллят требует концентрации кремниевой кислоты внутри клетки. Это осуществляется белками -переносчиками кремния (SiT). Анализ SiT хоанофлагеллят показывает, что они подобны переносчикам кремния типа SiT диатомовых водорослей образующих кремнезем и других страменопилей, . Семейство генов SiT практически не имеет гомологии с другими генами, даже с генами некремнистых хоанофлагеллят или страменопилей. Это указывает на то, что семейство генов SiT эволюционировало посредством бокового переноса генов между Acanthoecids и Stramenopiles. Это замечательный случай горизонтального переноса генов между двумя отдаленно родственными группами эукариот, который дал ключ к разгадке биохимии и кремний-белковых взаимодействий уникального семейства генов SiT. [31]
Классификация
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( февраль 2021 г. ) |
Связь с многоклеточными животными
[ редактировать ]Феликс Дюжарден , французский биолог, интересующийся эволюцией простейших, зафиксировал морфологическое сходство хоанофлагеллят и хоаноцитов губок и предположил возможность тесного родства еще в 1841 году. [13] За последнее десятилетие эта гипотетическая связь между хоанофлагеллятами и животными была подтверждена независимыми анализами множества несвязанных генетических последовательностей: 18S рДНК, генов, кодирующих ядерные белки, и митохондриальных геномов (Steenkamp et al., 2006; Burger et al. , 2003; [15] Уэйнрайт и др., 1993). Важно отметить, что сравнение последовательностей митохондриального генома хоанофлагеллят и трех губок подтверждает размещение хоанофлагеллят в качестве внешней группы по отношению к Metazoa (животным, также известным как Animalia ) и отрицает возможность того, что хоанофлагелляты произошли от многоклеточных животных (Лавров и др., 2005). Наконец, исследование генов, экспрессируемых в хоанофлагеллятах в 2001 году, показало, что хоанофлагелляты синтезируют гомологи генов передачи сигналов и адгезии клеток многоклеточных животных. [32] Секвенирование генома показывает, что среди живых организмов хоанофлагелляты наиболее тесно связаны с животными. [10] Поскольку хоанофлагелляты и многоклеточные животные тесно связаны, сравнение между этими двумя группами обещает дать представление о биологии их последнего общего предка и самых ранних событиях в эволюции многоклеточных животных . Хоаноциты (считающиеся одними из (также известные как «клетки с воротником») губок самых базальных многоклеточных животных) имеют ту же основную структуру, что и хоанофлагелляты. Ошейниковые клетки встречаются и у других групп животных, например у ленточных червей , [33] предполагая, что это была морфология их последнего общего предка. Последний общий предок животных и хоанофлагеллят был одноклеточным, возможно, образующим простые колонии; напротив, последним общим предком всех эуметозойных животных был многоклеточный организм с дифференцированными тканями, определенным «планом тела» и эмбриональным развитием (включая гаструляцию). [10] Время разделения этих линий трудно определить, но, вероятно, это произошло в позднем докембрии, > 600 миллионов лет назад . [10]
Внешние взаимоотношения хоанофлагеллятеи. [34]
| Опистоконта |
| ||||||
Филогенетические отношения
[ редактировать ]Хоанофлагелляты были включены в состав Chrysophyceae до Hibberd, 1975. [35] Недавняя молекулярно -филогенетическая реконструкция внутренних взаимоотношений хоанофлагеллят позволяет поляризовать эволюцию признаков внутри клады. Большие фрагменты ядерной SSU и LSU рибосомальной РНК , альфа-тубулина и генов, кодирующих белок теплового шока 90, были использованы для разрешения внутренних взаимоотношений и полярности признаков внутри хоанофлагеллят. [19] Каждый из четырех генов независимо показал схожие результаты, а анализ объединенного набора данных ( объединенных ) вместе с последовательностями других близкородственных видов ( животных и грибов ) демонстрирует, что хоанофлагелляты считаются монофилетическими , и подтверждает их положение как наиболее близких из известных одноклеточных живых существ. родственник животных.
Ранее Choanoflagellida были разделены на эти три семейства в зависимости от состава и строения их перипласта: Codonosigidae, Salpingoecidae и Acanthoecidae. При исследовании с помощью световой микроскопии у представителей семейства Codonosigidae перипласт отсутствует, но может иметь тонкий внешний слой, видимый только при электронной микроскопии . Семейство Salpingoecidae состоит из видов, клетки которых заключены в твердую теку, видимую как в световую, так и в электронную микроскопию. Тека целлюлозы представляет собой секретируемую оболочку, состоящую преимущественно из или других полисахаридов . [36] Теперь известно, что эти подразделения являются парафилетическими , с широко распространенной конвергентной эволюцией этих форм. Третье семейство хоанофлагеллят, Acanthoecidae, считается монофилетической группой. Эта клада обладает синапоморфией клеток, находящихся внутри корзинчатой лорики, что дает альтернативное название «лорикатные хоанофлагелляты». Acanthoecid lorica состоит из серии кремнистых реберных полосок, расположенных в видоспецифическом узоре lorica». [11] [13]
Дерево хоанофлагеллят, основанное на молекулярной филогенетике, делится на три хорошо поддерживаемые клады . [19] Клада 1 и клада 2 состоят из комбинации видов, традиционно относимых к Codonosigidae и Salpingoecidae, тогда как клада 3 включает виды из группы, таксономически классифицируемой как Acanthoecidae. [19] Картирование признаков характера на эту филогению указывает на то, что последним общим предком хоанофлагеллят был морской организм с дифференцированным жизненным циклом с малоподвижной и подвижной стадиями. [19]
Таксономия
[ редактировать ]хоанофлагелляты; [8]
- Орден Краспедида Кавальер-Смит 1997г.эм. Ниче и др. 2011 г.
- Семья Salpingoecidae Кент 1880-1882 гг.
- ? Дикраспеделла Эллис 1930 г.
- ? Диплоека Эллис 1930 г.
- ? Диплозигопсис Франция 1897 г.
- ? Пачисоека Эллис 1930 г.
- ? Пиропсис Менье 1910
- ? Салпингориза Клуг 1936 г.
- ? Сфаэродендрон Жуков, Мыльников и Моисеев 1976 г., нон Зееманн 1865 г.
- ? Стелексомонас Лакей 1942 г.
- Астросига Кент 1880-1882 гг.
- Ауломонас Лакей 1942 г.
- Чоаноэка Эллис 1930 г.
- Кладоспонгия Айенгар и Раматан, 1940 г.
- Кодоносигопсис Сенн 1900
- Диплосига Френцель 1891 г.
- Хартаэтосига Карр, Рихтер и Ниче 2017
- Мылносига Карр, Рихтер и Ниче 2017
- Лагеноэка Кент 1881 г.
- Микростомоэка Карр, Рихтер и Ниче 2017
- Парамоносига Джек, Арндт и Ниче, 2014 г.
- Салпингоека Джеймс-Кларк 1868 г., но не Эллис 1933 г.
- Стагондоека Карр, Рихтер и Ниче 2017
- Семейство Codonosigaceae Кент 1880-1882 гг.
- Кодосига Джеймс-Кларк 1866 г.
- Десмарелла Кент 1880-1882 гг.
- Кентросига Шиллер 1953
- Моносига Кент 1880-1882 гг.
- Протероспонгия Кент 1882 г.
- Sphaeroeca Лаутерборн 1894 г., а не Мейрик 1895 г.
- Стилохромонас Лакей 1940 г.
- Семья Salpingoecidae Кент 1880-1882 гг.
- Отряд Acanthoecida Norris 1965 г.эм. Ниче и др. 2011 (Хоанофлагелляты лорикатные)
- Кониоэка Томсен и Остергаард 2019
- Семейство Acanthoecidae Norris 1965 г. Ниче и др. 2011 (Голодные хоанофлагелляты)
- Семейство Stephanoecidae Leadbeater 2011 (Tectiform choanoflagellates)
- ? Конион Томсен 1982 г.
- ? Спиралоецион Маршан и Перрен, 1986 г.
- Акантокорбис Хара и Такахаши, 1984 г.
- Аменоскоп Хара и Такахаши, 1987 г.
- Афелоцион Томсен 1983
- Бикоста Ледбитер 1978
- Каллиаканта Ледбитер 1978
- Calotheca Thomsen & Moestrup 1983, не Desv. 1810 монахиня Спренг. 1817 г. монахиня Гейден 1887 г.
- Космоэка Томсен 1984 г.
- Кринолина Томсен 1976 г., а не Сметана 1982 г.
- Круциспина Эспеланд и Трондсен, 1986 г.
- Диафаноэка Эллис 1930 г.
- Дидимека Доуэлд, 2003 г.
- Какоека Бак и Марчант, 1991 г.
- Монокоста Томсен 1979 не Монокост Шуман 1904
- Нанноэка Томсен 1988 г.
- Парвикорбикула Дефландр 1960 г.
- Плеврасига Шиллер 1925
- Полифибула Мэнтон 1981
- Саэпикула Ледбитер 1980
- Сароэка Томсен 1979 г.
- Спиноэка Томсен, Остергаард и Хансен, 1995 г., но не Поульсен, 1973 г.
- Стефанаканта Томсен 1983
- Стефаноэка Эллис 1930 г.
- Синдетофиллум Томсен и Моэструп, 1983 г.
- Томсенелла Оздикмен 2009 г.
Геномы и транскриптомы
[ редактировать ]Monosiga brevicollis Геном
[ редактировать ]Геном Monosiga brevicollis содержит 41,6 миллиона пар оснований. [10] по размеру подобен нитчатым грибам и другим свободноживущим одноклеточным эукариотам, но намного меньше, чем у типичных животных. [10] В 2010 году филогеномное исследование показало, что в геноме Monosiga brevicollis присутствует несколько генов водорослей . Это могло быть связано с тем, что на ранней стадии эволюции хоанофлагелляты потребляли водоросли в качестве пищи посредством фагоцитоза . [37] Карр и др. (2010) [29] проверили геном M. brevicollis на наличие известных генов эукариотического мейоза. Из 19 известных протестированных эукариотических мейотических генов (в том числе 8, которые не участвуют ни в каком другом процессе, кроме мейоза), 18 были идентифицированы у M. brevicollis . Присутствие мейотических генов, включая гены, специфичные для мейоза, указывает на то, что мейоз и, как следствие, пол присутствуют внутри хоанофлагеллят.
сальпингоеки розетты Геном
[ редактировать ]Размер генома Salpingoeca Rosetta составляет 55 мегабаз. [38] В геноме присутствуют гомологи генов клеточной адгезии, нейропептидного и гликосфинголипидного метаболизма. S. Rosetta имеет половой жизненный цикл и переходы между гаплоидными и диплоидными стадиями . [28] В ответ на ограничение питательных веществ гаплоидные культуры S. Rosetta становятся диплоидными. Этот сдвиг плоидности совпадает с спариванием, во время которого маленькие жгутиковые клетки сливаются с более крупными жгутиковыми клетками. Есть также свидетельства исторического спаривания и рекомбинации у S. Rosetta .
S. Rosetta вызывает половое размножение морской бактерией Vibrio fischeri . [27] Единственный белок V. fischeri , EroS, полностью повторяет афродизиаковую активность живого V. fisheri .
Другие геномы
[ редактировать ]Одноклеточные амплифицированные геномы четырех некультивируемых морских хоанофлагеллят, предварительно названных UC1–UC4, были секвенированы в 2019 году. Геномы UC1 и UC4 относительно полны. [39]
Транскриптомы
[ редактировать ]Набор данных EST из Monosiga ovata был опубликован в 2006 году. [40] Основным открытием этого транскриптома стал домен Hoglet хоанофлагелляты, который пролил свет на роль перетасовки доменов в эволюции сигнального пути Hedgehog . M. ovata имеет как минимум четыре эукариотических мейотических гена. [29]
Транскриптом Stephanoeca diplocostata был опубликован в 2013 году. Это первый транскриптом лорикатной хоанофлагелляты. [31] привели к открытию переносчиков кремния в хоанофлагеллятах. Впоследствии подобные гены были идентифицированы у второго вида лоричатых, Diaphanoeca grandis . Анализ этих генов показал, что переносчики кремния хоанофлагеллят гомологичны переносчикам кремния типа SIT диатомовых водорослей и эволюционировали посредством горизонтального переноса генов .
Еще 19 транскриптомов были опубликованы в 2018 году. Было обнаружено большое количество семейств генов, которые ранее считались предназначенными только для животных. [41]
Галерея
[ редактировать ]
 Сальпингоека под PCM
Сальпингоека под PCM Салпингоека сп. раздел под ТЕМ
Салпингоека сп. раздел под ТЕМ Колония Desmarella moniliformis под PCM
Колония Desmarella moniliformis под PCM
 Колония Sphaeroeca (около 230 особей) под световой микроскопией.
Колония Sphaeroeca (около 230 особей) под световой микроскопией.






Ссылки
[ редактировать ]- ^ Фонсека, Каролина; Мендонса Фильо, Жоау Грасиано; Реолид, Матиас; Дуарте, Луис В.; де Оливейра, Антониу Доницети; Соуза, Жаклин Торрес; Лезен, Карин (23 января 2023 г.). «Первое предполагаемое появление в летописи окаменелостей хоанофлагеллят, сестринской группы Metazoa» . Научные отчеты . 13 (1): 1242. Бибкод : 2023NatSR..13.1242F . дои : 10.1038/s41598-022-26972-8 . ISSN 2045-2322 . ПМЦ 9870899 . ПМИД 36690681 .
- ^ Парфри Л.В., Лар DJ, Нолл А.Х., Кац Л.А. (август 2011 г.). «Оценка времени ранней диверсификации эукариот с помощью мультигенных молекулярных часов» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (33): 13624–9. Бибкод : 2011PNAS..10813624P . дои : 10.1073/pnas.1110633108 . ПМК 3158185 . ПМИД 21810989 .
- ^ Сэвилл-Кент, В. (1880). Руководство по Инфузории . Лондон, том. 1, с. 324, [1] .
- ^ Адл С.М., Басс Д., Лейн CE, Лукеш Дж., Шох К.Л., Смирнов А., Агата С., Берни С., Браун М.В., Бурки Ф., Карденас П., Чепичка И., Чистякова Л., дель Кампо Дж., Данторн М., Эдвардсен Б., Эглит. Ю, Гийу Л, Хампл В, Хейсс А.А., Хоппенрат М, Джеймс Тай, Карнковска А, Карпов С, Ким Е, Колиско М, Кудрявцев А, Лар DJG, Лара Е, Ле Галл Л, Линн Д.Х., Манн Д.Г., Массана Р. , Митчелл И.А.Д., Морроу С., Парк Дж.С., Павловски Дж.В., Пауэлл М.Дж., Рихтер Д.Дж., Рюкерт С., Шедвик Л., Шимано С., Шпигель Ф.В., Торруэлла Г., Юсеф Н., Златогурский В., Чжан К. (2019). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ПМК 6492006 . ПМИД 30257078 .
- ^ Кинг Н., Уэстбрук М.Дж., Янг С.Л., Куо А., Абедин М., Чепмен Дж. и др. (февраль 2008 г.). «Геном хоанофлагелляты Monosiga brevicollis и происхождение многоклеточных животных» . Природа . 451 (7180): 783–8. Бибкод : 2008Natur.451..783K . дои : 10.1038/nature06617 . ПМК 2562698 . ПМИД 18273011 .
- ^ Нитше Ф., Карр М., Арндт Х., Ледбитер Б.С. (2011). «Таксономия высокого уровня и молекулярная филогенетика Choanoflagellatea». Журнал эукариотической микробиологии . 58 (5): 452–62. дои : 10.1111/j.1550-7408.2011.00572.x . ПМИД 21895836 . S2CID 2076733 .
- ^ Кавалер-Смит Т. (1998). «Неомонада и происхождение животных и грибов». В Кумбс Г.Х., Викерман К., Сани М.А., Уоррен А. (ред.). Эволюционные взаимоотношения между простейшими . Лондон: Клювер. стр. 375–407.
- ^ Перейти обратно: а б Ледбитер Б.С. (2015). Хоанофлагелляты: эволюция, биология и экология . Университет Бирмингема. ISBN 978-0-521-88444-0 .
- ^ Перейти обратно: а б «Бактериальный белок действует как афродизиак для хоанофлагеллят» .
- ^ Перейти обратно: а б с д и ж г час я Кинг Н. , Уэстбрук М.Дж., Янг С.Л., Куо А., Абедин М., Чепмен Дж. и др. (февраль 2008 г.). «Геном хоанофлагелляты Monosiga brevicollis и происхождение многоклеточных животных» . Природа . 451 (7180): 783–8. Бибкод : 2008Natur.451..783K . дои : 10.1038/nature06617 . ПМК 2562698 . ПМИД 18273011 .
- ^ Перейти обратно: а б с д и Ледбитер Б.С., Томсен Х (2000). «Отряд Хоанофлагеллиды». Иллюстрированный справочник по простейшим, второе издание. Лоуренс: Общество протозоологов . 451 : 14–38.
- ^ Перейти обратно: а б Карпов С., Ледбитер Б.С. (май 1998 г.). «Строение и состав цитоскелета хоанофлагеллят». Журнал эукариотической микробиологии . 45 (3): 361–367. дои : 10.1111/j.1550-7408.1998.tb04550.x . S2CID 86287656 .
- ^ Перейти обратно: а б с Ледбитер Б.С., Келли М. (2001). «Эволюция хоанофлагеллят и губок животных». Вода и атмосфера онлайн . 9 (2): 9–11.
- ^ Перейти обратно: а б Ледбитер Б.С. (февраль 1983 г.). «История жизни и ультраструктура нового морского вида Proterospongia (Choanoflagellida)». Ж. Мар. Биол. доц. Великобритания 63 (1): 135–160. Бибкод : 1983JMBUK..63..135L . дои : 10.1017/S0025315400049857 . S2CID 84666673 .
- ^ Перейти обратно: а б Мендоса Л., Тейлор Дж.В., Аджелло Л. (2002). «Класс mesomycetozoea: гетерогенная группа микроорганизмов на границе животных и грибов». Ежегодный обзор микробиологии . 56 : 315–44. дои : 10.1146/annurev.micro.56.012302.160950 . ПМИД 12142489 . S2CID 14764188 .
- ^ Томсен Х (1982). Планктонные хоанофлагелляты из Диско Бугт, Западная Гренландия, с исследованием морского нанопланктона этого района . Meddelelser om Gronland, Биологические науки. Том. 8. стр. 3–63. ISBN 978-87-635-1149-0 .
- ^ Бак К.Р., Гаррисон Д.Л. (июнь 1988 г.). «Распределение и численность хоанофлагеллят (Acanthoecidae) в кромке льда в море Уэдделла, Антарктида». Мар Биол. 98 (2): 263–269. Бибкод : 1988MarBi..98..263B . дои : 10.1007/BF00391204 . S2CID 84931348 .
- ^ Томсен Х., Бак К., Чавес Ф. (1991). «Хоанофлагелляты вод центральной Калифорнии: таксономия, морфология и видовые комплексы». Офелия . 33 (2): 131–164. дои : 10.1080/00785326.1991.10429736 .
- ^ Перейти обратно: а б с д и Карр М., Ледбитер Б.С., Хасан Р., Нельсон М., Балдауф С.Л. (октябрь 2008 г.). «Молекулярная филогения хоанофлагеллят, сестринской группы Metazoa» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (43): 16641–6. Бибкод : 2008PNAS..10516641C . дои : 10.1073/pnas.0801667105 . ПМЦ 2575473 . ПМИД 18922774 .
- ^ «Недавно открытые микроорганизмы объединяются, «переворачиваются» » . HHMI.org . Проверено 29 октября 2019 г.
- ^ Баттерфилд, штат Нью-Джерси (1 апреля 1997 г.). «Экология планктона и протерозой-фанерозойский переход» . Палеобиология . 23 (2): 247–262. Бибкод : 1997Pbio...23..247B . дои : 10.1017/S009483730001681X . S2CID 140642074 .
- ^ Ву, Кэтрин Дж. (24 сентября 2020 г.). «Ничто не ест вирусы, верно? Познакомьтесь с некоторыми голодными протистами: новые генетические данные доказывают, что одноклеточные морские микробы могут питаться вирусами» . Нью-Йорк Таймс . п. D3 (29 сентября 2020 г., печатная ред.) . Проверено 1 декабря 2020 г.
- ^ Де Луф, Арнольд; Шуфс, Лилиан (2019). «Способ действия фарнезола, «благородного неизвестного», в частности, в гомеостазе Ca2+, и его ювенильных гормональных эфиров в эволюционной ретроспективе» . Границы в неврологии . 13 : 141. дои : 10.3389/fnins.2019.00141 . ПМК 6397838 . PMID 30858798 .
- ^ Клаус Нильсен. Эволюция животных: взаимоотношения живых типов. 3-е изд. Клаус Нильсен. Оксфорд, Великобритания: Издательство Оксфордского университета, 2012, с. 14.
- ^ Карр М., Ледбитер Б.С., Балдауф С.Л. (2002). «Консервативные мейотические гены указывают на пол хоанофлагеллят». Журнал эукариотической микробиологии . 57 (1): 56–62. дои : 10.1111/j.1550-7408.2009.00450.x . ПМИД 20015185 . S2CID 205759832 .
- ^ Ледбитер Б.С., Карпов С.А. (сентябрь – октябрь 2000 г.). «Образование кисты у пресноводного штамма хоанофлагелляты Desmarella moniliformis Kent». Журнал эукариотической микробиологии . 47 (5): 433–9. дои : 10.1111/j.1550-7408.2000.tb00071.x . ПМИД 11001139 . S2CID 23357186 .
- ^ Перейти обратно: а б Возница А., Гердт Дж.П., Хьюлетт Р.Э., Кларди Дж., Кинг Н. (сентябрь 2017 г.). «Спаривание у ближайших ныне живущих родственников животных индуцируется бактериальной хондроитиназой» . Клетка . 170 (6): 1175–1183.e11. дои : 10.1016/j.cell.2017.08.005 . ПМК 5599222 . ПМИД 28867285 .
- ^ Перейти обратно: а б Левин TC, King N (ноябрь 2013 г.). «Доказательства пола и рекомбинации у хоанофлагеллят Salpingoeca Rosetta» . Современная биология . 23 (21): 2176–80. Бибкод : 2013CBio...23.2176L . дои : 10.1016/j.cub.2013.08.061 . ПМК 3909816 . ПМИД 24139741 .
- ^ Перейти обратно: а б с Карр М., Ледбитер Б.С., Балдауф С.Л. (2010). «Консервативные мейотические гены указывают на пол хоанофлагеллят». Журнал эукариотической микробиологии . 57 (1): 56–62. дои : 10.1111/j.1550-7408.2009.00450.x . ПМИД 20015185 . S2CID 205759832 .
- ^ Ледбитер Б.С., Ю Кью, Кент Дж, Стекел DJ (январь 2009 г.). «Трёхмерные изображения хоанофлагеллят лорики» . Слушания. Биологические науки . 276 (1654): 3–11. дои : 10.1098/rspb.2008.0844 . ПМЦ 2581655 . ПМИД 18755674 .
- ^ Перейти обратно: а б Маррон А.О., Олстон М.Дж., Хевенс Д., Акам М., Каккамо М., Холланд П.В. , Уокер Дж. (апрель 2013 г.). «Семейство диатомоподобных переносчиков кремния в кремнистых лорикатных хоанофлагеллятах» . Слушания. Биологические науки . 280 (1756): 20122543. doi : 10.1098/rspb.2012.2543 . ПМЦ 3574361 . ПМИД 23407828 .
- ^ Кинг Н. , Кэрролл С.Б. (декабрь 2001 г.). «Рецепторная тирозинкиназа из хоанофлагеллят: молекулярное понимание ранней эволюции животных» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (26): 15032–7. Бибкод : 2001PNAS...9815032K . дои : 10.1073/pnas.261477698 . ПМК 64978 . ПМИД 11752452 .
- ^ Кантелл CE, Франзен О, Сенсенбо Т (октябрь 1982 г.). «Ультраструктура многореснитчатых воротниковых клеток личинки пилидия Lineus bilineatus (Nemertini)». Зооморфология . 101 (1): 1–15. дои : 10.1007/BF00312027 . S2CID 42242685 .
- ^ Торруэлла, Гифре; Мендоса, Алекс де; Грау-Бове, Ксавье; Анто, Меритчель; Чаплин, Марк А.; Кампо, Хавьер дель; Эме, Лаура; Перес-Кордон, Грегорио; Уиппс, Кристофер М. (2015). «Филогеномика раскрывает конвергентную эволюцию образа жизни у близких родственников животных и грибов» . Современная биология . 25 (18): 2404–2410. Бибкод : 2015CBio...25.2404T . дои : 10.1016/j.cub.2015.07.053 . ПМИД 26365255 .
- ^ Ревьерс, Б. де. (2006). Биология и филогения водорослей . Editora Artmed, Порту-Алегри, с. 156.
- ^ (Адл и др., 2005)
- ^ Сунь Г, Ян З, Ишвар А, Хуан Дж (декабрь 2010 г.). «Гены водорослей у ближайших родственников животных» . Молекулярная биология и эволюция . 27 (12): 2879–89. дои : 10.1093/molbev/msq175 . ПМИД 20627874 .
- ^ Фэйркло С.Р., Чен З., Крамер Э., Цзэн К., Янг С., Робертсон Х.М., Бегович Э., Рихтер Д.Д., Расс С., Уэстбрук М.Дж., Мэннинг Дж., Ланг Б.Ф., Хаас Б., Нусбаум С., Кинг Н. (февраль 2013 г.). «Эволюция генома преметазоа и регуляция дифференцировки клеток хоанофлагелляты Salpingoeca Rosetta» . Геномная биология . 14 (2): Р15. дои : 10.1186/gb-2013-14-2-r15 . ПМК 4054682 . ПМИД 23419129 .
- ^ Лопес-Эскардо, защитник; Грау-Бове, X; Гийоме-Адкинс, А; Гут, М; Серацкий, Мэн; Руис-Трилло, I (25 ноября 2019 г.). «Реконструкция эволюции белковых доменов с использованием одноклеточных амплифицированных геномов некультивируемых хоанофлагеллят проливает свет на происхождение животных» . Философские труды Лондонского королевского общества. Серия Б: Биологические науки . 374 : 20190088.doi : (1786 ) 10.1098/rstb.2019.0088 . ПМК 6792448 . ПМИД 31587642 .
- ^ Снелл Э.А., Брук Н.М., Тейлор В.Р., Казан Д., Филипп Х., Холланд П.В. (февраль 2006 г.). «Необычный белок хоанофлагеллят, выделяемый в результате автокаталитической обработки Hedgehog» . Слушания. Биологические науки . 273 (1585): 401–7. дои : 10.1098/rspb.2005.3263 . ПМК 1560198 . ПМИД 16615205 .
- ^ Рихтер, Дэниел Дж; Фозуни, Париназ; Эйзен, Майкл Б; Кинг, Николь (31 мая 2018 г.). «Инновации, сохранение и утрата семейства генов в линии ствола животных» . электронная жизнь . 7 : е34226. doi : 10.7554/eLife.34226 . ПМК 6040629 . ПМИД 29848444 .
Внешние ссылки
[ редактировать ]- ChoanoWiki - совместный ресурс, поддерживаемый исследовательским сообществом хоанофлагеллят.
- Веб-страница Древа жизни хоанофлагеллят
- Monosiga brevicollis Браузер генома
- Химия вашего мозга существовала до того, как появились животные
- Choanobase — генетическая библиотека хоанофлагеллят, разработанная и поддерживаемая лабораторией Николь Кинг в Калифорнийском университете в Беркли.
