Микроспоридии
| Микроспоридии | |
|---|---|
 | |
| Споробласт Фибрилланосема крангоничис | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | Аморфея |
| Клэйд : | Подчиняться |
| (без рейтинга): | Опистоконта |
| Клэйд : | Холомикота |
| Королевство: | Грибы |
| Subkingdom: | Розелломицета |
| Сорт: | Микроспоридиомикота Бенни 2007 |
| Занятия и заказы [1] | |
| Синонимы | |
Микроспоридии группа спорообразующих одноклеточных — паразитов . Эти споры содержат экструзионное устройство, имеющее спиральную полярную трубку, оканчивающуюся закрепляющим диском в апикальной части споры. [7] Когда-то их считали простейшими или протистами , но теперь известно, что это грибы . [8] или сестринская группа грибов. [9] Эти грибковые микробы являются облигатными эукариотическими паразитами, которые используют уникальный механизм заражения клеток-хозяев. [7] Недавно в ходе исследования Корнелла, проведенного в 2017 году, они были обнаружены в заражении жесткокрылых крупномасштабном . На данный момент около 1500 из, вероятно, более миллиона [10] виды названы. Микроспоридии встречаются только у животных- хозяев , и все основные группы животных являются носителями микроспоридий. Большинство заражают насекомые , но они также являются причиной распространенных заболеваний ракообразных и рыб . Названные виды микроспоридий обычно поражают один вид-хозяин или группу близкородственных таксонов. Примерно 10 процентов видов являются паразитами позвоночных животных — несколько видов, большинство из которых являются условно-патогенными, могут инфицировать человека, у которого они могут вызывать микроспоридиоз .
После заражения они по-разному влияют на своих хозяев, захватывая все органы и ткани, хотя, как правило, разными видами специализированных микроспоридий. Некоторые виды смертоносны, а некоторые используются для биологической борьбы с насекомыми-вредителями. Паразитарная кастрация , гигантизм или смена пола хозяина — все это потенциальные последствия микроспоридийного паразитизма (у насекомых). В наиболее запущенных случаях паразитизма микроспоридии полностью управляют клеткой-хозяином и контролируют ее метаболизм и размножение, образуя ксеному . [11]
Репликация происходит внутри клеток хозяина, которые заражаются одноклеточными спорами . Их размер варьируется от 1 до 40 мкм, что делает их одними из самых маленьких эукариотов . [ нужна ссылка ] Микроспоридии, поражающие млекопитающих, имеют размер 1,0–4,0 мкм. [12] Они также имеют самые маленькие эукариотические геномы .
Термины «микроспоридий» (мн. «микроспоридии») и «микроспоридии» используются как народные названия членов группы. Название Microsporidium Balbiani, 1884 г. [13] также используется как общий род для представителей incertae sedis . [14]

Морфология
[ редактировать ]У микроспоридий отсутствуют митохондрии , вместо них имеются митосомы . У них также отсутствуют подвижные структуры, такие как жгутики .
Микроспоридии образуют высокоустойчивые споры, способные выживать вне хозяина до нескольких лет. Морфология спор полезна для различения разных видов. Споры большинства видов имеют овальную или грушевидную форму, но споры палочковидной или сферической формы не являются чем-то необычным. Некоторые роды производят споры уникальной для этого рода формы.
Спора защищена стенкой, состоящей из трех слоев:
- внешняя электронно-плотная экзоспора
- срединная, широкая и на вид бесструктурная эндоспора , содержащая хитин.
- тонкая внутренняя плазматическая мембрана
В большинстве случаев имеется два тесно связанных ядра , образующих диплокарион , но иногда бывает только одно.
Передняя половина споры содержит гарпунообразный аппарат с длинной нитевидной полярной нитью , которая свернута в задней половине споры. Передняя часть полярной нити окружена поляропластом — пластинкой мембран. Позади полярной нити находится задняя вакуоль . [11]
Инфекция
[ редактировать ]В кишечнике хозяина спора прорастает; он создает осмотическое давление до тех пор, пока его жесткая стенка не разрывается в самом тонком месте на вершине. Задняя вакуоль набухает, заставляя полярную нить быстро выбрасывать инфекционное содержимое в цитоплазму потенциального хозяина. Одновременно материал нити перестраивается, образуя трубку, которая действует как игла для подкожных инъекций и проникает в эпителий кишечника.
Попав внутрь клетки-хозяина, спороплазма растет, делясь или образуя многоядерный плазмодий , прежде чем производить новые споры. Жизненный цикл значительно варьируется. Некоторые имеют простой бесполый жизненный цикл, [16] в то время как другие имеют сложный жизненный цикл, включающий несколько хозяев и как бесполое, так и половое размножение . Различные типы спор могут образовываться на разных стадиях, вероятно, с разными функциями, включая аутоинфекцию (передачу внутри одного хозяина).
Медицинские последствия
[ редактировать ]У животных и людей микроспоридии часто вызывают хронические, изнурительные заболевания, а не смертельные инфекции. Воздействие на хозяина включает снижение продолжительности жизни, фертильности, веса и общей работоспособности. о вертикальной передаче Часто сообщается микроспоридий.
В случае насекомых-хозяев вертикальная передача часто происходит в виде трансовариальной передачи, при которой микроспоридийные паразиты переходят из яичников самки-хозяина в яйца и в конечном итоге размножаются в инфицированных личинках. Амблиоспора Salinaria n. сп. Culex Amblyospora , Californica поражающая комара tarsalis Coquillett. типичными примерами трансовариальной передачи микроспоридий являются [17] [18] [19] [20] Микроспоридии, особенно комары Vavraia culicis , исследуются как возможный «эволюционно-устойчивый» метод борьбы с малярией. [21] Микроспоридийная инфекция Anopheles gambiae (основной переносчик малярии Plasmodium falciparum ) снижает малярийную инфекцию комаров и сокращает продолжительность их жизни. [22] Поскольку большинство инфицированных малярией комаров естественным образом умирают до того, как малярийный паразит становится достаточно зрелым для передачи, любое увеличение смертности комаров из-за микроспоридийной инфекции может снизить передачу малярии человеку. В мае 2020 года исследователи сообщили, что Microsporidia MB , симбионт средней кишки и яичников An. arabiensis , значительно ухудшал передачу P. falciparum , не оказывал «явного влияния» на приспособленность комаров-хозяев и передавался вертикально (по наследству). [23]
Клинический
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( ноябрь 2013 г. ) |
Микроспоридийные инфекции человека иногда вызывают заболевание, называемое микроспоридиозом . По меньшей мере 14 видов микроспоридий, относящихся к восьми родам, признаны патогенами человека . К ним относятся Trachipleistophora hominis . [24]
Как гиперпаразиты
[ редактировать ]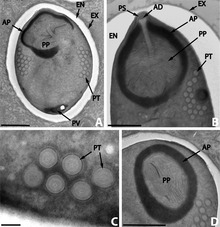
Микроспоридии могут инфицировать различных хозяев, включая хозяев, которые сами являются паразитами. В этом случае вид микроспоридий является гиперпаразитом , то есть паразитом паразита. Например, известно более восемнадцати видов, паразитирующих на дигенеях (паразитических плоских червях ). Эти дигенеи сами являются паразитами различных позвоночных и моллюсков . Восемь из этих видов относятся к роду Nosema . [25] Точно так же вид микроспоридий Toguebayea baccigeri является паразитом дигенеи, фаустулида Bacciger israelensis , который сам является кишечным паразитом морской рыбы, болотца Boops Boops (Teleostei, Sparidae). [26]
Геномы
[ редактировать ]Микроспоридии имеют наименьший из известных (ядерных) эукариотических геномов . Паразитический образ жизни микроспоридий привел к потере многих митохондриальных генов и генов Гольджи , и даже их рибосомальные РНК уменьшаются в размерах по сравнению с таковыми у большинства эукариот . Как следствие, геномы микроспоридий намного меньше, чем у других эукариот. Известные в настоящее время геномы микроспоридий имеют размер от 2,5 до 11,6 Мб и кодируют от 1848 до 3266 белков, что находится в том же диапазоне, что и у многих бактерий . [27]
Горизонтальный перенос генов (HGT), по-видимому, неоднократно происходил в микроспоридиях. Например, геномы Encephalitozoon romaleae и Trachipleistophora hominis содержат гены, происходящие от животных и бактерий, а некоторые даже от грибов. [27]
восстановление ДНК
[ редактировать ]Белковый комплекс Rad9 -Hus1 (также известный как комплекс 9-1-1) у - Rad1 эукариот рекрутируется в места повреждения ДНК , где считается, что он помогает запустить каскад сигнализации контрольных точек. Гены, кодирующие гетеротример 9-1-1, присутствуют в микроспоридиях. [28] Помимо комплекса 9-1-1, репарации ДНК , что указывает на то, что репарация повреждений ДНК, вероятно, происходит в микроспоридиях. присутствуют и другие компоненты механизма [28]
Филогения
[ редактировать ]Филогения Rozellomycota [29] [30]
| Розелломицета |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Классификация
[ редактировать ]Первый описанный род микроспоридий, Nosema , первоначально был отнесен Нэгели к группе грибов Schizomycetes вместе с некоторыми бактериями и дрожжами . [31] [32] Некоторое время микроспоридии считались очень примитивными эукариотами, отнесенными к группе простейших Cnidospora . [5] Позже, особенно из-за отсутствия митохондрий, их поместили вместе с другими простейшими, такими как дипломонады , парабазалиды и архамебы, в простейших группу Archezoa . [33] Более поздние исследования фальсифицировали эту теорию раннего происхождения (всех этих видов). Вместо этого предполагается, что микроспоридии являются высокоразвитыми и специализированными организмами, которые просто выполняют функции, которые больше не нужны, поскольку они предоставляются хозяином. [34] Более того, спорообразующие организмы вообще имеют сложную систему размножения, как полового, так и бесполого, которая выглядит далеко не примитивной.
С середины 2000-х годов микроспоридии относят к грибам или к сестринской группе грибов с общим предком. [35] [36] [37] [38]
Работа по идентификации клад во многом зависит от среды обитания и хозяина. Воссбринк и Дебруннер-Фоссбринк предложили три класса микроспоридий в зависимости от среды обитания: Aquasporidia, Marinosporidia и Terresporidia. [39]
Вторая классификация Кавальер-Смита, 1993 г.: [40]
- Подтип Rudimicrospora Кавальер-Смит 1993 г.
- Класс Миниспорея Кавальер-Смит 1993 г.
- Отряд Minisporida Sprague, 1972 г.
- Класс Мечниковелла Вайзер, 1977 г.
- Орден Мечниковеллида Вивье, 1975 г.
- Класс Миниспорея Кавальер-Смит 1993 г.
- Подтип Polaroplasta Кавальер-Смит 1993 г.
- Класс Pleistophoridea Кавальер-Смит 1993 г.
- Отряд Pleistophorida Stempell 1906 г.
- Класс Диспорея Кавальер-Смит 1993 г.
- Подкласс Unikaryotia Cavalier-Smith 1993 г.
- Подкласс Diplokaryotia Cavalier-Smith 1993 г.
- Класс Pleistophoridea Кавальер-Смит 1993 г.
|
См. также
[ редактировать ]- Список родов микроспоридий
- Глюгея — род микроспоридий.
- Nosema apis — микроспоридийный паразит пчел.
Ссылки
[ редактировать ]- ^ Виджаявардене, штат Нью-Йорк; Хайд, К.Д.; Дай, ДК; Санчес-Гарсия, М.; Гото, БТ; Саксена, РК; и др. (2022). «Очерк грибов и грибоподобных таксонов – 2021» . Микосфера . 13 (1): 53–453. дои : 10.5943/микосфера/13/1/2 . hdl : 10481/76378 . S2CID 249054641 .
- ^ Бальбиани, Г (1882). «О микроспоридиях или псороспермии членистоногих» . ЧР акад. Наука . 95 :1168–71.
- ^ Делфи, Дж. 1936. Подцарство простейших. В: Перье, Р. (ред.). Фауна Франции в иллюстрированных синоптических таблицах , т. 1А. Делагрейв: Париж.
- ^ Левин, Северная Дакота; и др. (1980). «Недавно пересмотренная классификация простейших» . Журнал протозоологии . 27 (1): 37–58. дои : 10.1111/j.1550-7408.1980.tb04228.x . ПМИД 6989987 .
- ^ Перейти обратно: а б Корлисс Дж.О., Левин Н.Д. (1963). «Создание Microsporidea как нового класса в подтипе простейших Cnidospora». Журнал протозоологии . 10 (Приложение): 26–27. doi : 10.1111/jeu.1963.10.issue-s3 .
- ^ Спрэг, В. (1977). Классификация и филогения микроспоридий. В: Сравнительная патобиология . том. 2. Систематика микроспоридий. Ли А. Булла и Томас К. Ченг (ред.). стр. 1–30. Нью-Йорк: Пленум Пресс, [1] .
- ^ Перейти обратно: а б Франзен, К. (2005). Как микроспоридии проникают в клетки? Folia Parasitologica, 52 (1–2), 36–40. doi.org/10.14411/fp.2005.005
- ^ Хиббетт, Д.С.; и др. (2007). «Филогенетическая классификация грибов более высокого уровня» (PDF) . Микологические исследования . 111 (5): 509–47. дои : 10.1016/j.mycres.2007.03.004 . ПМИД 17572334 . S2CID 4686378 .
- ^ Силар, Филипп (2016). Эукариотические протисты: происхождение, эволюция и биология эукариотических микробов . ХЭЛ. п. 462. ИСБН 978-2-9555841-0-1 .
- ^ Хоксворт, Дэвид (2001). «Масштабы грибкового разнообразия: пересмотренная оценка в 1,5 миллиона специй». Микологические исследования . 105 (12): 1422. doi : 10.1017/S0953756201004725 .
- ^ Перейти обратно: а б Ронни Ларссон, Лундский университет (факультет биологии клеток и организмов). Цитология и таксономия микроспоридий. Архивировано 12 сентября 2009 г. в Wayback Machine , 2004 г.
- ^ Дидье, Э.С. (апрель 2005 г.). «Микроспоридиоз: новая оппортунистическая инфекция у людей и животных». Акта Троп . 94 (1): 61–76. doi : 10.1016/j.actatropica.2005.01.010 . ПМИД 15777637 .
- ^ Бальбиани, Г. 1884. Псороспермия суставных или микроспоридий, стр. 150–168, 184. В кн.: Уроки споровиков . Париж: Дуэн, [2] .
- ^ Хоффман, Г. (1999). Паразиты пресноводных рыб Северной Америки , 2-е изд., Калифорнийский университет Press, Беркли, Калифорния, США, с. 89, [3] .
- ^ Уинтерс, AD; Фейсал, М. (2014). «Молекулярная и ультраструктурная характеристика Dictyocoela diporeiae n. sp. (Microsporidia), паразита видов Diporeia (Amphipoda, Gammaridea)» . Паразит . 21 : 26. doi : 10.1051/parasite/2014028 . ПМК 4059264 . ПМИД 24934702 .
- ^ Айронсайд Дж.Э. (2007). «Многократная потеря пола в пределах одного рода Microsporidia» . Эволюционная биология BMC . 7:48 . дои : 10.1186/1471-2148-7-48 . ПМК 1853083 . ПМИД 17394631 .
- ^ Андреадис Т.Г., Холл Д.В. (август 1979 г.). «Развитие, ультраструктура и способ передачи Amblyospora sp. (Microspora) у комаров». Журнал протозоологии . 26 (3): 444–52. дои : 10.1111/j.1550-7408.1979.tb04651.x . ПМИД 536933 .
- ^ Андреадис Т.Г., Холл Д.В. (сентябрь 1979 г.). «Значение трансовариальных инфекций Amblyospora sp. (Microspora:Thelohaniidae) в отношении поддержания паразитов у комаров Culex salinarius». Журнал патологии беспозвоночных . 34 (2): 152–7. дои : 10.1016/0022-2011(79)90095-8 . ПМИД 536610 .
- ^ Ян Г.К., Холл Д.В., Зам С.Г. (1986). «Сравнение жизненных циклов двух амблиоспор (Microspora: Amblyosporidae) у комаров Culex salinarius и Culex tarsalis Coquillett». Журнал Флоридской ассоциации борьбы с комарами . 57 (1): 24–27.
- ^ Бекнель Дж. Дж., Андреадис Т. Г. (май 1998 г.). « Amblyospora salinaria n. sp. (Microsporidia: amblyosporidae), паразит Culex salinarius (Diptera: culicidae): этапы жизненного цикла у промежуточного хозяина» . Журнал патологии беспозвоночных . 71 (3): 258–62. дои : 10.1006/jipa.1998.4729 . ПМИД 9538031 .
- ^ Коэлла, Джейкоб С.; Лоренц, Лена; Баргеловский, Ирка (2009). Глава 12. Микроспоридии как устойчивые к эволюции агенты борьбы с малярией? . Достижения паразитологии. Том. 68. стр. 315–327. дои : 10.1016/S0065-308X(08)00612-X . ISBN 978-0-12-374787-7 . ПМИД 19289199 .
- ^ Баргеловски I, Коэлла Дж. К. (2009). Бейлис М. (ред.). «Возможный механизм подавления развития Plasmodium berghei у комара Anopheles gambiae с помощью микроспоридий Vavraia culicis» . ПЛОС ОДИН . 4 (3): e4676. Бибкод : 2009PLoSO...4.4676B . дои : 10.1371/journal.pone.0004676 . ПМЦ 2651578 . ПМИД 19277119 .
- ^ Херрен, Дж. К.; Мбаиси, Л; Мараро, Э; и др. (2020). «Микроспоридии ухудшают передачу Plasmodium falciparum у Anopheles arabiensis комаров » . Природные коммуникации . 11 (2187): 2187. Бибкод : 2020NatCo..11.2187H . дои : 10.1038/s41467-020-16121-y . ПМК 7198529 . ПМИД 32366903 .
- ^ Хайнц, Э; Уильямс, штат Калифорния; Накджанг, С; и др. (октябрь 2012 г.). «Геном облигатного внутриклеточного паразита Trachipleistophora hominis : новый взгляд на динамику генома микроспоридий и редуктивную эволюцию» . ПЛОС Патог . 8 (10): e1002979. дои : 10.1371/journal.ppat.1002979 . ПМК 3486916 . ПМИД 23133373 .
- ^ Перейти обратно: а б Тогебай, бакалавр наук; Киличини, Ю.; Диань, премьер-министр; Маршан, Б. (2014). «Ультраструктура и развитие Nosema podocotyloidis n. sp. (Microsporidia), гиперпаразита Podocotyloides magnatestis (Trematoda), паразита Parapristipoma Octolineatum (Teleostei)» . Паразит . 21 : 44. doi : 10.1051/parasite/2014044 . ПМК 4150386 . ПМИД 25174849 .

- ^ Микель, Хорди; Касем, Хичем; Баз-Гонсалес, Эдгар; Форонда, Пилар; Маршан, Бернар (2022). «Ультраструктурное и молекулярное исследование микроспоридий Toguebayea baccigeri n. gen., n. sp., гиперпаразита дигеновой трематоды Bacciger israelensis (Faustulidae), паразита Boops Boops (Teleostei, Sparidae)» . Паразит . 29 . EDP Sciences: 2. doi : 10.1051/parasite/2022007 . ISSN 1776-1042 . ПМЦ 8805611 . ПМИД 35103588 . S2CID 246443154 .
- ^ Перейти обратно: а б Корради, Н.; Селман, М. (2013). «Последние достижения в исследовании генома микроспоридий». Журнал эукариотической микробиологии . 60 (3): 309–312. дои : 10.1111/jeu.12030 . ПМИД 23445243 . S2CID 24504235 .
- ^ Перейти обратно: а б Маскареньяш Дос Сантос, Джулиан А.Т., Помберт Ж.Ф. (10 апреля 2022 г.). «Зажим для восстановления ДНК Rad9-Rad1-Hus1 обнаружен в микроспоридиях» . Геномная биология и эволюция . 14 (4): evac053. дои : 10.1093/gbe/evac053 . ПМЦ 9053307 . ПМИД 35439302 .
- ^ Перейти обратно: а б Виджаявардене Н.Н., Хайд К.Д., Аль-Ани Л.К., Тедерсу Л., Хэлеуотерс Д., Раджешкумар К.К. и др. (2020). «Описание грибов и грибоподобных таксонов» (PDF) . Микосфера . 11 (1): 1060–1456. дои : 10.5943/микосфера/11/1/8 . ISSN 2077-7019 .
- ^ Бойко, Джейми; Рейнке, Аарон В.; Стентифорд, Грант Д.; Уильямс, Бриони; Роджерс, Мартин С.Дж.; Басс, Дэвид (2022). «Микроспоридии: новый таксономический, эволюционный и экологический синтез» . Тенденции в паразитологии . 38 (8): 642–659. дои : 10.1016/j.pt.2022.05.007 .
- ^ Нэгели, К. фон (1857). «О новом заболевании тутового шелкопряда и родственных ему организмов. С. 760–61. В: Каспари Р. (ред.). Отчет о работе 33-го совещания немецких естествоиспытателей и врачей, состоявшегося в Бонне 18 сентября. до 24 1857 года» . Ботаническая газета . 15 :749-776.
- ^ Килинг, П.Дж.; Фаст, Нью-Мексико (2002). «Микроспоридии: биология и эволюция сильно уменьшенных внутриклеточных паразитов» (PDF) . Ежегодный обзор микробиологии . 56 (1): 93–116. дои : 10.1146/annurev.micro.56.012302.160854 . ПМИД 12142484 .
- ^ Кавальер-Смит, Т. (1993). «Царство простейших и его 18 типов» . Микробиологические обзоры . 57 (4): 953–994. дои : 10.1128/MR.57.4.953-994.1993 . ПМК 372943 . ПМИД 8302218 .
- ^ Килинг П.Дж., Сламовиц CH (декабрь 2004 г.). «Простота и сложность геномов микроспоридий» . Эукариотическая клетка . 3 (6): 1363–9. doi : 10.1128/EC.3.6.1363-1369.2004 . ПМК 539024 . ПМИД 15590811 .
- ^ Фишер В.М., Палмер Дж.Д. (сентябрь 2005 г.). «Доказательства грибкового происхождения микроспоридий на основе последовательностей рибосомальных РНК малых субъединиц». Молекулярная филогенетика и эволюция . 36 (3): 606–22. дои : 10.1016/j.ympev.2005.03.031 . ПМИД 15923129 .
- ^ Лю Ю.Дж., Ходсон MC, Холл BD (2006). «Потеря жгутика произошла только один раз в грибной линии: филогенетическая структура царства грибов выведена из генов субъединицы РНК-полимеразы II» . Эволюционная биология BMC . 6:74 . дои : 10.1186/1471-2148-6-74 . ПМЦ 1599754 . ПМИД 17010206 .
- ^ Гилл Э.Э., Фаст, Нью-Мексико (июнь 2006 г.). «Оценка взаимоотношений микроспоридий и грибов: комбинированный филогенетический анализ восьми генов». Джин . 375 : 103–9. дои : 10.1016/j.gene.2006.02.023 . ПМИД 16626896 .
- ^ Ли С.К., Корради Н., Бирнс Э.Дж. и др. (ноябрь 2008 г.). «Микроспоридии произошли от предков половых грибов» . Современная биология . 18 (21): 1675–9. дои : 10.1016/j.cub.2008.09.030 . ПМК 2654606 . ПМИД 18976912 .
- ^ Воссбринк CR, Дебраннер-Фосбринк BA (май 2005 г.). «Молекулярная филогения микроспоридий: экологические, ультраструктурные и таксономические соображения» . Фолиа Паразитологическая . 52 (1–2): 131–42, обсуждение 130. doi : 10.14411/fp.2005.017 . ПМИД 16004372 .
- ^ Кавалер-Смит (1993). «Царство простейших и его 18 типов» . Микробиологические обзоры . 57 (4): 953–94. дои : 10.1128/MR.57.4.953-994.1993 . ПМК 372943 . ПМИД 8302218 .
- ^ Алимов А.Ф., изд. (май 2007 г.). Протиста 2: Справочник по зоологии . Наука. п. 1141. ИСБН 9785020262249 .
- ^ Виджаявардене, штат Нью-Йорк; Хайд, К.Д.; Дай, ДК; Санчес-Гарсия, М.; Гото, БТ; Саксена, РК; и др. (2022). «Очерк грибов и грибоподобных таксонов – 2021» . Микосфера . 13 (1): 53–453. дои : 10.5943/микосфера/13/1/2 . hdl : 1854/LU-8754813 . S2CID 249054641 .
Внешние ссылки
[ редактировать ] Данные, относящиеся к микроспоридиям , в Wikispecies
Данные, относящиеся к микроспоридиям , в Wikispecies - Ресурсный центр биоинформатики BioHealthBase База данных последовательностей микроспордии и сопутствующая информация.
- Микроспоридии в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
| Базы данных органов управления : Национальные |
|---|