Паразитизм

Паразитизм - это тесная связь между видами , где один организм, паразит, живет на или внутри другого организма, хозяин , причиняет ему некоторый вред, и адаптируется структурно к этому образу жизни. [ 1 ] Энтомолог Эо Уилсон охарактеризовал паразитов как «хищников, которые едят добычу в единицах менее одного». [ 2 ] Паразиты включают в себя одноклеточные простейшие, такие как агенты малярии , спящая болезнь и амебическая дизентерия ; Животные, такие как анкилостомы , вши , комары и летучие мыши вампиров ; грибы, такие как медовый гриб и агенты стригущего лишания ; и такие растения, как омела , Доддер и Брумрап .
Существует шесть основных паразитических стратегий эксплуатации хозяев животных, а именно паразитарная кастрация , непосредственно переданный паразитизм (путем контакта), паразитизм с трофически передаваемым (путем еды), переносимый вектором паразитизм, паразитоизм и микропеключение. Одна основная ось классификации касается инвазивности: эндопаразит живет внутри тела хозяина; Эктопаразит живет снаружи, на поверхности хозяина.
Как и хищничество, паразитизм является типом взаимодействия потребителей и ресурсов , [ 3 ] Но в отличие от хищников , паразиты, за исключением паразитоидов, как правило, намного меньше, чем их хозяева, не убивают их и часто живут в их хозяевах или на их хозяевах. Паразиты животных высокоспециализированы и размножаются более высокой скоростью, чем их хозяева. Классические примеры включают взаимодействие между хозяевами позвоночных и ленточными червями , Flukes , вызывающим малярию виды Plasmodium и блохи .
Паразиты уменьшают физическую форму хозяина по общей или специализированной патологии , от паразитной кастрации до модификации поведения хозяина. Паразиты увеличивают свою собственную пригодность, используя хозяев для ресурсов, необходимых для их выживания, в частности, питая их и используя промежуточных (вторичных) хозяев, чтобы помочь в их передаче от одного окончательного (первичного) хоста к другому. Хотя паразитизм часто однозначный, он является частью спектра взаимодействия между видами , градификация посредством паразитоидзма в хищничество, путем эволюции в взаимность и в некоторых грибах, задерживаясь в сапрофитическом .
Люди знают о паразитах, таких как круглые черви и ленточные черви, со времен Древнего Египта , Греции и Рима . В ранние современные времена Энтони Ван Леувенхук наблюдала за Джардией Ламблией в своем микроскопе в 1681 году, в то время как Франческо Реди описал внутренние и внешние паразиты, включая случайность печени овец и клещей . Современная паразитология, разработанная в 19 веке. В человеческой культуре паразитизм имеет негативные коннотации. Они были эксплуатированы для сатирического эффекта в стихотворении Джонатана Свифта 1733 года «о поэзии: рапсодия», сравнивая поэты с гиперпаразитрическими «паразитами». В художественной литературе « Брэм Стокер » 1897 года ужасов готический роман «Дракула» и его многочисленные адаптации показали паразит с пьющим кровь . Фильм Ридли Скотта 1979 года «Чужой» был одним из многих произведений научной фантастики, в которых изображены паразитические инопланетные виды. [ 4 ]
Этимология
[ редактировать ]году, слово «паразит» происходит от средневекового французского паразита , от латинизированного паразита Впервые используется на английском языке в 1539 из древнегреческого παράσιτος [ 5 ] (Паразито) «Тот, кто ест за столом другого» по очереди из παρά [ 6 ] (Пара) «кроме, по 'и σῖτος (sitos) ' пшеница, еда». [ 7 ] Связанный термин паразитизм появляется на английском языке с 1611 года. [ 8 ]
Эволюционные стратегии
[ редактировать ]
Основные понятия
[ редактировать ]
Паразитизм-это своего рода симбиоз , близкое и постоянное долгосрочное биологическое взаимодействие между паразитом и его хозяином. В отличие от сапротрофов , паразиты питаются живыми хозяевами, хотя, например, некоторые паразитические грибы могут продолжать питаться хозяевами, которых они убили. В отличие от комменсализма и взаимного инфуализации , паразитические отношения вредят хозяину, либо питаются им, либо, как в случае кишечных паразитов, потребляя часть своей пищи. Поскольку паразиты взаимодействуют с другими видами, они могут легко действовать как векторы патогенов, вызывая заболевание . [ 9 ] [ 10 ] [ 11 ] Хищничество по определению не симбиоз, так как взаимодействие является кратким, но энтомолог Эо Уилсон охарактеризовал паразитов как «хищников, которые едят добычу в единицах менее одного». [2]
Within that scope are many possible strategies. Taxonomists classify parasites in a variety of overlapping schemes, based on their interactions with their hosts and on their life cycles, which are sometimes very complex. An obligate parasite depends completely on the host to complete its life cycle, while a facultative parasite does not. Parasite life cycles involving only one host are called "direct"; those with a definitive host (where the parasite reproduces sexually) and at least one intermediate host are called "indirect".[12][13] An endoparasite lives inside the host's body; an ectoparasite lives outside, on the host's surface.[14] Mesoparasites—like some copepods, for example—enter an opening in the host's body and remain partly embedded there.[15] Some parasites can be generalists, feeding on a wide range of hosts, but many parasites, and the majority of protozoans and helminths that parasitise animals, are specialists and extremely host-specific.[14] An early basic, functional division of parasites distinguished microparasites and macroparasites. These each had a mathematical model assigned in order to analyse the population movements of the host–parasite groupings.[16] The microorganisms and viruses that can reproduce and complete their life cycle within the host are known as microparasites. Macroparasites are the multicellular organisms that reproduce and complete their life cycle outside of the host or on the host's body.[16][17]
Much of the thinking on types of parasitism has focused on terrestrial animal parasites of animals, such as helminths. Those in other environments and with other hosts often have analogous strategies. For example, the snubnosed eel is probably a facultative endoparasite (i.e., it is semiparasitic) that opportunistically burrows into and eats sick and dying fish.[18] Plant-eating insects such as scale insects, aphids, and caterpillars closely resemble ectoparasites, attacking much larger plants; they serve as vectors of bacteria, fungi and viruses which cause plant diseases. As female scale insects cannot move, they are obligate parasites, permanently attached to their hosts.[16]
The sensory inputs that a parasite employs to identify and approach a potential host are known as "host cues". Such cues can include, for example, vibration,[19] exhaled carbon dioxide, skin odours, visual and heat signatures, and moisture.[20] Parasitic plants can use, for example, light, host physiochemistry, and volatiles to recognize potential hosts.[21]
Major strategies
[edit]There are six major parasitic strategies, namely parasitic castration; directly transmitted parasitism; trophically-transmitted parasitism; vector-transmitted parasitism; parasitoidism; and micropredation. These apply to parasites whose hosts are plants as well as animals.[16][22] These strategies represent adaptive peaks; intermediate strategies are possible, but organisms in many different groups have consistently converged on these six, which are evolutionarily stable.[22]
A perspective on the evolutionary options can be gained by considering four key questions: the effect on the fitness of a parasite's hosts; the number of hosts they have per life stage; whether the host is prevented from reproducing; and whether the effect depends on intensity (number of parasites per host). From this analysis, the major evolutionary strategies of parasitism emerge, alongside predation.[23]
| Host fitness | Single host, stays alive | Single host, dies | Multiple hosts |
|---|---|---|---|
| Able to reproduce (fitness > 0) |
Conventional parasite Pathogen |
Trophically-transmitted parasite[a] Trophically-transmitted pathogen |
Micropredator Micropredator |
| Unable to reproduce (fitness = 0) |
----- Parasitic castrator |
Trophically-transmitted parasitic castrator Parasitoid |
Social predator[b] Solitary predator |
Parasitic castrators
[edit]
Parasitic castrators partly or completely destroy their host's ability to reproduce, diverting the energy that would have gone into reproduction into host and parasite growth, sometimes causing gigantism in the host. The host's other systems remain intact, allowing it to survive and to sustain the parasite.[22][24] Parasitic crustaceans such as those in the specialised barnacle genus Sacculina specifically cause damage to the gonads of their many species[25] of host crabs. In the case of Sacculina, the testes of over two-thirds of their crab hosts degenerate sufficiently for these male crabs to develop female secondary sex characteristics such as broader abdomens, smaller claws and egg-grasping appendages. Various species of helminth castrate their hosts (such as insects and snails). This may happen directly, whether mechanically by feeding on their gonads, or by secreting a chemical that destroys reproductive cells; or indirectly, whether by secreting a hormone or by diverting nutrients. For example, the trematode Zoogonus lasius, whose sporocysts lack mouths, castrates the intertidal marine snail Tritia obsoleta chemically, developing in its gonad and killing its reproductive cells.[24][26]
Directly transmitted
[edit]
Directly transmitted parasites, not requiring a vector to reach their hosts, include such parasites of terrestrial vertebrates as lice and mites; marine parasites such as copepods and cyamid amphipods; monogeneans; and many species of nematodes, fungi, protozoans, bacteria, and viruses. Whether endoparasites or ectoparasites, each has a single host-species. Within that species, most individuals are free or almost free of parasites, while a minority carry a large number of parasites; this is known as an aggregated distribution.[22]
Trophically transmitted
[edit]
Trophically-transmitted parasites are transmitted by being eaten by a host. They include trematodes (all except schistosomes), cestodes, acanthocephalans, pentastomids, many roundworms, and many protozoa such as Toxoplasma.[22] They have complex life cycles involving hosts of two or more species. In their juvenile stages they infect and often encyst in the intermediate host. When the intermediate-host animal is eaten by a predator, the definitive host, the parasite survives the digestion process and matures into an adult; some live as intestinal parasites. Many trophically transmitted parasites modify the behaviour of their intermediate hosts, increasing their chances of being eaten by a predator. As with directly transmitted parasites, the distribution of trophically transmitted parasites among host individuals is aggregated.[22] Coinfection by multiple parasites is common.[27] Autoinfection, where (by exception) the whole of the parasite's life cycle takes place in a single primary host, can sometimes occur in helminths such as Strongyloides stercoralis.[28]
Vector-transmitted
[edit]
Vector-transmitted parasites rely on a third party, an intermediate host, where the parasite does not reproduce sexually,[14] to carry them from one definitive host to another.[22] These parasites are microorganisms, namely protozoa, bacteria, or viruses, often intracellular pathogens (disease-causers).[22] Their vectors are mostly hematophagic arthropods such as fleas, lice, ticks, and mosquitoes.[22][29] For example, the deer tick Ixodes scapularis acts as a vector for diseases including Lyme disease, babesiosis, and anaplasmosis.[30] Protozoan endoparasites, such as the malarial parasites in the genus Plasmodium and sleeping-sickness parasites in the genus Trypanosoma, have infective stages in the host's blood which are transported to new hosts by biting insects.[31]
Parasitoids
[edit]Parasitoids are insects which sooner or later kill their hosts, placing their relationship close to predation.[32] Most parasitoids are parasitoid wasps or other hymenopterans; others include dipterans such as phorid flies. They can be divided into two groups, idiobionts and koinobionts, differing in their treatment of their hosts.[33]
Idiobiont parasitoids sting their often-large prey on capture, either killing them outright or paralysing them immediately. The immobilised prey is then carried to a nest, sometimes alongside other prey if it is not large enough to support a parasitoid throughout its development. An egg is laid on top of the prey and the nest is then sealed. The parasitoid develops rapidly through its larval and pupal stages, feeding on the provisions left for it.[33]
Koinobiont parasitoids, which include flies as well as wasps, lay their eggs inside young hosts, usually larvae. These are allowed to go on growing, so the host and parasitoid develop together for an extended period, ending when the parasitoids emerge as adults, leaving the prey dead, eaten from inside. Some koinobionts regulate their host's development, for example preventing it from pupating or making it moult whenever the parasitoid is ready to moult. They may do this by producing hormones that mimic the host's moulting hormones (ecdysteroids), or by regulating the host's endocrine system.[33]
-
![Idiobiont parasitoid wasps immediately paralyse their hosts for their larvae (Pimplinae, pictured) to eat.[22]](//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Live_Tetragnatha_montana_%28RMNH.ARA.14127%29_parasitized_by_Acrodactyla_quadrisculpta_larva_%28RMNH.INS.593867%29_-_BDJ.1.e992.jpg/230px-Live_Tetragnatha_montana_%28RMNH.ARA.14127%29_parasitized_by_Acrodactyla_quadrisculpta_larva_%28RMNH.INS.593867%29_-_BDJ.1.e992.jpg) Idiobiont parasitoid wasps immediately paralyse their hosts for their larvae (Pimplinae, pictured) to eat.[22]
Idiobiont parasitoid wasps immediately paralyse their hosts for their larvae (Pimplinae, pictured) to eat.[22] -
 Koinobiont parasitoid wasps like this braconid lay their eggs via an ovipositor inside their hosts, which continue to grow and moult.
Koinobiont parasitoid wasps like this braconid lay their eggs via an ovipositor inside their hosts, which continue to grow and moult. -
 Phorid fly (centre left) is laying eggs in the abdomen of a worker honey-bee, altering its behaviour.
Phorid fly (centre left) is laying eggs in the abdomen of a worker honey-bee, altering its behaviour.
![Idiobiont parasitoid wasps immediately paralyse their hosts for their larvae (Pimplinae, pictured) to eat.[22]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/65/Live_Tetragnatha_montana_%28RMNH.ARA.14127%29_parasitized_by_Acrodactyla_quadrisculpta_larva_%28RMNH.INS.593867%29_-_BDJ.1.e992.jpg/230px-Live_Tetragnatha_montana_%28RMNH.ARA.14127%29_parasitized_by_Acrodactyla_quadrisculpta_larva_%28RMNH.INS.593867%29_-_BDJ.1.e992.jpg)


Micropredators
[edit]
A micropredator attacks more than one host, reducing each host's fitness by at least a small amount, and is only in contact with any one host intermittently. This behavior makes micropredators suitable as vectors, as they can pass smaller parasites from one host to another.[22][34][23] Most micropredators are hematophagic, feeding on blood. They include annelids such as leeches, crustaceans such as branchiurans and gnathiid isopods, various dipterans such as mosquitoes and tsetse flies, other arthropods such as fleas and ticks, vertebrates such as lampreys, and mammals such as vampire bats.[22]
Transmission strategies
[edit]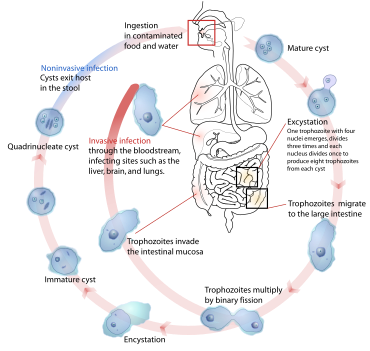
Parasites use a variety of methods to infect animal hosts, including physical contact, the fecal–oral route, free-living infectious stages, and vectors, suiting their differing hosts, life cycles, and ecological contexts.[35] Examples to illustrate some of the many possible combinations are given in the table.
| Parasite | Host | Transmission method | Ecological context |
|---|---|---|---|
| Gyrodactylus turnbulli (a monogenean) |
Poecilia reticulata (guppy) |
physical contact | social behaviour |
| Nematodes e.g. Strongyloides |
Macaca fuscata (Japanese macaque) |
fecal–oral |
social behaviour |
| Heligmosomoides polygyrus (a nematode) |
Apodemus flavicollis (yellow-necked mouse) |
fecal–oral | sex-biased transmission (mainly to males) |
| Amblyomma (a tick) |
Sphenodon punctatus (tuatara) |
free-living infectious stages | social behaviour |
| Plasmodium (malaria parasite) |
Birds, mammals (inc. humans) |
Anopheles mosquito vector, attracted by odour of infected human host[36] |
— |
Variations
[edit]Among the many variations on parasitic strategies are hyperparasitism,[37] social parasitism,[38] brood parasitism,[39] kleptoparasitism,[40] sexual parasitism,[41] and adelphoparasitism.[42]
Hyperparasitism
[edit]Hyperparasites feed on another parasite, as exemplified by protozoa living in helminth parasites,[37] or facultative or obligate parasitoids whose hosts are either conventional parasites or parasitoids.[22][33] Levels of parasitism beyond secondary also occur, especially among facultative parasitoids. In oak gall systems, there can be up to five levels of parasitism.[43]
Hyperparasites can control their hosts' populations, and are used for this purpose in agriculture and to some extent in medicine. The controlling effects can be seen in the way that the CHV1 virus helps to control the damage that chestnut blight, Cryphonectria parasitica, does to American chestnut trees, and in the way that bacteriophages can limit bacterial infections. It is likely, though little researched, that most pathogenic microparasites have hyperparasites which may prove widely useful in both agriculture and medicine.[44]
Social parasitism
[edit]Social parasites take advantage of interspecific interactions between members of eusocial animals such as ants, termites, and bumblebees. Examples include the large blue butterfly, Phengaris arion, its larvae employing ant mimicry to parasitise certain ants,[38] Bombus bohemicus, a bumblebee which invades the hives of other bees and takes over reproduction while their young are raised by host workers, and Melipona scutellaris, a eusocial bee whose virgin queens escape killer workers and invade another colony without a queen.[45] An extreme example of interspecific social parasitism is found in the ant Tetramorium inquilinum, an obligate parasite which lives exclusively on the backs of other Tetramorium ants.[46] A mechanism for the evolution of social parasitism was first proposed by Carlo Emery in 1909.[47] Now known as "Emery's rule", it states that social parasites tend to be closely related to their hosts, often being in the same genus.[48][49][50]
Intraspecific social parasitism occurs in parasitic nursing, where some individual young take milk from unrelated females. In wedge-capped capuchins, higher ranking females sometimes take milk from low ranking females without any reciprocation.[51]
Brood parasitism
[edit]In brood parasitism, the hosts act as parents as they raise the young as their own. Brood parasites include birds in different families such as cowbirds, whydahs, cuckoos, and black-headed ducks. These do not build nests of their own, but leave their eggs in nests of other species. The eggs of some brood parasites mimic those of their hosts, while some cowbird eggs have tough shells, making them hard for the hosts to kill by piercing, both mechanisms implying selection by the hosts against parasitic eggs.[39][52][53] The adult female European cuckoo further mimics a predator, the European sparrowhawk, giving her time to lay her eggs in the host's nest unobserved.[54]
Kleptoparasitism
[edit]In kleptoparasitism (from Greek κλέπτης (kleptēs), "thief"), parasites steal food gathered by the host. The parasitism is often on close relatives, whether within the same species or between species in the same genus or family. For instance, the many lineages of cuckoo bees lay their eggs in the nest cells of other bees in the same family.[40] Kleptoparasitism is uncommon generally but conspicuous in birds; some such as skuas are specialised in pirating food from other seabirds, relentlessly chasing them down until they disgorge their catch.[55]
Sexual parasitism
[edit]A unique approach is seen in some species of anglerfish, such as Ceratias holboelli, where the males are reduced to tiny sexual parasites, wholly dependent on females of their own species for survival, permanently attached below the female's body, and unable to fend for themselves. The female nourishes the male and protects him from predators, while the male gives nothing back except the sperm that the female needs to produce the next generation.[41]
Adelphoparasitism
[edit]Adelphoparasitism, (from Greek ἀδελφός (adelphós), brother[56]), also known as sibling-parasitism, occurs where the host species is closely related to the parasite, often in the same family or genus.[42] In the citrus blackfly parasitoid, Encarsia perplexa, unmated females may lay haploid eggs in the fully developed larvae of their own species, producing male offspring,[57] while the marine worm Bonellia viridis has a similar reproductive strategy, although the larvae are planktonic.[58]
Illustrations
[edit]Examples of the major variant strategies are illustrated.
-
 A hyperparasitoid pteromalid wasp on the cocoons of its host, itself a parasitoid braconid wasp
A hyperparasitoid pteromalid wasp on the cocoons of its host, itself a parasitoid braconid wasp -
 The large blue butterfly is an ant mimic and social parasite.
The large blue butterfly is an ant mimic and social parasite. -
 In brood parasitism, the host raises the young of another species, here a cowbird's egg, that has been laid in its nest.
In brood parasitism, the host raises the young of another species, here a cowbird's egg, that has been laid in its nest. -
 The great skua is a powerful kleptoparasite, relentlessly pursuing other seabirds until they disgorge their catches of food.
The great skua is a powerful kleptoparasite, relentlessly pursuing other seabirds until they disgorge their catches of food. -
 The male of the anglerfish species Ceratias holboelli lives as a tiny sexual parasite permanently attached below the female's body.
The male of the anglerfish species Ceratias holboelli lives as a tiny sexual parasite permanently attached below the female's body. -
 Encarsia perplexa (centre), a parasitoid of citrus blackfly (lower left), is also an adelphoparasite, laying eggs in larvae of its own species
Encarsia perplexa (centre), a parasitoid of citrus blackfly (lower left), is also an adelphoparasite, laying eggs in larvae of its own species






Taxonomic range
[edit]Parasitism has an extremely wide taxonomic range, including animals, plants, fungi, protozoans, bacteria, and viruses.[59]
Animals
[edit]| Phylum | Class/Order | No. of species |
Endo- paras. |
Ecto- paras. |
Invert def. host |
Vert def. host |
No. of hosts |
Marine | Fresh- water |
Terres- trial |
|---|---|---|---|---|---|---|---|---|---|---|
| Cnidaria | Myxozoa | 1,350 | Yes | Yes | 2 or more | Yes | Yes | |||
| Cnidaria | Polypodiozoa | 1 | Yes | Yes | 1 | Yes | ||||
| Flatworms | Trematodes | 15,000 | Yes | Yes | 2 or more | Yes | Yes | Yes | ||
| Flatworms | Monogeneans | 20,000 | Yes | Yes | 1 | Yes | Yes | |||
| Flatworms | Cestodes | 5,000 | Yes | Yes | 2 or more | Yes | Yes | Yes | ||
| Horsehair worms | 350 | Yes | Yes | 1 or more | Yes | Yes | ||||
| Nematodes | 10,500 | Yes | Yes | Yes | 1 or more | Yes | Yes | Yes | ||
| Acanthocephala | 1,200 | Yes | Yes | 2 or more | Yes | Yes | Yes | |||
| Annelids | Leeches | 400 | Yes | Yes | 1 | Yes | Yes | |||
| Molluscs | Bivalves | 600 | Yes | Yes | 1 | Yes | ||||
| Molluscs | Gastropods | 5,000 | Yes | Yes | 1 | Yes | ||||
| Arthropods | Ticks | 800 | Yes | Yes | 1 or more | Yes | ||||
| Arthropods | Mites | 30,000 | Yes | Yes | Yes | Yes | 1 | Yes | Yes | Yes |
| Arthropods | Copepods | 4,000 | Yes | Yes | Yes | 1 | Yes | Yes | ||
| Arthropods | Lice | 4,000 | Yes | Yes | 1 | Yes | ||||
| Arthropods | Fleas | 2,500 | Yes | Yes | 1 | Yes | ||||
| Arthropods | True flies | 2,300 | Yes | Yes | 1 | Yes | ||||
| Arthropods | Twisted-wing insects | 600 | Yes | Yes | 1 | Yes | ||||
| Arthropods | Parasitoid wasps | 130,000[61] - 1,100,000[62] | Yes | Yes | Yes | 1 | Yes |
Parasitism is widespread in the animal kingdom,[63] and has evolved independently from free-living forms hundreds of times.[22] Many types of helminth including flukes and cestodes have complete life cycles involving two or more hosts. By far the largest group is the parasitoid wasps in the Hymenoptera.[22] The phyla and classes with the largest numbers of parasitic species are listed in the table. Numbers are conservative minimum estimates. The columns for Endo- and Ecto-parasitism refer to the definitive host, as documented in the Vertebrate and Invertebrate columns.[60]
Plants
[edit]
A hemiparasite or partial parasite such as mistletoe derives some of its nutrients from another living plant, whereas a holoparasite such as dodder derives all of its nutrients from another plant.[64] Parasitic plants make up about one per cent of angiosperms and are in almost every biome in the world.[65][66][67] All these plants have modified roots, haustoria, which penetrate the host plants, connecting them to the conductive system—either the xylem, the phloem, or both. This provides them with the ability to extract water and nutrients from the host. A parasitic plant is classified depending on where it latches onto the host, either the stem or the root, and the amount of nutrients it requires. Since holoparasites have no chlorophyll and therefore cannot make food for themselves by photosynthesis, they are always obligate parasites, deriving all their food from their hosts.[66] Some parasitic plants can locate their host plants by detecting chemicals in the air or soil given off by host shoots or roots, respectively. About 4,500 species of parasitic plant in approximately 20 families of flowering plants are known.[68][66]
Species within the Orobanchaceae (broomrapes) are among the most economically destructive of all plants. Species of Striga (witchweeds) are estimated to cost billions of dollars a year in crop yield loss, infesting over 50 million hectares of cultivated land within Sub-Saharan Africa alone. Striga infects both grasses and grains, including corn, rice, and sorghum, which are among the world's most important food crops. Orobanche also threatens a wide range of other important crops, including peas, chickpeas, tomatoes, carrots, and varieties of cabbage. Yield loss from Orobanche can be total; despite extensive research, no method of control has been entirely successful.[69]
Many plants and fungi exchange carbon and nutrients in mutualistic mycorrhizal relationships. Some 400 species of myco-heterotrophic plants, mostly in the tropics, however effectively cheat by taking carbon from a fungus rather than exchanging it for minerals. They have much reduced roots, as they do not need to absorb water from the soil; their stems are slender with few vascular bundles, and their leaves are reduced to small scales, as they do not photosynthesize. Their seeds are very small and numerous, so they appear to rely on being infected by a suitable fungus soon after germinating.[70]

Fungi
[edit]Parasitic fungi derive some or all of their nutritional requirements from plants, other fungi, or animals.
Plant pathogenic fungi are classified into three categories depending on their mode of nutrition: biotrophs, hemibiotrophs and necrotrophs. Biotrophic fungi derive nutrients from living plant cells, and during the course of infection they colonise their plant host in such a way as to keep it alive for a maximally long time.[71] One well-known example of a biotrophic pathogen is Ustilago maydis, causative agent of the corn smut disease. Necrotrophic pathogens on the other hand, kill host cells and feed saprophytically, an example being the root-colonising honey fungi in the genus Armillaria.[72] Hemibiotrophic pathogens begin their colonising their hosts as biotrophs, and subsequently killing off host cells and feeding as necrotrophs, a phenomenon termed the biotrophy-necrotrophy switch.[73]
Pathogenic fungi are well-known causative agents of diseases on animals as well as humans. Fungal infections (mycosis) are estimated to kill 1.6 million people each year.[74] One example of a potent fungal animal pathogen are Microsporidia - obligate intracellular parasitic fungi that largely affect insects, but may also affect vertebrates including humans, causing the intestinal infection microsporidiosis.[75]
Protozoa
[edit]Protozoa such as Plasmodium, Trypanosoma, and Entamoeba[76] are endoparasitic. They cause serious diseases in vertebrates including humans—in these examples, malaria, sleeping sickness, and amoebic dysentery—and have complex life cycles.[31]
Bacteria
[edit]Many bacteria are parasitic, though they are more generally thought of as pathogens causing disease.[77] Parasitic bacteria are extremely diverse, and infect their hosts by a variety of routes. To give a few examples, Bacillus anthracis, the cause of anthrax, is spread by contact with infected domestic animals; its spores, which can survive for years outside the body, can enter a host through an abrasion or may be inhaled. Borrelia, the cause of Lyme disease and relapsing fever, is transmitted by vectors, ticks of the genus Ixodes, from the diseases' reservoirs in animals such as deer. Campylobacter jejuni, a cause of gastroenteritis, is spread by the fecal–oral route from animals, or by eating insufficiently cooked poultry, or by contaminated water. Haemophilus influenzae, an agent of bacterial meningitis and respiratory tract infections such as influenza and bronchitis, is transmitted by droplet contact. Treponema pallidum, the cause of syphilis, is spread by sexual activity.[78]

Viruses
[edit]Viruses are obligate intracellular parasites, characterised by extremely limited biological function, to the point where, while they are evidently able to infect all other organisms from bacteria and archaea to animals, plants and fungi, it is unclear whether they can themselves be described as living. They can be either RNA or DNA viruses consisting of a single or double strand of genetic material (RNA or DNA, respectively), covered in a protein coat and sometimes a lipid envelope. They thus lack all the usual machinery of the cell such as enzymes, relying entirely on the host cell's ability to replicate DNA and synthesise proteins. Most viruses are bacteriophages, infecting bacteria.[79][80][81][82]
Evolutionary ecology
[edit]
Parasitism is a major aspect of evolutionary ecology; for example, almost all free-living animals are host to at least one species of parasite. Vertebrates, the best-studied group, are hosts to between 75,000 and 300,000 species of helminths and an uncounted number of parasitic microorganisms. On average, a mammal species hosts four species of nematode, two of trematodes, and two of cestodes.[83] Humans have 342 species of helminth parasites, and 70 species of protozoan parasites.[84] Some three-quarters of the links in food webs include a parasite, important in regulating host numbers. Perhaps 40 per cent of described species are parasitic.[83]
Fossil record
[edit]Parasitism is hard to demonstrate from the fossil record, but holes in the mandibles of several specimens of Tyrannosaurus may have been caused by Trichomonas-like parasites.[85] Saurophthirus, the Early Cretaceous flea, parasitized pterosaurs.[86][87] Eggs that belonged to nematode worms and probably protozoan cysts were found in the Late Triassic coprolite of phytosaur. This rare find in Thailand reveals more about the ecology of prehistoric parasites.[88]
Coevolution
[edit]As hosts and parasites evolve together, their relationships often change. When a parasite is in a sole relationship with a host, selection drives the relationship to become more benign, even mutualistic, as the parasite can reproduce for longer if its host lives longer.[89] But where parasites are competing, selection favours the parasite that reproduces fastest, leading to increased virulence. There are thus varied possibilities in host–parasite coevolution.[90]
Evolutionary epidemiology analyses how parasites spread and evolve, whereas Darwinian medicine applies similar evolutionary thinking to non-parasitic diseases like cancer and autoimmune conditions.[91]
Coevolution favouring mutualism
[edit]
Long-term coevolution sometimes leads to a relatively stable relationship tending to commensalism or mutualism, as, all else being equal, it is in the evolutionary interest of the parasite that its host thrives. A parasite may evolve to become less harmful for its host or a host may evolve to cope with the unavoidable presence of a parasite—to the point that the parasite's absence causes the host harm. For example, although animals parasitised by worms are often clearly harmed, such infections may also reduce the prevalence and effects of autoimmune disorders in animal hosts, including humans.[89] In a more extreme example, some nematode worms cannot reproduce, or even survive, without infection by Wolbachia bacteria.[92]
Lynn Margulis and others have argued, following Peter Kropotkin's 1902 Mutual Aid: A Factor of Evolution, that natural selection drives relationships from parasitism to mutualism when resources are limited. This process may have been involved in the symbiogenesis which formed the eukaryotes from an intracellular relationship between archaea and bacteria, though the sequence of events remains largely undefined.[93][94]
Competition favouring virulence
[edit]Competition between parasites can be expected to favour faster reproducing and therefore more virulent parasites, by natural selection.[90][95]

Among competing parasitic insect-killing bacteria of the genera Photorhabdus and Xenorhabdus, virulence depended on the relative potency of the antimicrobial toxins (bacteriocins) produced by the two strains involved. When only one bacterium could kill the other, the other strain was excluded by the competition. But when caterpillars were infected with bacteria both of which had toxins able to kill the other strain, neither strain was excluded, and their virulence was less than when the insect was infected by a single strain.[90]
Cospeciation
[edit]A parasite sometimes undergoes cospeciation with its host, resulting in the pattern described in Fahrenholz's rule, that the phylogenies of the host and parasite come to mirror each other.[96]
An example is between the simian foamy virus (SFV) and its primate hosts. The phylogenies of SFV polymerase and the mitochondrial cytochrome c oxidase subunit II from African and Asian primates were found to be closely congruent in branching order and divergence times, implying that the simian foamy viruses cospeciated with Old World primates for at least 30 million years.[97]
The presumption of a shared evolutionary history between parasites and hosts can help elucidate how host taxa are related. For instance, there has been a dispute about whether flamingos are more closely related to storks or ducks. The fact that flamingos share parasites with ducks and geese was initially taken as evidence that these groups were more closely related to each other than either is to storks. However, evolutionary events such as the duplication, or the extinction of parasite species (without similar events on the host phylogeny) often erode similarities between host and parasite phylogenies. In the case of flamingos, they have similar lice to those of grebes. Flamingos and grebes do have a common ancestor, implying cospeciation of birds and lice in these groups. Flamingo lice then switched hosts to ducks, creating the situation which had confused biologists.[98]

Parasites infect sympatric hosts (those within their same geographical area) more effectively, as has been shown with digenetic trematodes infecting lake snails.[99] This is in line with the Red Queen hypothesis, which states that interactions between species lead to constant natural selection for coadaptation. Parasites track the locally common hosts' phenotypes, so the parasites are less infective to allopatric hosts, those from different geographical regions.[99]
Modifying host behaviour
[edit]
Some parasites modify host behaviour in order to increase their transmission between hosts, often in relation to predator and prey (parasite increased trophic transmission). For example, in the California coastal salt marsh, the fluke Euhaplorchis californiensis reduces the ability of its killifish host to avoid predators.[100] This parasite matures in egrets, which are more likely to feed on infected killifish than on uninfected fish. Another example is the protozoan Toxoplasma gondii, a parasite that matures in cats but can be carried by many other mammals. Uninfected rats avoid cat odors, but rats infected with T. gondii are drawn to this scent, which may increase transmission to feline hosts.[101] The malaria parasite modifies the skin odour of its human hosts, increasing their attractiveness to mosquitoes and hence improving the chance that the parasite will be transmitted.[36] The spider Cyclosa argenteoalba often have parasitoid wasp larvae attached to them which alter their web-building behavior. Instead of producing their normal sticky spiral shaped webs, they made simplified webs when the parasites were attached. This manipulated behavior lasted longer and was more prominent the longer the parasites were left on the spiders.[102]

Trait loss
[edit]Parasites can exploit their hosts to carry out a number of functions that they would otherwise have to carry out for themselves. Parasites which lose those functions then have a selective advantage, as they can divert resources to reproduction. Many insect ectoparasites including bedbugs, batbugs, lice and fleas have lost their ability to fly, relying instead on their hosts for transport.[103] Trait loss more generally is widespread among parasites.[104] An extreme example is the myxosporean Henneguya zschokkei, an ectoparasite of fish and the only animal known to have lost the ability to respire aerobically: its cells lack mitochondria.[105]
Host defences
[edit]Хозяева разработали множество защитных мер против своих паразитов, включая физические барьеры, такие как кожа позвоночных, [106] the immune system of mammals,[107] insects actively removing parasites,[108] and defensive chemicals in plants.[109]
Эволюционный биолог В.Д. Гамильтон предположил, что сексуальное размножение могло бы развиваться, чтобы помочь победить множественные паразиты, обеспечивая генетическую рекомбинацию , перетасование генов для создания различных комбинаций. Математическое моделирование Гамильтон показал, что сексуальное воспроизведение будет эволюционно стабильным в разных ситуациях, и что прогнозы теории соответствовали фактической экологии сексуального размножения. [ 110 ] [ 111 ] Тем не менее, может быть компромисс между иммунокомпетентностью мужских позвоночных и размножением вторичных половых характеристик , такими как оперение павлина и грибы львов . Это связано с тем, что мужской гормон тестостерон поощряет рост вторичных половых характеристик, предпочитая таких мужчин в сексуальном отборе , по цене снижения их иммунной защиты. [ 112 ]
Позвоночные
[ редактировать ]
Физический барьер жесткой и часто сухой и водонепроницаемой кожи рептилий, птиц и млекопитающих удерживает вторжение микроорганизмов в организм в организм. Человеческая кожа также выделяет кожную кожную , которая токсична для большинства микроорганизмов. [ 106 ] С другой стороны, более крупные паразиты, такие как трематоды, обнаруживают химические вещества, полученные кожей, чтобы найти своих хозяев, когда они попадают в воду. и слезы позвоночных Слюна содержат лизоцим , фермент, который разрушает клеточные стенки вторгающихся бактерий. [ 106 ] Если организм пройдет в рот, желудок с его соляной кислотой , токсичной для большинства микроорганизмов, является следующей линией защиты. [ 106 ] Некоторые кишечные паразиты имеют толстое, крепкое внешнее покрытие, которое медленно или нет, что позволяет паразиту проходить через желудок живым, после чего они попадают в кишечник и начинают следующую стадию своей жизни. Оказавшись внутри тела, паразиты должны преодолеть иммунной системы и сывороточные белки рецепторы распознавания картины адаптивной иммунной системы, , внутриклеточные и клеточные, которые запускают лимфоциты такие как Т-клетки и антитело, производящие B-клетки . У них есть рецепторы, которые распознают паразитов. [ 107 ]
Насекомые
[ редактировать ]
Насекомые часто адаптируют свои гнезда, чтобы уменьшить паразитизм. Например, одна из ключевых причин, по которой оса Polistes Canadensis гнездятся по нескольким расческам , а не строить одну расческу, как большую часть остальной части его рода, заключается в том, чтобы избежать заражения тинидными мотыльками . Тинеидная моль откладывает яйца в гнездах ос, а затем эти яйца вы цепляются в личинки, которые могут забросить от клетки к клетке и добывать куколки осы. Взрослые осы пытаются удалить и убить яйца моли и личинок, пережевывая края клеток, покрывая клетки пероральной секрецией, которая придает гнезду темно -коричневато -коричневатую внешность. [ 108 ]
Растения
[ редактировать ]Растения реагируют на атаку паразитов серией химической защиты, такой как полифенолоксидаза , под контролем сигнальных путей , нечувствительных к ясмоновой кислоте (JA) и салициловой кислоты (SA). [ 109 ] [ 113 ] Различные биохимические пути активируются различными атаками, и эти два пути могут взаимодействовать положительно или отрицательно. В целом, растения могут либо инициировать конкретный или неспецифический ответ. [ 114 ] [ 113 ] Конкретные ответы включают распознавание паразита клеточными рецепторами растения, что приводит к сильной, но локализованной реакции: оборонительные химические вещества производятся вокруг области, где был обнаружен паразит, блокируя его распространение и избегая тратить оборонительное производство, где он не нужен. [ 114 ] Неспособные защитные реакции являются системными, а это означает, что ответы не ограничиваются площадью растения, а распространяются по всему растению, что делает их дорогостоящими по энергии. Они эффективны против широкого спектра паразитов. [ 114 ] При повреждении, например, с лепидоптераном гусеницы , листья растений, включая высвобождение кукурузы и хлопка, увеличивалось количество летучих химических веществ, таких как терпены , которые сигнализируют о том, что на них атакованы; Одним из эффектов этого является привлечение паразитоидных ос, которые, в свою очередь, атакуют гусениц. [ 115 ]
Биология и сохранение
[ редактировать ]Экология и паразитология
[ редактировать ]
Паразитизм и эволюция паразитов были до двадцати первого века, изучаемого паразитологами , в науке, в которой преобладают медицина, а не экологи или эволюционные биологи . Несмотря на то, что взаимодействие паразитов-хост было явно экологическим и важным в эволюции, история паразитологии вызвала то, что эволюционный эколог Роберт Пулин назвал «поглощением паразитизма паразитологами», что привело к тому, что экологи игнорируют область. По его мнению, это было «несчастным», поскольку паразиты являются «вездесущими агентами естественного отбора» и значительными силами в эволюции и экологии. [ 116 ] По его мнению, давний раскол между науками ограничивал обмен идей, отдельными конференциями и отдельными журналами. Технические языки экологии и паразитологии иногда включали различные значения для одних и тех же слов. Были также философские различия: Пулен отмечает, что, под влиянием медицины, «многие паразитологи признали, что эволюция привела к снижению вирулентности паразитов, тогда как современная эволюционная теория предсказала бы больший диапазон результатов». [ 116 ]
Их сложные отношения затрудняют размещение паразитов в пищевых сетях: трематод с несколькими хостами для его различных этапов жизненного цикла займет многие позиции в продовольственной сети одновременно и создаст петли потока энергии, что путает анализ. Кроме того, поскольку почти у каждого животного есть (множественные) паразиты, паразиты будут занимать высшие уровни каждой пищевой сети. [ 84 ]
Паразиты могут играть роль в пролиферации неродных видов. Например, инвазивные зеленые крабы минимально затрагивают местные трематоды на восточном атлантическом побережье. Это помогает им переоценить местных крабов, таких как Атлантический Рок и Иона Крабс . [ 117 ]
Экологическая паразитология может быть важна для попыток контроля, как во время кампании по уничтожению морского червя . Несмотря на то, что паразит был уничтожен во всех странах, кроме четырех, червь начал использовать лягушек в качестве посредника до заражения собак, что затрудняет контроль, чем было бы, если бы отношения были лучше поняты. [ 118 ]
Обоснование сохранения
[ редактировать ]| Внешние видео | |
|---|---|
Хотя паразиты считаются вредными, искоренение всех паразитов не будет полезно. Паразиты составляют не менее половины жизни разнообразия жизни; Они выполняют важные экологические роли; И без паразитов организмы могут склонны к беспощеническому размножению, уменьшая разнообразие признаков, вызванных сексуальным размножением. [ 119 ] Паразиты предоставляют возможность для переноса генетического материала между видами, способствуя эволюционным изменениям. [ 120 ] Многие паразиты требуют нескольких хозяев разных видов, чтобы завершить свои жизненные циклы и полагаться на Predator-Prey или другие стабильные экологические взаимодействия, чтобы добраться от одного хозяина к другому. Таким образом, присутствие паразитов указывает на то, что экосистема является здоровой. [ 121 ]
Эктопаразит, калифорнийский кондор, Colpocephalum californici , стал известной проблемой сохранения. В Соединенных Штатах была проведена крупная и очень дорогостоящая программа разведения в неволе, чтобы спасти калифорнийского кондора . Это было хозяином вошей, которая жила только на нем. Любые найденные вши были «намеренно убиты» во время программы, чтобы поддерживать кондоров в наилучшем состоянии здоровья. В результате один вид, кондор, был спасен и возвращен в дикую природу, в то время как другой вид, паразит, вымер. [ 122 ]
Хотя паразиты часто пропускаются в изображениях пищевых сетей , они обычно занимают верхнюю позицию. Паразиты могут функционировать как виды Keystone , уменьшая доминирование превосходных конкурентов и позволяя конкурирующих видов . сосуществовать [ 84 ] [ 123 ] [ 124 ]

Количественная экология
[ редактировать ]Один виды паразитов обычно имеет агрегированное распределение среди животных -хозяев, что означает, что большинство хозяев носят мало паразитов, в то время как несколько хозяев несут подавляющее большинство людей паразитов. Это создает значительные проблемы для учащихся паразитов экологии, поскольку оно делает параметрическую статистику , как обычно используется биологами недействительными. Лог-трансформация данных перед применением параметрического теста или использование непараметрической статистики рекомендуется несколькими авторами, но это может привести к дальнейшим проблемам, поэтому количественная паразитология основана на более продвинутых биостатистических методах. [ 125 ]
История
[ редактировать ]Древний
[ редактировать ]Человеческие паразиты , включая круглые черви, морской червя , червя, ниточки и ленточные черви, упоминаются в записях египетского папируса с 3000 г. до н.э. Эберс папирус описывает анкилостому галосту . В Древней Греции паразиты, включая червя мочевого пузыря , описаны в корпусе Гиппократа , в то время как комический драматург Аристофана называется ленты «градами». Римские врачи Цельс и Гален задокументировали круглые черви Ascaris Lumbricoides и Enterobius vermicularis . [ 126 ]
Средневековый
[ редактировать ]
В своем каноне медицины , завершенном в 1025 году, персидский врач Авиченна записал паразитов человека и животных, включая круглые черви, нитовые червя, червя Гвинеи и ленты. [ 126 ]
В своей книге 1397 года Traité de L'Etat, Science et pratique de l'Art de la Bergerie (рассказ о государстве, науке и практике искусства Шепердинга) Джехан де Бри написал первое описание эндопаразита трематода , Овечья печеночная фасляка Hepatica . [ 127 ] [ 128 ]
Ранний современный
[ редактировать ]В раннем современном периоде книга Реди Франческо в 1668 году Esperienze intorno alla generazione degl'insetti ( опыт генерации насекомых ), явно описанные экто- и эндопаразиты, иллюстрирующие клещи, личинки назальных мух оленя и овечьи печени, иллюстрирующие клещи , личинки назальных мух оленя и овечьи печени, иллюстрирующие клещи, личинки назальных мух оленя и овечьи Полем [ 129 ] Реди отметил, что паразиты развиваются из яиц, противоречате теории спонтанного поколения . [ 130 ] В своих наблюдениях за книгами 1684 года о живых животных, обнаруженных у живых животных ( наблюдения за живыми животными, обнаруженными у живых животных ), Реди описал и проиллюстрировал более 100 паразитов, включая большой круглый червь у людей, который вызывает аскорис . [ 129 ] Реди был первым, кто назвал кисты Echinococcus granulosus, рассматриваемые у собак и овец как паразитные; Спустя столетие, в 1760 году, Питер Саймон Паллас правильно предположил, что это были личинки ленточных червей. [ 126 ]
В 1681 году Энтони Ван Леувенхук наблюдала и проиллюстрировала простейшего паразита Джардии Ламблия и связала его с «своими собственными свободными табуретками». Это был первый простейший паразит людей, который был замечен под микроскопом. [ 126 ] Несколько лет спустя, в 1687 году, итальянские биологи Джованни Козимо Бономо и Диасинто Цестони описали Скаби , вызванные паразитными саркорами клеща , а также отмечая его как первое заболевание людей с известным микроскопическим возбудителем. [ 131 ]

Паразитология
[ редактировать ]Современная паразитология, разработанная в 19 веке с точными наблюдениями и экспериментами многих исследователей и врачей; [ 127 ] Термин был впервые использован в 1870 году. [ 132 ] В 1828 году Джеймс Аннерсли описал амебиаз , простейшие инфекции кишечника и печень, хотя патоген, etamoeba histolytica , не был обнаружен до 1873 года Фридрихом Лёш. Джеймс Педжет обнаружил кишечную нематоду Trichinella Spiralis у людей в 1835 году. Джеймс Макконнелл описал человеческую печень, Clonorchis sinensis , в 1875 году. [ 126 ] Алджернон Томас и Рудольф Лейкарт независимо сделали первое открытие жизненного цикла трематоды, овечьей печени, экспериментом в 1881–1883 годах. [ 127 ] В 1877 году Патрик Мэнсон обнаружил жизненный цикл филариальных червей , которые вызывают слоненз, передаваемый комарами. Мансон также предсказал, что у паразита малярии , Plasmodium , был вектор комаров и убедил Рональда Росса расследовать. Росс подтвердил, что прогноз был правильным в 1897–1898 годах. В то же время, Джованни Баттиста Грасси и другие описали стадии жизненного цикла паразита малярии у комаров Анофелеса . Росс был спорно награжден Нобелевской премией 1902 года за свою работу, а Грасси не было. [ 126 ] В 1903 году Дэвид Брюс идентифицировал простейшего паразита и мух с циксом вектор африканского трипаносомоза . [ 133 ]
Вакцина
[ редактировать ]Учитывая важность малярии, когда около 220 миллионов человек заражаются ежегодно, было предпринято много попыток прервать ее передачу. Были опробованы различные методы профилактики малярии , включая использование противомалярийных препаратов для убийства паразитов в крови, ликвидации векторов комаров с органохлорином и другими инсектицидами и развитие вакцины против малярии . Все это оказалось проблематичной, с лекарственной устойчивостью , устойчивостью к инсектицидам среди комаров и повторной недостаточностью вакцин в качестве паразита. [ 134 ] Первая и по состоянию на 2015 год единственной лицензированной вакциной для любого паразитического заболевания людей является RTS, S для малярии Plasmodium falciparum . [ 135 ]
Биологический контроль
[ редактировать ]
Несколько групп паразитов, включая микробные патогены и паразитоидальные осы, использовались в качестве биологических контрольных агентов в сельском хозяйстве и садоводстве . [ 137 ] [ 138 ]
Сопротивление
[ редактировать ]Пулен отмечает, что широко распространенное профилактическое употребление антифельминтных препаратов у домашних овец и крупного рогатого скота представляет собой всемирный контролируемый эксперимент ООН в эволюции их паразитов. Результаты зависят от того, уменьшают ли препараты вероятность того, что личинка гельминта достигнет взрослой жизни. Если это так, можно ожидать, что естественный отбор будет способствовать производству яиц в более раннем возрасте. Если, с другой стороны, препараты в основном влияют на паразитических червей взрослых , отбор может вызвать отсроченную зрелость и повышенную вирулентность . Такие изменения, по -видимому, ведутся: нематода Teladorsagia Curincincta меняет свой размер взрослого и репродуктивную частоту в ответ на лекарства. [ 139 ]
Культурное значение
[ редактировать ]
Классические времена
[ редактировать ]В классическую эпоху концепция паразита не была строго уничижительной: был общепринятой роль паразит терпеть унижение ». [ 140 ] [ 141 ]
Общество
[ редактировать ]Паразитизм имеет уничижительный смысл в популярном использовании. По словам иммунолога Джона Плейфэра, [ 142 ]
В повседневной речи термин «паразит» загружен унизительным значением. Паразит - это спонсор, ленивый спекулятор, утечка общества. [ 142 ]
Сатирический : священнослужитель Джонатан Свифт намекает на гиперпаразитизм в своем стихотворении 1733 года «о поэзии: рапсодию», сравнивая поэтов с «паразитами», которые «дают и ущипнут свои враги» [ 143 ]
Слоры только Тиз и ущипнуть
Их враги превосходят на дюйм.
Итак, Nat'ralists наблюдают, блоха
Есть меньшие блохи, которые на него добывают;
И у них есть меньшие блохи, чтобы укусить их.
И так продолжается ad infinitum .
Таким образом, каждый поэт в своем роде,
Он немного позади:
В исследовании 2022 года изучалось именование около 3000 видов паразитов, обнаруженных в предыдущие два десятилетия. Из тех, названных в честь ученых, более 80% были названы в честь мужчин, тогда как около трети авторов документов на паразитах были женщины. Исследование показало, что процент видов паразитов, названных для родственников или друзей автора, резко возрос за тот же период. [ 144 ]
Вымысел
[ редактировать ]
В Брэма Стокера « ужасов готическом романе Дракула» «Дракула » и его многочисленные экранизации , одноименное граф Дракула -это паразит с пьющим кровь (вампир). Критик Лаура Отис утверждает, что как «вор, соблазнитель, создатель и имитируют, Дракула - конечный паразит. Весь смысл вампиризма сосает кровь других людей - к крови других людей за счет других людей». [ 145 ]
Отвратительные и ужасающие паразитические инопланетные виды широко распространены в научной фантастике , [ 146 ] [ 147 ] как, например, в фильме Ридли Скотта 1979 года «Чужой» . [ 148 ] [ 149 ] В одной сцене ксеноморф вырывается из груди мертвого человека, а кровь брызгает под высоким давлением, которым помогает взрывчатые скважины . Органы животных были использованы для усиления шокового эффекта. Сцена была снята за один раз, и испуганная реакция актеров была подлинной. [ 4 ] [ 150 ]
Смотрите также
[ редактировать ]- Антаркатический
- Канцерогенный паразит
- Влияние паразитических червей на иммунную систему
- Список паразитов людей
Примечания
[ редактировать ]- ^ Трофически переданные паразиты передаются на их окончательный хост, хищник, когда их промежуточный хозяин едят. Эти паразиты часто модифицируют поведение своих промежуточных хозяев, заставляя их вести себя таким образом, что делает их вероятными, например, поднимаясь до заметной точки: это получает паразиты, передаваемые за счет жизни промежуточного хозяина.
- ^ Волк - это социальный хищник, охота в упаковках; Пумы - одиночный хищник, охота в одиночку. Ни одна стратегия обычно не считается паразитической. [ 23 ]
Ссылки
[ редактировать ]- ^ Poulin 2007 , с. 4–5.
- ^ Jump up to: а беременный Уилсон, Эдвард О. (2014). Значение человеческого существования . WW Norton & Company. п. 112. ISBN 978-0-87140-480-0 Полем
Паразиты, во фразе, являются хищниками, которые едят добычу в единицах менее одного. Допустимые паразиты - это те, которые развивались, чтобы обеспечить их собственное выживание и размножение, но в то же время с минимальной болью и стоимостью для хозяина.
- ^ Getz, Wm (2011). «Сетя преобразования биомассы обеспечивают единый подход к моделированию потребительских ресурсов» . Экологические письма . 14 (2): 113–124. Bibcode : 2011 Ecoll..14..113G . doi : 10.1111/j.1461-0248.2010.01566.x . PMC 3032891 . PMID 21199247 .
- ^ Jump up to: а беременный «Создание сцены Челнстера Чужого» . Хранитель . 13 октября 2009 г. Архивировано с оригинала 30 апреля 2010 года . Получено 29 мая 2010 года .
- ^ Паразит , Лидделл, Генри Джордж; Скотт, Роберт, греко -английский лексикон , на цифровой библиотеке Персея
- ^ παρά , Генри Джордж Лидделл, Роберт Скотт, грек -английский лексикон , на цифровой библиотеке Персея
- ^ σῖτος , Лидделл, Генри Джордж; Скотт, Роберт, грик -английский лексикон , на цифровой библиотеке Персея
- ^ σιτισμός , Лидделл, Генри Джордж; Скотт, Роберт, грик -английский лексикон , на цифровой библиотеке Персея
- ^ Обзор паразитологии . Австралийское общество паразитологических и Австралийского исследовательского совета/Национальный совет по здравоохранению и медицинским исследованиям) исследовательская сеть для паразитологии. Июль 2010. ISBN 978-1-86499-991-4 Полем
Паразитизм - это форма симбиоза, интимная связь между двумя разными видами. Существует биохимическое взаимодействие между хозяином и паразитом; то есть они распознают друг друга, в конечном итоге на молекулярном уровне, и ткани хозяина стимулируются для каким -то образом реагировать. Это объясняет, почему паразитизм может привести к болезням, но не всегда.
- ^ Suzuki, Sayaki U.; Сасаки, Акира (2019). «Экологическая и эволюционная стабильность биотрофизма, некротрофизма и сапротрофизма» (PDF) . Американский натуралист . 194 (1): 90–103. doi : 10.1086/703485 . ISSN 0003-0147 . PMID 31251653 . S2CID 133349792 . Архивировано (PDF) из оригинала 6 марта 2020 года.
- ^ Rozsa, L.; Garay, J. (2023). «Определения паразитизма, учитывая его потенциально противоположное влияние на разных уровнях иерархической организации» . Паразитология . 150 (9): 761–768. doi : 10.1017/s0031182023000598 . PMC 10478066 . PMID 37458178 .
- ^ «Классификация животных паразитовых нематод» . plpnemweb.ucdavis.edu . Архивировано с оригинала 6 октября 2017 года . Получено 25 февраля 2016 года .
- ^ Гарсия, Л.С. (1999). «Классификация человеческих паразитов, векторов и подобных организмов» . Клинические инфекционные заболевания . 29 (4): 734–746. doi : 10.1086/520425 . PMID 10589879 .
- ^ Jump up to: а беременный в Обзор паразитологии . Австралийское общество паразитологических и Австралийского исследовательского совета/Национальный совет по здравоохранению и медицинским исследованиям) исследовательская сеть для паразитологии. Июль 2010. ISBN 978-1-86499-991-4 .
- ^ Веккионе, Анна; Aznar, Francisco Javier (2008). «Месопаразитный копепод Pennella balaenopterae и его значение как видимый индикатор состояния здоровья у дельфинов (Delphinidae): обзор» (PDF) . Журнал морских животных и их экологии . 7 (1): 4–11. Архивировано из оригинала (PDF) 10 апреля 2018 года . Получено 11 апреля 2018 года .
- ^ Jump up to: а беременный в дюймовый Пулин, Роберт (2011). Роллинсон, Д.; Hay, Si (ред.). «Многочисленные дороги к паразитизму: рассказ о конвергенции» . Достижения в области паразитологии . 74 Академическая пресса: 27–28. doi : 10.1016/b978-0-12-385897-9.00001-x . ISBN 978-0-12-385897-9 Полем PMID 21295676 .
- ^ «Паразитизм | Энциклопедия экологии и управления окружающей средой» . Blackwell Science . Получено 8 апреля 2018 года .
- ^ Кайра, JN; Benz, GW; Borucinska, J.; Колер, Н.Е. (1997). «Угоры пугнозы, Simenchelys Parasichidae (Synaphanphandchidae) из сердечного мако, выживаемого оксирисона (Laminidae) » Экологическая биология рыб 49 (1): 139–1 Bibda : 1997evbf..49..139c . Doi : 10.1023/a: 100739860 S2CID 37865366 .
- ^ Лоуренс, По (1981). «Вибрация хозяина - подсказка для местоположения хозяина у паразита, Biosteres longicaudatus ». Oecologia . 48 (2): 249–251. Bibcode : 1981oecol..48..249L . doi : 10.1007/bf00347971 . PMID 28309807 . S2CID 6182657 .
- ^ Cardé, RT (2015). «Интеграция с несколькими кривыми: как женщины-комары находят человека-хозяина» . Текущая биология . 25 (18): R793 - R795. BIBCODE : 2015CBIO ... 25.R793C . doi : 10.1016/j.cub.2015.07.057 . PMID 26394099 .

- ^ Рэндл, CP; Кэннон, Британская Колумбия; Фауст, Ал; и др. (2018). «Подсказки хозяина опосредуют рост и создание дубовой омелы (Phoradendron Leucarpum, Viscaceae), воздушного паразитического растения». Кастанея . 83 (2): 249–262. doi : 10.2179/18-173 . S2CID 92178009 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а Пулин, Роберт ; Рандхава, Хасиб С. (февраль 2015 г.). «Эволюция паразитизма вдоль конвергентных линий: от экологии до геномики» . Паразитология . 142 (Suppl 1): S6 - S15. doi : 10.1017/s0031182013001674 . PMC 4413784 . PMID 24229807 .
- ^ Jump up to: а беременный в дюймовый Lafferty, KD; Курис, А.М. (2002). «Трофические стратегии, разнообразие животных и размер тела» (PDF) . Тенденции в экологии и эволюции . 17 (11): 507–513. doi : 10.1016/s0169-5347 (02) 02615-0 . Архивировано из оригинала (PDF) 3 октября 2019 года.
- ^ Jump up to: а беременный Poulin 2007 , p. 111.
- ^ Elumalai, v.; Вишванатан, C.; Pravinkumar, M.; Раффи, С.М. (2013). «Заражение паразитической саккулины Spp. В коммерческих морских крабах» . Журнал паразитических заболеваний . 38 (3): 337–339. doi : 10.1007/s12639-013-0247-z . PMC 4087306 . PMID 25035598 .
- ^ Ченг, Томас С. (2012). Общая паразитология . Elsevier Science. С. 13–15. ISBN 978-0-323-14010-2 .
- ^ Cox, Fe (2001). «Сопутствующие инфекции, паразиты и иммунные ответы» (PDF) . Паразитология . 122. Дополнение: S23–38. doi : 10.1017/s003118200001698x . PMID 11442193 . S2CID 150432 . Архивировано (PDF) из оригинала 2 декабря 2017 года.
- ^ "Гельминт -паразиты" . Австралийское общество паразитологии . Получено 9 октября 2017 года .
- ^ «Патогенные паразитические инфекции» . Пеоя . Получено 18 июля 2013 года .
- ^ Steere, AC (июль 2001 г.). «Болезнь Лайма». Новая Англия Журнал медицины . 345 (2): 115–125. doi : 10.1056/nejm200107123450207 . PMID 11450660 .
- ^ Jump up to: а беременный Поллитт, Лора С.; МакГрегор, Паула; Мэтьюз, Кит; Рис, Сара Э. (2011). "Передача малярии и трипаносом: разные паразиты, те же правила?" Полем Тенденции в паразитологии . 27 (5): 197–203. doi : 10.1016/j.pt.2011.01.004 . PMC 3087881 . PMID 21345732 .
- ^ Стивенс, Элисон Н.П. (2010). «Хищничество, травоядные и паразитизм» . Знание по природе . 3 (10): 36 . Получено 12 февраля 2018 года .
Хищничество, травоядные и паразитизм существуют вдоль континуума тяжести с точки зрения степени, в которой они негативно влияют на пригодность организма. ... В большинстве ситуаций паразиты не убивают своих хозяев. Исключение, однако, происходит с паразитоидами, которые размывают грань между паразитизмом и хищничеством.
- ^ Jump up to: а беременный в дюймовый Гуллан, PJ; Cranston, PS (2010). Насекомые: схема энтомологии (4 -е изд.). Wiyyeera. Стр. 308 , 365–367, 375, 440–441. ISBN 978-1-118-84615-5 .
- ^ Уилсон, Энтони Дж.; и др. (Март 2017). "Что такое вектор?" Полем Философские транзакции Королевского общества B: биологические науки . 372 (1719): 20160085. DOI : 10.1098/rstb.2016.0085 . PMC 5352812 . PMID 28289253 .
- ^ Jump up to: а беременный Годфри, Стефани С. (декабрь 2013 г.). «Сети и экология передачи паразитов: структура паразитологии дикой природы» . Дикая природа . 2 : 235–245. doi : 10.1016/j.ijppaw.2013.09.001 . PMC 3862525 . PMID 24533342 .
- ^ Jump up to: а беременный Правление, Jetske G.; Робинсон, Айлион; Пауэрс, Стивен Дж.; Гражданские жители, Саския LGE; Колфилд, Джон С.; Биркетс, Майкл А.; Smillion, Renate C.; Gendens, Perry JJ; Bousema, Toun; Sauerwein, Robert W.; Пикетт, Джон А.; Задачи, Уильям; Логан, Джеймс Г. (август 2017 г.). «Запахи или участники Plasmodium falciparum, инфекционные, влияют на взаимодействие комаров с хозяином» . Напаживание отчетов . 7 (1): 9283. Код BIB : 2017Natsr… 7,9283d . doi : 10 1038/S41598-017-08978-9 . PMC 5570919 . PMID 28839251 .
- ^ Jump up to: а беременный Dissanaike, as (1957). «На простейших гиперпаразита в Гельминте, с некоторыми наблюдениями за Nosema helminthorum Moniez, 1887». Журнал гельминтологии . 31 (1–2): 47–64. doi : 10.1017/s0022149x00033290 . PMID 13429025 . S2CID 35487084 .
- ^ Jump up to: а беременный Томас, JA; Schönrogge, K.; Bonelli, S.; Барберо, Ф.; Балетто Э. (2010). «Коррупция акустических сигналов муравья миметическими социальными паразитами: бабочки -макулинея достигают повышенного статуса в обществах -хозяевах, имитируя акустику королевских муравьев» . Commun Integr Biol . 3 (2): 169–171. doi : 10.4161/cib.3.2.10603 . PMC 2889977 . PMID 20585513 .
- ^ Jump up to: а беременный Пейн, Р.Б. (1997). «Птиевой паразитизм выводка» . В Клейтоне, DH; Мур, Дж. (Ред.). Эволюция хозяина - паразита: общие принципы и птичьи модели . Издательство Оксфордского университета. С. 338–369 . ISBN 978-0-19-854892-8 .
- ^ Jump up to: а беременный Слейтер, Питер Дж.Б.; Розенблатт, Джей С.; Сноудон, Чарльз Т.; Roper, Timothy J.; Брокманн, Х. Джейн ; Нагиб, Марк (30 января 2005 г.). Достижения в изучении поведения . Академическая пресса. п. 365. ISBN 978-0-08-049015-1 .
- ^ Jump up to: а беременный Питч, Теодор В. (25 августа 2005 г.). «Дайморфизм, паразитизм и пересмотр пола: способы размножения среди глубоководных рыболовов с цератиоидами (Teleostei: Lophiforifors)». Ихтиологические исследования . 52 (3): 207–236. Bibcode : 2005ichtr..52..207p . doi : 10.1007/s10228-005-0286-2 . S2CID 24768783 .
- ^ Jump up to: а беременный Рочат, Жак; Гутьеррес, Эндрю Пол (май 2001 г.). «Погодная регуляция оливковой шкалы двумя паразитоидами» . Журнал экологии животных . 70 (3): 476–490. Bibcode : 2001 ЯНЕК..70..476R . doi : 10.1046/j.1365-2656.2001.00505.x . S2CID 73607283 .
- ^ Askew, RR (1961). «О биологии жителей дубовых галлов Cynipidae (Hymenoptera) в Британии» . Сделки Общества британской энтомологии . 14 : 237–268.
- ^ Парратт, Стивен Р.; Лейн, Анна-Лиза (январь 2016 г.). «Роль гиперпаразитизма в экологии и эволюции микробного патогена» . Журнал ISME . 10 (8): 1815–1822. Bibcode : 2016ismej..10.1815p . doi : 10.1038/ismej.2015.247 . PMC 5029149 . PMID 26784356 .
- ^ Ван Ойстайен, Аннетт; Арауджо Алвес, Дениз; Калиари Оливейра, Рикардо; Лима до Насименто, Даниэла; Сантос до Насименто, Фабио; Биллен, Йохан; Wenseleers, Tom (сентябрь 2013 г.). «Подлые королевы в Melipona пчел. Селективное обнаружение и проникнуть в колонии без королевы». Поведение животное . 86 (3): 603–609. Citeserx 10.1.1.309.6081 . Doi : 10.1016/j.anbehav.2013.07.001 . S2CID 12921696 .
- ^ «Социальные паразиты в муравейной колонии» . Антежиры . Получено 4 апреля 2016 года .
- ^ Эмери, Карло (1909). «О происхождении дулотических, паразитических и мирмекофильных муравьев». Биологический центральный лист . 29 : 352–362.
- ^ Deslippe, Richard (2010). «Социальный паразитизм у муравьев» . Знание по природе . Получено 29 октября 2010 года .
- ^ Emery, C. (1909). «О происхождении дулотических, паразитических и мирмекофильных муравьев» . Биологический центральный лист . 29 : 352–362.
- ^ Бурк, Эндрю Ф.Г.; Фрэнкс, Найджел Р. (июль 1991 г.). «Альтернативные адаптации, симпатрическое видообразование и эволюция паразитических, инквилиновых муравьев». Биологический журнал Линневого общества . 43 (3): 157–178. doi : 10.1111/j.1095-8312.1991.tb00591.x . ISSN 0024-4066 .
- ^ О'Брайен, Тимоти Г. (1988). «Паразитарное поведение ухода за больными в клиновой обезьяне обезьяны ( Cebus olivaceus )». Американский журнал приматологии . 16 (4): 341–344. doi : 10.1002/ajp.1350160406 . PMID 32079372 . S2CID 86176932 .
- ^ Ротштейн, С.И. (1990). «Модельная система для коэволюции: паразитизм птичьего вывода». Ежегодный обзор экологии и систематики . 21 : 481–508. doi : 10.1146/annurev.ecolsys.21.1.481 .
- ^ Де Марсико, MC; Gloag, R.; Урсино, Калифорния; Rebroeda, JC (март 2013 г.). «Новый метод отказа от паразитических яиц выводка снижает интенсивность паразитизма у хозяина коровьи» . Биологические письма . 9 (3): 20130076. DOI : 10.1098/rsbl.2013.0076 . PMC 3645041 . PMID 23485877 .
- ^ Welbergen, J.; Дэвис, NB (2011). «Паразит в одежде Волка: ястреб -мимикрия уменьшает моббинг кукушек от хозяева» . Поведенческая экология . 22 (3): 574–579. doi : 10.1093/beheco/arr008 .
- ^ Фернесс, RW (1978). «Клептопаразитизм великого скоаса ( Катаракта Скуа Брюнн.) И Арктика Скуаса ( Stercorarius Parasiticus L.) в колонии Shetland Seabird». Поведение животных . 26 : 1167–1177. doi : 10.1016/0003-3472 (78) 90107-0 . S2CID 53155057 .
- ^ Maggenti, Armand R.; Маггенти, Мэри Энн; Гарднер, Скотт Лайелл (2005). Онлайн словарь зоологии беспозвоночных (PDF) . Университет Небраски. п. 22. Архивировано из оригинала (PDF) 18 апреля 2018 года.
- ^ «Избранные существа. Энгарсия одурэкс » . Университет Флориды . Получено 6 января 2018 года .
- ^ Берек, Лук; Schembri, Patrick J.; Букал, Дэвид С. (2005). «Определение пола у Bonellia viridis (Echiura: Boneliidae): динамика и эволюция населения» (PDF ) Оикос 108 (3): 473–4 Bibcode : 2005oikos.108..473b . Doi : 10.1111/j.0030-1299.2005.13350.x . Архивировано (PDF) из оригинала 3 октября
- ^ Роллинсон, Д.; Hay, SI (2011). Достижения в области паразитологии . Оксфорд: Elsevier Science. С. 4–7. ISBN 978-0-12-385897-9 .
- ^ Jump up to: а беременный Poulin 2007 , p. 6
- ^ Поласкек, Эндрю; Vilhemsen, Lars (2023). «Биоразнообразие гименоптерана паразитоидов» . Текущее мнение в науке насекомых . 56 : 101026. Bibcode : 2023cois ... 5601026p . doi : 10.1016/j.cois.2023.101026 . PMID 36966863 . S2CID 257756440 .
- ^ Форбс, Эндрю А.; Бэгли, Робин К.; Пиво, Марк А.; и др. (12 июля 2018 г.). «Количественная оценка не поддающейся инфекции: почему Hymenoptera, а не Coleoptera, является наиболее специфическим порядком животных» . Экология BMC . 18 (1): 21. Bibcode : 2018bmce ... 18 ... 21f . doi : 10.1186/s12898-018-0176-x . ISSN 1472-6785 . PMC 6042248 . PMID 30001194 .
- ^ Моранд, Серж; Краснов, Борис Р.; Литтлвуд, Д. Тимоти Дж. (2015). Разнообразие и диверсификация паразитов . Издательство Кембриджского университета. п. 44. ISBN 978-1-107-03765-6 .
- ^ Rastogi, V. B. (1997). Modern Biology . Pitambar Publishing. p. 115. ISBN 978-81-209-0496-5 .
- ^ Кокла, Анна; Мельник, Чарльз У. (1 октября 2018 г.). «Развитие вора: формация Хаустории на паразитических растениях». Биология развития . 442 (1): 53–59. doi : 10.1016/j.ydbio.2018.06.013 . ISSN 0012-1606 . PMID 29935146 . S2CID 49394142 .
- ^ Jump up to: а беременный в Heide-Jørgensen, Henning S. (2008). Паразитические цветущие растения . Брилль ISBN 978-90-04-16750-6 .
- ^ Никрент, Даниэль Л. (2002). «Паразитические растения мира» (PDF) . Архивировано (PDF) из оригинала 6 марта 2016 года . Получено 10 апреля 2018 года . который появился на испанском языке как глава 2, с. 7–27 в: Ja López-Sáez, P. Catalán и L. Sáez [Eds.], Паразитические растения на Иберийском полуострове и балеарских островах .
- ^ Никрент, DL; Musselman, LJ (2004). «Введение в паразитические цветущие растения». Инструктор по здоровью растений . doi : 10.1094/phi-i-2004-0330-01 .
- ^ Вествуд, Джеймс Х.; Йодер, Джон I.; Тимко, Майкл П.; Depamphilis, Claude W. (2010). «Эволюция паразитизма в растениях». Тенденции в науке о растениях . 15 (4): 227–235. doi : 10.1016/j.tlants.2010.01.004 . PMID 20153240 .
- ^ Лик, младший (1994). «Биология мико-гетеротрофических (« сапрофитных ») растений». Новый фитолог . 127 (2): 171–216. doi : 10.1111/j.1469-8137.1994.tb04272.x . PMID 33874520 . S2CID 85142620 .
- ^ Фэй, Ван; Лю, Йе (11 августа 2022 г.). «Биотрофические грибковые патогены: критический обзор» . Прикладная биохимия и биотехнология . 195 (1): 1–16. doi : 10.1007/s12010-022-04087-0 . ISSN 0273-2289 . PMID 35951248 . S2CID 251474576 .
- ^ "Что такое медовый гриб?" Полем Королевское садоводческое общество . Получено 12 октября 2017 года .
- ^ Чоудхури, Суприё; Басу, Арпита; Кунду, Surekha (8 декабря 2017 г.). «Биотрофия-северная переключатель в патогене вызывает дифференциальный ответ в устойчивом и восприимчивом кунжуте, включающем несколько сигнальных путей на разных фазах» . Научные отчеты . 7 (1): 17251. Bibcode : 2017natsr ... 717251c . doi : 10.1038/s41598-017-17248-7 . ISSN 2045-2322 . PMC 5722813 . PMID 29222513 .
- ^ «Перестань пренебрегать грибами» . Природная микробиология . 2 (8): 17120. 25 июля 2017 г. DOI : 10.1038/nmicrobiol.2017.120 . PMID 28741610 .
- ^ Дидье, эс; Стовалл, я; Green, LC; Бриндли, PJ; Sestak, K.; Дидье, PJ (9 декабря 2004 г.). «Эпидемиология микроспоридиоза: источники и способы передачи» . Ветеринарная паразитология . 126 (1–2): 145–166. doi : 10.1016/j.vetpar.2004.09.006 . PMID 15567583 .
- ^ Esch, KJ; Петерсен, Калифорния (январь 2013 г.). «Передача и эпидемиология зоонозных простейших заболеваний животных -компаньонов» . Клинические обзоры микробиологии . 26 (1): 58–85. doi : 10.1128/cmr.00067-12 . PMC 3553666 . PMID 23297259 .
- ^ McFall-Ngai, Маргарет (январь 2007 г.). «Адаптивный иммунитет: забота о сообществе» . Природа . 445 (7124): 153. Bibcode : 2007natur.445..153m . doi : 10.1038/445153a . PMID 17215830 . S2CID 9273396 .
- ^ Фишер, Брюс; Харви, Ричард П.; Champe, Pamela C. (2007). Иллюстрированные обзоры Липпинкотта: Микробиология (серия иллюстрированных обзоров Lippincott) . Липпинкотт Уильямс и Уилкинс. С. 332–353. ISBN 978-0-7817-8215-9 .
- ^ Кунин, ЭВ; Senkevich, TG; Dolja, VV (2006). «Древний вирусный мир и эволюция клеток» . Биология прямой . 1 : 29. doi : 10.1186/1745-6150-1-29 . PMC 1594570 . PMID 16984643 .
- ^ Брейтбарт, М .; Rohwer, F. (2005). «Здесь вирус, там вирус, везде один и тот же вирус?». Тенденции в микробиологии . 13 (6): 278–284. doi : 10.1016/j.tim.2005.04.003 . PMID 15936660 .
- ^ Лоуренс, CM; Menon, S.; Eilers, BJ; и др. (2009). «Структурные и функциональные исследования архаальных вирусов» . Журнал биологической химии . 284 (19): 12599–603. doi : 10.1074/jbc.r800078200 . PMC 2675988 . PMID 19158076 .
- ^ Эдвардс, Ра; Rohwer, F. (2005). «Вирусная метагеномика» (PDF) . Nature Reviews Microbiology . 3 (6): 504–510. doi : 10.1038/nrmicro1163 . PMID 15886693 . S2CID 8059643 . Архивировано (PDF) из оригинала 3 октября 2019 года.
- ^ Jump up to: а беременный Добсон, А.; Lafferty, KD; Курис, Ам; Hechinger, RF; Jetz, W. (2008). «Дань уважения Линнеусу: сколько паразитов? Сколько хозяев?» Полем Труды Национальной академии наук . 105 (Дополнение 1): 11482–11489. Bibcode : 2008pnas..10511482d . doi : 10.1073/pnas.0803232105 . PMC 2556407 . PMID 18695218 .
- ^ Jump up to: а беременный в Сухдео, Майкл В.К. (2012). "Где паразиты в пищевых сетях?" Полем Паразиты и векторы . 5 (1): 239. doi : 10.1186/1756-3305-5-239 . PMC 3523981 . PMID 23092160 .
- ^ Вольф, Эван Д.С.; Солсбери, Стивен У.; Хорнер, Джон Р.; Варричио, Дэвид Дж. (2009). «Общая птичья инфекция мучила динозавров тирана» . Plos один . 4 (9): E7288. Bibcode : 2009ploso ... 4.7288W . doi : 10.1371/journal.pone.0007288 . PMC 2748709 . PMID 19789646 .
- ^ Ponomarenko, AG (1976) Новое насекомое из мелового периода Трансбайкалии, возможный паразит птерозавров. Палеонтологический журнал 10 (3): 339-343 (английский) / палеонтологически Zhurnal 1976 (3): 102-106 (русский)
- ^ Чжан, Янджи; Ши, Чунгкун; Раснитсин, Александр; Рен, Донг; Гао, Тайпинг (2020). «Новая блоха из раннего мелового китайского языка» . Acta Palaeontologica Polonica . 65 doi : 10.4202/app.00680.2019 .
- ^ Таник Нонсрирак, Серж Моранд, Алексис Рибас, Сита Маниткун, Комсон Лаупразерт, Жюльен Клод (9 августа 2023 г.). «Первое открытие паразитовых яиц в копролите позвоночных позднего триаса в Таиланде» . Plos один . 18 (8): E0287891. Bibcode : 2023ploso..1887891n . doi : 10.1371/journal.pone.0287891 . PMC 10411797 . PMID 37556448 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а беременный РУК, GA (2007). «Гипотеза гигиены и растущая распространенность хронических воспалительных расстройств» . Сделки Королевского общества тропической медицины и гигиены . 101 (11): 1072–1074. doi : 10.1016/j.trstmh.2007.05.014 . PMID 17619029 .
- ^ Jump up to: а беременный в Massey, RC ; Buckling, A.; Ffrench-Constant, R. (2004). «Вмешательство конкуренции и вирулентность паразитов» . Труды Королевского общества B: Биологические науки . 271 (1541): 785–788. doi : 10.1098/rspb.2004.2676 . PMC 1691666 . PMID 15255095 .
- ^ Эвальд, Пол В. (1994). Эволюция инфекционных заболеваний . Издательство Оксфордского университета. п. 8. ISBN 978-0-19-534519-3 .
- ^ Веррен, Джон Х. (февраль 2003 г.). «Вторжение в гендерные изгибы: манипулируя полом и размножением у своих хозяев, многие паразиты улучшают свои собственные шансы на выживание и могут формировать эволюцию самого секса» . Естественная история . 112 (1): 58. OCLC 1759475 . Архивировано из оригинала 8 июля 2012 года . Получено 15 ноября 2008 года .
- ^ Маргулис, Линн ; Саган, Дорион ; Элдредж, Найлс (1995). Что такое жизнь? Полем Саймон и Шустер. ISBN 978-0-684-81087-4 .
- ^ Саркар, Сахотра; Плутинский, Аня (2008). Компаньон философии биологии . Джон Уайли и сыновья. п. 358. ISBN 978-0-470-69584-5 .
- ^ Rigaud, T.; Perrot-Minnot, M.-J.; Браун, MJF (2010). «Сборки паразита и хозяина: принятие реальности улучшит наши знания о передаче паразитов и вирулентности» . Труды Королевского общества B: Биологические науки . 277 (1701): 3693–3702. doi : 10.1098/rspb.2010.1163 . PMC 2992712 . PMID 20667874 .
- ^ Пейдж, Родерик Д.М. (27 января 2006 г.). «Копекс». Энциклопедия наук о жизни . Джон Уайли. doi : 10.1038/npg.els.0004124 . ISBN 978-0-470-01617-6 .
- ^ Switzer, William M.; Салеми, Марко; Шанмугам, Ведапури; и др. (2005). «Древнее совместное обнаружение пенистых вирусов и приматов» . Природа . 434 (7031): 376–380. Bibcode : 2005natur.434..376s . doi : 10.1038/nature03341 . PMID 15772660 . S2CID 4326578 .
- ^ Джонсон, КП; Кеннеди, М.; McCracken, K. G (2006). "Переосмысливая происхождение вшей фламинго: копетки или переключение хозяина?" Полем Биологические письма . 2 (2): 275–278. doi : 10.1098/rsbl.2005.0427 . PMC 1618896 . PMID 17148381 .
- ^ Jump up to: а беременный Живой, CM; Dybdahl, MF (2000). «Адаптация паразитов к местным генотипам хозяина» (PDF) . Природа . 405 (6787): 679–81. Bibcode : 2000natur.405..679L . doi : 10.1038/35015069 . PMID 10864323 . S2CID 4387547 . Архивировал (PDF) из оригинала 7 июня 2016 года.
- ^ Lafferty, KD; Моррис, А.К. (1996). «Измененное поведение паразитированного убийства повышает восприимчивость к хищничеству со стороны конечных хозяев птиц» (PDF) . Экология . 77 (5): 1390–1397. Bibcode : 1996ecol ... 77.1390L . doi : 10.2307/22655536 . JSTOR 2265536 . Архивировано (PDF) из оригинала 3 марта 2019 года.
- ^ Berdoy, M.; Вебстер, JP; MacDonald, DW (2000). «Фатальное притяжение у крыс, инфицированных токсоплазмой Gondii » . Прокурор Биол. Наука 267 (1452): 1591–4. doi : 10.1098/rspb.2000.1182 . PMC 1690701 . PMID 11007336 .
- ^ Кейзо (16 сентября 2019 г.). «Оценка манипулятивных эффектов с помощью личинки с шариком-пауком-пауком (Araneidae: Cyclosa Argenteoalba) посредством хирургического удаления и трансплантации» . Журнал арахнологии 47 2): doi : ( 181. ISSN 0161-8 202579182S2CID
- ^ Александр, Дэвид Э. (2015). На крыле: насекомые, птерозавры, птицы, летучие мыши и эволюция полета животных . Издательство Оксфордского университета. п. 119. ISBN 978-0-19-999679-7 .
- ^ Пулин, Роберт (сентябрь 1995 г.). «Эволюция черт истории жизни паразита: мифы и реальность» (PDF) . Паразитология сегодня . 11 (9): 342–345. doi : 10.1016/0169-4758 (95) 80187-1 . PMID 15275316 . Архивировано из оригинала (PDF) 16 февраля 2012 года.
- ^ Яхалом, Дайана; Аткинсон, Стивен Д.; Нойхоф, Моран; Чанг, Э. Салли; Филипп, Эрве; Картрайт, Полин; Варфоломей, Джерри Л.; Хучон, Дороте (19 февраля 2020 г.). «Книдарный паразит лосося (Myxozoa: Henneguya) не хватает митохондриального генома» . Труды Национальной академии наук . 117 (10): 5358–5363. Bibcode : 2020pnas..117.5358y . doi : 10.1073/pnas.1909907117 . PMC 7071853 . PMID 32094163 .
- ^ Jump up to: а беременный в дюймовый «Взаимодействие хозяина -паразита врожденные защиты хоста» (PDF) . Университет Колорадо. Архивировано из оригинала (PDF) 4 марта 2016 года . Получено 7 мая 2014 года .
- ^ Jump up to: а беременный Maizels, RM (2009). «Иммуномодуляция паразитов и полиморфизмы иммунной системы» . Дж. Биол . 8 (7): 62. doi : 10.1186/jbiol166 . PMC 2736671 . PMID 19664200 .
- ^ Jump up to: а беременный Жанна, Роберт Л. (1979). «Строительство и использование множества расческов в Polistes canadensis в отношении биологии предсказанной мотыльки» . Поведенческая экология и социобиология . 4 (3): 293–310. doi : 10.1007/bf00297649 . S2CID 36132488 .
- ^ Jump up to: а беременный Runyon, JB; Месчер, MC; De Moraes, CM (2010). «Защита растений от паразитических растений демонстрирует сходство с теми, которые вызваны травоядными животными и патогенами» . Поведение сигнала растения . 5 (8): 929–31. Bibcode : 2010plsib ... 5..929r . doi : 10.4161/psb.5.8.11772 . PMC 3115164 . PMID 20495380 .
- ^ Гамильтон, WD ; Axelrod, R.; Tanese, R. (май 1990). «Сексуальное воспроизведение как адаптация для сопротивления паразитам (обзор)» . Труды Национальной академии наук . 87 (9): 3566–3573. Bibcode : 1990pnas ... 87.3566h . doi : 10.1073/pnas.87.9.3566 . PMC 53943 . PMID 2185476 .
- ^ Эберт, Дитер; Гамильтон, Уильям Д. (1996). «Секс против вирулентности: коэволюция паразитических заболеваний». Тенденции в экологии и эволюции . 11 (2): 79–82. doi : 10.1016/0169-5347 (96) 81047-0 . PMID 21237766 .
- ^ Фолстад, Ивар; Картер, Эндрю Джон (1992). «Паразиты, яркие мужчины и иммунокомпетентность» . Американский натуралист . 139 (3): 603–622. doi : 10.1086/285346 . S2CID 85266542 . [ Постоянная мертвая ссылка ]
- ^ Jump up to: а беременный Талер, Дженнифер С.; Карбан, Ричард; Ullman, Diane E.; Бож, Карина; Босток, Ричард М. (2002). «Перекрестный разговор между путями защиты от джассоната и салицилата растительных заводов: воздействие на нескольких паразитов растений» . Oecologia . 131 (2): 227–235. Bibcode : 2002oecol.131..227t . doi : 10.1007/s00442-002-0885-9 . PMID 28547690 . S2CID 25912204 .
- ^ Jump up to: а беременный в Фрэнк, С.А. (2000). «Конкретная и неспецифическая защита от паразитной атаки» (PDF) . J. Theor. Биол . 202 (4): 283–304. Bibcode : 2000jthbi.202..283f . Citeseerx 10.1.1.212.7024 . doi : 10.1006/jtbi.1999.1054 . PMID 10666361 . Архивировано (PDF) из оригинала 14 июня 2001 года.
- ^ Paré, Paul W.; Тумлинсон, Джеймс Х. (1 октября 1999 г.). «Платание растений как защита от травоядных насекомых» . Физиология растений . 121 (2): 325–332. doi : 10.1104/pp.121.2.325 . PMC 1539229 . PMID 10517823 .
- ^ Jump up to: а беременный Poulin 2007 , с. X, 1–2.
- ^ Блейксли, апрель MH; Кеог, Кэролин Л.; Фаулер, Эми Э.; Гриффен, Блейн Д.; Тодд, Питер Алан (1 июня 2015 г.). «Оценка влияния инфекции трематод на инвазивные зеленые крабы в восточной части Северной Америки» . Plos один . 10 (6): E0128674. BIBCODE : 2015PLOSO..1028674B . doi : 10.1371/journal.pone.0128674 . PMC 4451766 . PMID 26030816 .
- ^ Эберхард, ML (август 2016 г.). «Возможная роль рыбы и лягушек как паратенических хозяев Dracunculus medinensis, Чад» . Возникающие инфекционные заболевания . 22 (8): 1428–1430. doi : 10.3201/eid2208.160043 . PMC 4982183 . PMID 27434418 .
- ^ Холт, Р.Д. (2010). «SOAPBOX IJEE: Мир, свободный от паразитов и векторов: будет ли это небеса, или это будет ад?» (PDF) . Израильский журнал экологии и эволюции . 56 (3): 239–250. doi : 10.1560/ijee.56.3-4.239 . Архивировано (PDF) из оригинала 8 сентября 2015 года.
- ^ Combes, Claude (2005). Искусство быть паразитом . Университет Чикагской Прессы. ISBN 978-0-226-11438-5 .
- ^ Хадсон, Питер Дж.; Добсон, Эндрю П.; Лафферти, Кевин Д. (2006). "Является ли здоровая экосистема, которая богата паразитами?" (PDF) . Тенденции в экологии и эволюции . 21 (7): 381–385. doi : 10.1016/j.tree.2006.04.007 . PMID 16713014 . Архивировано из оригинала (PDF) 10 августа 2017 года.
- ^ Стрингер, Эндрю Пол; Linklater, Wayne (2014). «Все в меру: принципы контроля паразитов для сохранения дикой природы» . Биоссака . 64 (10): 932–937. doi : 10.1093/biosci/biu135 .
- ^ Лафферти, Кевин Д.; Аллезина, Стефано; Арим, Матиас; Бриггс, Чери Дж.; и др. (2008). «Паразиты в пищевых сетях: окончательные недостающие ссылки» . Экологические письма . 11 (6): 533–546. Bibcode : 2008ecoll..11..533l . doi : 10.1111/j.1461-0248.2008.01174.x . PMC 2408649 . PMID 18462196 .
- ^ Чейз, Джонатан (2013). «Паразиты в пищевых сетях: распутывание запутанного банка» . PLOS Биология . 11 (6): E1001580. doi : 10.1371/journal.pbio.1001580 . PMC 3678997 . PMID 23776405 .
- ^ Rózsa, L.; Reiczigel, J.; Majoros, G. (2000). «Количественная оценка паразитов в образцах хозяев» (PDF) . J. Parasitol . 86 (2): 228–32. doi : 10.1645/0022-3395 (2000) 086 [0228: qpisoh] 2.0.co; 2 . PMID 10780537 . S2CID 16228008 . Архивировано из оригинала (PDF) 19 июня 2018 года.
- ^ Jump up to: а беременный в дюймовый и фон Кокс, Фрэнсис Эг (июнь 2004 г.). «История человеческих паразитических заболеваний». Инфекционные болезни Клиники Северной Америки . 18 (2): 173–174. doi : 10.1016/j.idc.2004.01.001 . PMID 15145374 .
- ^ Jump up to: а беременный в Ченг, Томас С. (1973). Общая паразитология . Академическая пресса. С. 120–134. ISBN 978-0-12-170750-7 Полем
19 -й век можно рассматривать как генезис современной паразитологии.
- ^ Хамфри-Смит, Ян, изд. (1993). Семь веков паразитологии во Франции [ Французская школа паразитологии ] (на французском языке). Французское паразитологическое общество. стр. 26–29.
- ^ Jump up to: а беременный Иоли, а.; Петитори, JC; Theodorides, J. (1997). «Франческо Реди и рождение экспериментальной паразитологии». Hist Sci Med . 31 (1): 61–66. PMID 11625103 .
- ^ Буш, Ао; Фернандес, JC; ESCH, GW; SEED, JR (2001). Паразитизм: разнообразие и экология паразитов животных . Издательство Кембриджского университета. п. 4 ISBN 978-0-521-66447-9 .
- ^ «Акарус как причина кабины» . Университет Кальяри . Получено 11 апреля 2018 года .
- ^ «Паразитология» . Мерриам-Уэбстер . Получено 13 апреля 2018 года .
- ^ Эллис, Гарольд (март 2006 г.). «Сэр Дэвид Брюс, пионер тропической медицины». Британский журнал больничной медицины . 67 (3): 158. doi : 10.12968/hmed.2006.67.3.20624 . PMID 16562450 .
- ^ «Кандидаты против малярии и малярии» . Колледж врачей Филадельфии. 19 апреля 2017 года . Получено 11 февраля 2018 года .
- ^ Уолш, Фергус (24 июля 2015 г.). «Вакцина против малярии получает« зеленый свет » » . Би -би -си . Получено 25 июля 2015 года .
- ^ Ходдл, MS; Ван Дризше, RG; Сандерсон, JP (1998). «Биология и использование паразитоидной пилотидной лайтформы». Ежегодный обзор энтомологии . 43 : 645–669. doi : 10.1146/annurev.ento.43.1.645 . PMID 15012401 .
- ^ «Паразитоидные осы (Hymenoptera)» . Университет Мэриленда. Архивировано с оригинала 27 августа 2016 года . Получено 6 июня 2016 года .
- ^ Обнаружение инноваций в развитии биопестицидов. Архивировано 15 мая 2012 года в Европейской комиссии «Уэйкк Машины» (2008). Доступ 9 января 2017 года
- ^ Poulin 2007 , с. 265–266.
- ^ Matyszak, Philip (2017). 24 часа в Древнем Риме: день из жизни людей, которые там жили . Майкл О'Мара. п. 252. ISBN 978-1-78243-857-1 .
- ^ Дэймон, Синтия (1997). "5" . Маска паразита: патология римского покровительства . Университет Мичиганской прессы. п. 148. ISBN 978-0-472-10760-5 Полем
Сатирик, стремящийся изобразить страдания клиента, естественно, фокусируется на отношениях с наибольшей зависимостью, в которой клиент получает свою еду от своего покровителя, и для этого изготовленная персонация паразита оказалась чрезвычайно полезной.
- ^ Jump up to: а беременный Playfair, Джон (2007). Жизнь с микробами: в отношении здоровья и болезней . Издательство Оксфордского университета. п. 19. ISBN 978-0-19-157934-9 Полем Playfair сравнивает популярное использование с точки зрения биолога о паразитизме, который он называет (возглавляя ту же страницу) «древний и респектабельный взгляд на жизнь».
- ^ Свифт, Джонатан (1733). По поэзии: рапсодия . И продается Дж. Хуггонсоном, рядом с кофейной домом Кента, недалеко от Сержант-Инна, на канцелярии; [и] в книжных и брошетке.
- ^ Пулин, Роберт ; Макдугалл, Кэмерон; Прессвелл, Бронвен (11 мая 2022 г.). «Что в названии? Таксономические и гендерные смещения в этимологии названий новых видов» . Труды Королевского общества B: Биологические науки . 289 (1974). doi : 10.1098/rspb.2021.2708 . PMC 9091844 . PMID 35538778 .
- ^ Отис, Лаура (2001). Сеть: общение с телами и машинами в девятнадцатом веке . Университет Мичиганской прессы. п. 216. ISBN 978-0-472-11213-5 .
- ^ «Паразитизм и симбиоз» . Энциклопедия научной фантастики . 10 января 2016 года.
- ^ Dove, Alistair (9 мая 2011 г.). «Это явно важный вид, с которым мы имеем дело» . Deep Sea News.
- ^ Паппас, Стефани (29 мая 2012 г.). «5 инопланетных паразитов и их реальные коллеги» . Живая наука.
- ^ Серсель, Алекс (19 мая 2017 г.). «Паразитизм в инопланетных фильмах» . Журнал Signal to Noise.
- ^ Нордия, Майкл (25 апреля 2017 г.). « Эволюция« Чужой »: исследуйте каждый этап в ужасном жизненном цикле ксеноморфа. Отпразднуйте День инопланетян с взглядом на прошлое, настоящее и будущее самого ужасающего инопланетного кинематографа» . IndieWire .
Источники
[ редактировать ]- Пулин, Роберт (2007). Эволюционная экология паразитов . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА . ISBN 978-0-691-12085-0 .
Дальнейшее чтение
[ редактировать ]- Combes, Claude (2005). Искусство быть паразитом . Университет Чикагской Прессы . ISBN 978-0-226-11438-5 .
- Desowitz, Robert (1998). Кто дал Пинту Санта -Мария? Полем Книги урожая. ISBN 978-0-15-600585-2 .
- Циммер, Карл (2001). Паразит Рекс . Свободная пресса . ISBN 978-0-7432-0011-0 .
Внешние ссылки
[ редактировать ] Паразитические насекомые, клещи и клещи: роды медицинской и ветеринарной важности в Wikibooks
Паразитические насекомые, клещи и клещи: роды медицинской и ветеринарной важности в Wikibooks - Университет Аберистуита: паразитология - Класс с ссылками на полные текстовые статьи о паразитизме и паразитологии.
- Разделение паразитических заболеваний , Центры по контролю и профилактике заболеваний
- KSU: паразитологические исследования - ставки и ссылки на паразитологии
- Паразитологические ресурсы на всемирной паутине: мощный инструмент для практиков инфекционных заболеваний (издательство Оксфордского университета)
