Шистосома
Эта статья нуждается в дополнительных цитатах для проверки . ( июль 2016 г. ) |
| Шистосома | |
|---|---|
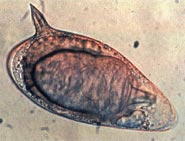 | |
| шистосомы мансони Яйцо | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Платихельминты |
| Сорт: | Трематоды |
| Заказ: | Диплостомида |
| Семья: | Шистосоматиды |
| Подсемейство: | Шистосоматинае |
| Род: | Шистосома Винная страна , 1858 год. |
| Разновидность | |
Шистосома бомфорди | |
Шистосома — род трематод , широко известных как кровяные сосальщики . Это паразитические плоские черви, ответственные за весьма значительную группу инфекций у людей , называемых шистосомозом , который, по мнению Всемирной организации здравоохранения, является вторым наиболее разрушительным в социально-экономическом отношении паразитарным заболеванием (после малярии ), которым заражены сотни миллионов человек во всем мире. [1] [2]
Взрослые плоские черви паразитируют в кровеносных капиллярах брыжейки или сплетения мочевого пузыря, в зависимости от вида заражения. Они уникальны среди трематод и любых других плоских червей тем, что раздельнополы с отчетливым половым диморфизмом между самцом и самкой . Тысячи яиц высвобождаются и достигают либо мочевого пузыря, либо кишечника (в зависимости от вида заражения), а затем выводятся с мочой или калом в пресную воду . Затем личинки должны пройти через промежуточного хозяина- улитку , прежде чем появится следующая личиночная стадия паразита, которая может заразить нового хозяина-млекопитающего, непосредственно проникая в кожу.
Эволюция
[ редактировать ]
Происхождение этого рода остается неясным. Многие годы считалось, что этот род имеет африканское происхождение, однако секвенирование ДНК позволяет предположить, что виды ( S. edwardiense и S. hippopotami ), поражающие гиппопотамов ( Hippopotamus amphibius ), могли быть базальными. Поскольку бегемоты обитали как в Африке, так и в Азии в кайнозойскую эпоху, этот род мог возникнуть как паразиты бегемотов. [3] Первоначальными хозяевами видов Юго-Восточной Азии, вероятно, были грызуны . [4]
Судя по филогенетике улиток-хозяев, вполне вероятно, что этот род возник в Гондване между 70 миллионами и 120 лет назад . [5]
Родственной группой шистосомы шистосом , является род заражающих слонов , — Bivitellobilharzia .
, крупного рогатого скота овец , коз и кашемировых коз Паразит Orientobilharzia turkestanicum, по-видимому, связан с африканскими шистосомами. [6] [7] Этот последний вид с тех пор был переведен в род Schistosoma . [8]
Внутри haematobium группы S. bovis и S. curassoni , по-видимому, тесно связаны, как и S. leiperi и S. mattheei . [ нужна ссылка ]
S. mansoni , по-видимому, возник в Восточной Африке 0,43–0,30 миллиона лет назад. [ нужна ссылка ]
S. mansoni и S.rodhaini , по-видимому, имели общего предка между 107,5 и 147,6 тысячами лет назад. [9] Этот период совпадает с самыми ранними археологическими свидетельствами рыболовства в Африке. Похоже, что S. mansoni возникла в Восточной Африке и испытала сокращение эффективной численности популяции 20-90 тысяч лет назад, прежде чем рассеяться по континенту во время голоцена . Позже этот вид был завезен в Америку в результате работорговли.
S. incognitum и S. nasale более тесно связаны с африканскими видами, чем с группой japonicum . [ нужна ссылка ]
S. sinensium , по-видимому, распространился во время плиоцена . [10] [11]
S. mekongi , по-видимому, вторгся в Юго-Восточную Азию в середине плейстоцена. [4]
Предполагаемые даты видообразования группы japonicum : ~3,8 миллиона лет назад для S. japonicum /Shistosoma Юго-Восточной Азии и ~2,5 миллиона лет назад для S. malayensis / S. mekongi . [4]
Schistosoma turkestanicum обнаружена у благородного оленя в Венгрии . Эти штаммы, похоже, отличаются от штаммов, обнаруженных в Китае и Иране . [12] Дата расхождения, по-видимому, насчитывает 270 000 лет до настоящего времени.
Таксономия
[ редактировать ]Род Schistosoma в настоящее время [ когда? ] определяется как парафилетический , [ нужна ссылка ] так что изменения вероятны. В пределах этого рода известно более двадцати видов.
Род был разделен. [ нужна ссылка ] на четыре группы: indicum , japonicum , haematobium и mansoni . Родство остальных видов все еще выясняется.
Тринадцать видов обитают в Африке. Двенадцать из них разделены на две группы: с боковым шипом на яйце ( группа мансони ) и с терминальным шипом ( группа гематобиум ).
Группа Мансони
[ редактировать ]Четыре вида группы мансони : S. edwardiense , S. hippotami , S. mansoni и S.rodhaini .
Группа гематобиумов
[ редактировать ]Девять видов группы haematobium : S. bovis , S. curassoni , S. guineensis , S. haematobium , S. intercalatum , S. kisumuensis , S. leiperi , S. margrebowiei и S. mattheei .
S. leiperi и S. matthei , по-видимому, связаны между собой. [13] S. margrebowiei является базальным в этой группе. [14] S. guineensis является родственным видом групп S. bovis и S. curassoni . S. intercalatum на самом деле может представлять собой видовой комплекс, состоящий как минимум из двух видов. [15] [16]
Индийская группа
[ редактировать ]Группа Indicum включает три вида: S. indicum , S. nasale и S. spindale . Эта группа, по-видимому, возникла в плейстоцене. Все используют легочных улиток в качестве хозяев. [17] S. spindale широко распространен в Азии, но встречается и в Африке. [ нужна ссылка ] Они встречаются в Азии и Индии . [18]
S. indicum встречается в Индии и Таиланде . [ нужна ссылка ]
Группа индикаторов, по-видимому, является сестринской кладой африканских видов. [19]
Группа Японикум
[ редактировать ]Группа japonicum включает пять видов: S. japonicum , S. malayensis и S. mekongi , S. ovuncatum и S. sinensium , эти виды встречаются в Китае и Юго-Восточной Азии. [20]
S. ovuncatum образует кладу с S. sinensium и встречается в северном Таиланде. Окончательный хозяин неизвестен, промежуточным хозяином является улитка Tricula bollingi . Известно, что этот вид использует в качестве хозяев улиток семейства Pomatiopsidae . [20]
S. incognitum, по-видимому, является базальным в этом роде. Возможно, он более тесно связан с афро-индийскими видами, чем с группой Юго-Восточной Азии. В качестве хозяев этот вид использует легочных улиток. [ нужна ссылка ] Исследование митохондрий позволяет предположить, что Schistosoma incognitum может представлять собой видовой комплекс. [21]
Новые виды
[ редактировать ]По состоянию на 2012 год к этому роду были отнесены еще четыре вида. [8] ранее классифицированный как вид рода Orientobilharzia . Orientobilharzia отличается от Schistosoma морфологически только количеством семенников. Обзор морфологических и молекулярных данных показал, что различия между этими родами слишком малы, чтобы оправдать их разделение. Четыре вида являются
- Шистосома бомфорди
- Шистосома датта
- Шистосома харинасутай
- Шистосома туркестаникум
Гибриды
[ редактировать ]Гибрид S. haematobium-S.guineenis наблюдался в Камеруне в 1996 г. S. haematobium смог прижиться только после вырубки тропических лесов в Луме рядом с эндемичным S. guineensis ; гибридизация привела к конкурентному исключению S. guineensis . [22]
В 2003 г. S. mansoni-S. Гибрид родахайни был обнаружен у улиток в западной Кении . [23] По состоянию на 2009 год он не был обнаружен у людей. [24]
В 2009 г. S. haematobium–S. Гибриды bovis были описаны у детей северного Сенегала. Бассейн реки Сенегал сильно изменился с 1980-х годов после плотин Диама в Сенегале и плотины Манантали строительства в Мали. Плотина Диама препятствовала проникновению океанской воды и открыла новые формы сельского хозяйства. миграция людей, увеличение поголовья домашнего скота и мест, где люди и крупный рогатый скот загрязняют воду, способствовали смешиванию различных шистосом в Н'Дере . Например, [24] Тот же гибрид был идентифицирован во время расследования вспышки шистосомоза на Корсике в 2015 году , произошедшей от реки Каву . [25]
В 2019 г. S. haematobium–S. mansoni Hybrid был описан у 14-летнего пациента с гематурией из Кот-д'Ивуара . [26]
Кладограмма
[ редактировать ]Кладограмма , основанная на генах 18S рибосомальной РНК , 28S рибосомальной РНК и частичной субъединицы I цитохром с оксидазы (COI), показывает филогенные отношения видов рода Schistosoma : [27]
| |||||||||||||||||||
Сравнение яиц
[ редактировать ]
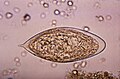

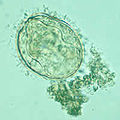
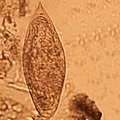
Географическое распространение
[ редактировать ]Географические территории, связанные с шистосомозом по данным Всемирной организации здравоохранения по состоянию на январь 2017 года, включают в алфавитном порядке: Африка, Бразилия, Камбоджа, Карибский бассейн, Китай, Корсика, Индонезия, Лаос, Ближний Восток, Филиппины, Суринам и Венесуэла. [28] В Европе не было случаев заболевания с 1965 года, пока вспышка не произошла на Корсике. [25]
Шистосомоз
[ редактировать ]Паразитические плоские черви Schistosoma вызывают группу хронических инфекций, называемых шистосомозом, известным также как бильгарциоз. [29] Противошистосомальный препарат — шистосомицид .
Виды, заражающие человека
[ редактировать ]Паразитизм людей, вызываемый шистосомами, по-видимому, возникал по крайней мере трижды как в Азии , так и в Африке .
- S. guineensis , недавно описанный вид, встречается в Западной Африке . К известным улиток промежуточным хозяевам относятся Bulinus forskalii .
- S. haematobium , обычно называемый пузырным сосальщиком , первоначально обнаруженный в Африке, на Ближнем Востоке и в Средиземноморском бассейне, был завезен в Индию во время Второй мировой войны. Пресноводные улитки рода Bulinus являются важным промежуточным хозяином этого паразита. Среди окончательных хозяев наибольшее значение имеют люди. Другими окончательными хозяевами редко являются павианы и обезьяны. [30]
- С. интеркалатум . Обычными окончательными хозяевами являются люди. Других животных можно заразить экспериментально. [30]
- S. japonicum , общее название которого просто кровяная двуустка , широко распространен в Восточной Азии и юго-западной части Тихого океана . Пресноводные улитки рода Oncomelania являются важным промежуточным хозяином S. japonicum. Окончательными хозяевами являются люди и другие млекопитающие, включая кошек, собак, коз, лошадей, свиней, крыс и водяных буйволов. [30]
- S. malayensis. Этот вид, по-видимому, является редкой инфекцией человека и считается зоонозом. [ нужна ссылка ] . Естественным позвоночным хозяином является гигантская крыса Мюллера Сунда ( Sundamys muelleri ). Улитками-хозяевами являются виды Robertsiella ( R. gismanni , R. kaporensis и R. silvicola (см. Attwood et al., 2005 Journal of Molluscan Studies, том 71, выпуск 4, стр. 379–391).
- S. mansoni встречается в Африке , Бразилии , Венесуэле , Суринаме , на Малых Антильских островах , в Пуэрто-Рико и Доминиканской Республике . Он также известен как кровяная двуустка Мэнсона или болотная лихорадка . Пресноводные улитки рода Biomphalaria являются важным промежуточным хозяином этой трематоды. Среди окончательных хозяев наибольшее значение имеют люди. Другими окончательными хозяевами являются бабуины, грызуны и еноты. [30]
- S. mekongi родственен S. japonicum и поражает как верхние, так и нижние брыжеечные вены. S. mekongi отличается тем, что у него яйца меньшего размера, другой промежуточный хозяин ( Neotricula aperta ) и более длительный препатентный период у хозяина-млекопитающего. Окончательными хозяевами являются люди и собаки. [30] Улитка Tricula aperta также может быть экспериментально заражена этим видом. [ нужна ссылка ]
| Научное название | Первый промежуточный хост | Эндемичная зона |
|---|---|---|
| Гвинейская шистосома | Булинус форскалии | Западная Африка |
| Вставочная шистосома | Булинус виды | Африка |
| Шистосома гематобиум | Булинус виды. | Африка , Ближний Восток |
| Шистосома японская | Онкомелания виды. | Китай , Восточная Азия , Филиппины |
| Шистосома малайская | Робертсиелла виды. | Юго-Восточная Азия |
| Шистосома Мансони | Биомфалярии виды. | Африка , Южная Америка , Карибский бассейн , Средний Восток |
| Шистосома меконги | Открытая неотрикула | Юго-Восточная Азия |
Виды, заражающие других животных
[ редактировать ]Schistosoma indicum , Schistosoma nasale , Schistosoma spindale , Schistosoma leiperi являются паразитами жвачных животных . [ нужна ссылка ]
Schistosoma edwardiense и Schistosoma бегемоты являются паразитами бегемотов. [ нужна ссылка ]
Schistosoma ovuncatum и Schistosoma sinensium являются паразитами грызунов. [ нужна ссылка ]
Морфология
[ редактировать ]Взрослые шистосомы разделяют все основные черты дигенеи. Они имеют основную двустороннюю симметрию , ротовую и брюшную присоски, тело покрыто синцитиальной покрышкой , слепую пищеварительную систему, состоящую из рта , пищевода и раздвоенных слепых отростков ; область между тегументом и пищеварительным каналом, заполненная рыхлой сетью мезодермы клеток , и экскреторной или осморегуляторной системой, основанной на пламенных клетках . Взрослые черви, как правило, имеют длину 10–20 мм (0,39–0,79 дюйма) и используют глобины своих хозяев из гемоглобина для своей собственной системы кровообращения.
Воспроизведение
[ редактировать ]
В отличие от других трематод и практически всех других плоских червей шистосомы раздельнополы , т . е . раздельнополы. Оба пола демонстрируют сильную степень полового диморфизма , причем самец значительно крупнее самки. Самец окружает самку и заключает ее в своем гинакофорном канале на всю взрослую жизнь червей. Питаясь кровью хозяина, самец передает часть ее самке. Самец также передает химические вещества, которые завершают развитие самки, после чего они размножаются половым путем. Хотя и редко, но иногда спарившиеся шистосомы «разводятся», при этом самка покидает самца ради другого самца. Точная причина неизвестна, хотя считается, что самки оставят своих партнеров, чтобы спариваться с более генетически отдаленными самцами. Такой биологический механизм будет способствовать уменьшению инбридинга и может быть фактором необычно высокого генетического разнообразия шистосом. [31]
Геном
[ редактировать ]о геномах Schistosoma haematobium , S. japonicum и S. mansoni . Сообщалось [32] [33] [34] [35]
История
[ редактировать ]Яйца этих паразитов впервые увидел Теодор Максимилиан Бильгарц , немецкий патологоанатом, работавший в Египте в 1851 году, который обнаружил яйца Schistosoma haematobium во время вскрытия . он написал своему бывшему учителю фон Зибольду В мае и августе 1851 года два письма с описанием своих открытий. Фон Зибольд опубликовал в 1852 году статью, в которой резюмировал открытия Бильгарца и назвал червей Distoma haematobium . [36] В 1856 году Билхарц написал статью, более подробно описывающую червей . [37] Их необычная морфология означала, что их нельзя было с комфортом включить в состав Дистомы . Так в 1856 году Меккель фон Хельмсбек ( де ) создал для них род Bilharzia . [38] В 1858 году Дэвид Фридрих Вейнланд предложил название Schistosoma (по-гречески: «разделенное тело»), потому что черви не были гермафродитами, а имели разные полы. [39] Несмотря на то, что Bilharzia имеет приоритет, рода название Schistosoma было официально принято Международной комиссией по зоологической номенклатуре . Термин Bilharzia для обозначения заражения этими паразитами до сих пор используется в медицинских кругах. [ нужна ссылка ]
Билхарц также описал Schistosoma mansoni , но этот вид был переописан Луи Вестенрой Самбоном в 1907 году в Лондонской школе тропической медицины , который назвал его в честь своего учителя Патрика Мэнсона . [40]
поместили все известные на тот момент виды в подсемейство В 1898 году Стайлз и Хассель в 1899 году возвел его в семейный статус. . Лоосс Поче в 1907 году исправил грамматическую ошибку в фамилии. Жизненный цикл Schistosoma mansoni определил бразильский паразитолог Пираха да Силва (1873-1961) в 1908 году. [41]
геномы Schistosoma mansoni и Schistosoma japonicum . В 2009 году были расшифрованы [32] [33] открывая путь к новым таргетным методам лечения. В частности, исследование показало, что геном S. mansoni содержит 11 809 генов , многие из которых производят ферменты для расщепления белков , что позволяет паразиту проникать сквозь ткани. Кроме того, у S. mansoni нет фермента для производства определенных жиров , поэтому для их производства он должен полагаться на своего хозяина. [42]
Уход
[ редактировать ]Ссылки
[ редактировать ]- ^ «Информационный бюллетень о шистосомозе» . Всемирная организация здравоохранения . Проверено 10 августа 2011 г.
- ^ «Шистосомоз» . Центры по контролю и профилактике заболеваний . Проверено 10 августа 2011 г.
- ^ Морган Дж.А., ДеДжонг Р.Дж., Казибве Ф., Мкоджи Г.М., Локер Э.С. (август 2003 г.). «Недавно выявленная линия шистосом». Международный журнал паразитологии . 33 (9): 977–85. дои : 10.1016/S0020-7519(03)00132-2 . ПМИД 12906881 .
- ^ Перейти обратно: а б с Эттвуд С.В., Фатих Ф.А., Упатам Э.С. (март 2008 г.). «Вариации последовательности ДНК среди популяций Schistosoma mekongi и родственных таксонов; филогеография и современное распространение азиатского шистосомоза» . PLOS Забытые тропические болезни . 2 (3): е200. дои : 10.1371/journal.pntd.0000200 . ПМК 2265426 . ПМИД 18350111 .
- ^ Beer SA, Voronin MV, Zazornova OP, Khrisanfova GG, Semenova SK (2010). "[Phylogenetic relationships among schistosomatidae]". Meditsinskaia Parazitologiia I Parazitarnye Bolezni (in Russian) (2): 53–9. PMID 20608188 .
- ^ Ван Ч.Р., Ли Л., Ни Х.Б., Чжай Ю.К., Чен А.Х., Чен Дж., Чжу XQ (февраль 2009 г.). «Orientobilharzia turkestanicum является представителем рода Schistosoma на основании филогенетического анализа с использованием последовательностей рибосомальной ДНК». Экспериментальная паразитология . 121 (2): 193–7. дои : 10.1016/j.exppara.2008.10.012 . ПМИД 19014940 .
- ^ Ван Ю, Ван ЧР, Чжао Г.Х., Гао Дж.Ф., Ли М.В., Чжу XQ (декабрь 2011 г.). «Полный митохондриальный геном Orientobilharzia turkestanicum подтверждает его родство с африканскими видами Schistosoma». Инфекция, генетика и эволюция . 11 (8): 1964–70. дои : 10.1016/j.meegid.2011.08.030 . ПМИД 21930247 .
- ^ Перейти обратно: а б Олдхун Дж. А., Литтлвуд DT (июнь 2012 г.). «Orientobilharzia Dutt & Srivastava, 1955 (Trematoda: Schistosomatidae), младший синоним Schistosoma Weinland, 1858». Систематическая паразитология . 82 (2): 81–8. дои : 10.1007/s11230-012-9349-8 . ПМИД 22581244 . S2CID 18890027 .
- ^ Креллен Т., Аллан Ф., Дэвид С., Даррант С., Хаквейл Т., Холройд Н., Эмери А.М., Роллинсон Д., Аненсен Д.М., Берриман М., Вебстер Дж.П., Коттон Дж.А. (февраль 2016 г.). «Полное секвенирование генома человеческого паразита Schistosoma mansoni раскрывает историю популяции и эффекты отбора» . Научные отчеты . 6 : 20954. Бибкод : 2016NatSR...620954C . дои : 10.1038/srep20954 . ПМЦ 4754680 . ПМИД 26879532 .
- ^ Эттвуд SW, Упатам ES, Мэн XH, Цю округ Колумбия, Саутгейт VR (август 2002 г.). «Филогеография азиатских шистосом (Trematoda: Schistosomatidae)». Паразитология . 125 (Часть 2): 99–112. дои : 10.1017/s0031182002001981 . ПМИД 12211613 . S2CID 40281441 .
- ^ Эттвуд С.В., Ибараки М., Сайто Ю., Нихей Н., Дженис Д.А. (2015). «Сравнительные филогенетические исследования Schistosoma japonicum и ее промежуточного хозяина улитки Oncomelania hupensis: происхождение, распространение и коэволюция» . PLOS Забытые тропические болезни . 9 (7): e0003935. дои : 10.1371/journal.pntd.0003935 . ПМК 4521948 . ПМИД 26230619 .
- ^ Лоутон С.П., Майорос Дж. (март 2013 г.). «Иностранный захватчик или местный затворник? Штрих-кодирование ДНК показывает отчетливую европейскую линию зоонозного паразита Schistosoma turkestanicum (син. Orientobilharzia turkestanicum ())» . Инфекция, генетика и эволюция . 14 : 186–93. дои : 10.1016/j.meegid.2012.11.013 . ПМИД 23220360 .
- ^ Каукас А., Диас Нето Е., Симпсон А.Дж., Саутгейт В.Р., Роллинсон Д. (апрель 1994 г.). «Филогенетический анализ видов группы Schistosoma haematobium на основе случайно амплифицированной полиморфной ДНК». Международный журнал паразитологии . 24 (2): 285–90. дои : 10.1016/0020-7519(94)90040-x . ПМИД 8026909 .
- ^ Вебстер Б.Л., Саутгейт В.Р., Литтлвуд Д.Т. (июль 2006 г.). «Пересмотр взаимоотношений Schistosoma, включая недавно описанную Schistosoma guineensis». Международный журнал паразитологии . 36 (8): 947–55. дои : 10.1016/j.ijpara.2006.03.005 . ПМИД 16730013 .
- ^ Кейн Р.А., Саутгейт В.Р., Роллинсон Д., Литтлвуд Д.Т., Локьер А.Э., Пажес-младший, Чуем Тчуэнте Л.А., Журдан Дж. (август 2003 г.). «Филогения, основанная на трех митохондриальных генах, поддерживает разделение Schistosoma intercalatum на два отдельных вида». Паразитология . 127 (Часть 2): 131–7. дои : 10.1017/s0031182003003421 . ПМИД 12954014 . S2CID 23973239 .
- ^ Пажес-младший, Дюран П., Саутгейт В.Р., Тчуэм Тчуэнте Л.А., Журдан Дж. (январь 2001 г.). «Молекулярные аргументы в пользу разделения Schistosoma intercalatum на два отдельных вида». Паразитологические исследования . 87 (1): 57–62. дои : 10.1007/s004360000301 . ПМИД 11199850 . S2CID 11121161 .
- ^ Лю Л., Мондал М.М., Идрис М.А., Локман Х.С., Раджапаксе П.Дж., Сатриха Ф., Диас Дж.Л., Упатам Э.С., Эттвуд С.В. (июль 2010 г.). «Филогеография Indoplanorbis exustus (Gastropoda: Planorbidae) в Азии » Паразиты и переносчики 3:57 . дои : 10.1186/1756-3305-3-57 . ПМЦ 2914737 . ПМИД 20602771 .
- ^ Эттвуд С.В., Фатих Ф.А., Мондал М.М., Алим М.А., Фаджар С., Раджапаксе Р.П., Роллинсон Д. (декабрь 2007 г.). «Исследование группы Schistosoma indicum (Trematoda: Digenea) на основе последовательностей ДНК: популяционная филогения, таксономия и историческая биогеография». Паразитология . 134 (Часть 14): 2009–20. дои : 10.1017/S0031182007003411 . ПМИД 17822572 . S2CID 22737354 .
- ^ Агацума Т., Ивагами М., Лю С.Х., Раджапаксе Р.П., Мондал М.М., Китикун В., Амбу С., Агацума Ю., Блер Д., Хигучи Т. (март 2002 г.). «Сродство между азиатскими видами шистосом, не относящимися к человеку, группой S. indicum и африканскими шистосомами человека». Журнал гельминтологии . 76 (1):7–19. дои : 10.1079/joh200191 . hdl : 10126/3484 . ПМИД 12018199 . S2CID 25582541 .
- ^ Перейти обратно: а б Эттвуд С.В., Панасопонкул С., Упатам Э.С., Мэн XH, Саутгейт В.Р. (январь 2002 г.). «Schistosoma ovuncatum n. sp. (Digenea: Schistosomatidae) из северо-западного Таиланда и историческая биогеография Schistosoma Weinland в Юго-Восточной Азии, 1858». Систематическая паразитология . 51 (1): 1–19. дои : 10.1023/а:1012988516995 . ПМИД 11721191 . S2CID 21696073 .
- ^ Вебстер Б.Л., Литтлвуд Д.Т. (2012)Изменение порядка митохондриальных генов у шистосом (Platyhelminthes: Digenea: Schistosomatidae). Int J Parasitol 42(3):313-321
- ^ Чуэм Тчуэнте Л.А., Саутгейт В.Р., Ньиоку Ф., Нджине Т., Куемени Л.Е., Журдан Дж. (1997). «Эволюция шистосомоза в Луме, Камерун: замена Schistosoma intercalatum S. haematobium посредством интрогрессивной гибридизации». Труды Королевского общества тропической медицины и гигиены . 91 (6): 664–5. дои : 10.1016/s0035-9203(97)90513-7 . ПМИД 9509173 .
- ^ Морган Дж.А., ДеДжонг Р.Дж., Лвамбо Нью-Джерси, Мунгаи Б.Н., Мкоджи ГМ, Локер Э.С. (апрель 2003 г.). «Первое сообщение о естественном гибриде Schistosoma mansoni и S.rodhaini». Журнал паразитологии . 89 (2): 416–8. doi : 10.1645/0022-3395(2003)089[0416:FROANH]2.0.CO;2 . ПМИД 12760671 . S2CID 948644 .
- ^ Перейти обратно: а б Хейс Т., Вебстер Б.Л., Гелдоф С., Стотард Дж.Р., Диау О.Т., Полман К., Роллинсон Д. (сентябрь 2009 г.). «Двунаправленная интрогрессивная гибридизация шистосом крупного рогатого скота и человека» . ПЛОС Патогены . 5 (9): e1000571. дои : 10.1371/journal.ppat.1000571 . ПМЦ 2731855 . ПМИД 19730700 .
- ^ Перейти обратно: а б Буасье Дж., Греч-Анджелини С., Вебстер Б.Л., Аллиен Дж.Ф., Хьюз Т., Мас-Кома С. и др. (август 2016 г.). «Вспышка урогенитального шистосомоза на Корсике (Франция): эпидемиологический пример» (PDF) . «Ланцет». Инфекционные болезни . 16 (8): 971–9. дои : 10.1016/S1473-3099(16)00175-4 . ПМИД 27197551 . S2CID 3725312 .
- ^ Депакит Дж., Ахунди М., Хаушин Д., Мантелет С., Изри А. (2019). «Нет предела межвидовой гибридизации шистосом: наблюдение из истории болезни» . Паразит . 26:10 . doi : 10.1051/parasite/2019010 . ПМК 6396650 . ПМИД 30821247 .

- ^ Брант С.В., Морган Дж.А., Мкоджи Г.М., Снайдер С.Д., Раджапаксе Р.П., Локер Э.С. (февраль 2006 г.). «Подход к выявлению жизненных циклов, таксономии и разнообразия кровяных двуусток: предоставление ключевых справочных данных, включая последовательность ДНК на отдельных стадиях жизненного цикла» . Журнал паразитологии . 92 (1): 77–88. дои : 10.1645/GE-3515.1 . ПМК 2519025 . ПМИД 16629320 .
- ^ Информационный бюллетень ВОЗ
- ^ Краткая энциклопедия Britannica, 2007 г.
- ^ Перейти обратно: а б с д и Мэнсон-Бар П.Е., Белл Д.Р., ред. (1987). Тропические болезни Мэнсона . Лондон: Байер Тиндалл. ISBN 978-0-7020-1187-0 .
- ^ «Даже кровяные сосальщики разводятся — ткацкий станок» . Ткацкий станок . 08.10.2008 . Проверено 24 мая 2016 г.
- ^ Перейти обратно: а б Консорциум функционального анализа секвенирования генома Schistosoma japonicum; и др. (июль 2009 г.). «Геном Schistosoma japonicum раскрывает особенности взаимодействия хозяина и паразита» . Природа . 460 (7253): 345–51. Бибкод : 2009Natur.460..345Z . дои : 10.1038/nature08140 . ПМЦ 3747554 . ПМИД 19606140 .
- ^ Перейти обратно: а б Берриман М., Хаас Б.Дж., ЛоВерде П.Т., Уилсон Р.А., Диллон Г.П., Серкейра Г.К. и др. (июль 2009 г.). «Геном кровяного двуустки Schistosoma mansoni» . Природа . 460 (7253): 352–8. Бибкод : 2009Natur.460..352B . дои : 10.1038/nature08160 . ПМЦ 2756445 . ПМИД 19606141 .
- ^ Янг Н.Д., Джекс А.Р., Ли Б., Лю С., Ян Л., Сюн З. и др. (январь 2012 г.). «Полногеномная последовательность Schistosoma haematobium» . Природная генетика . 44 (2): 221–5. дои : 10.1038/ng.1065 . hdl : 10072/45821 . ПМИД 22246508 . S2CID 13309839 .
- ^ Протасио А.В., Цай И.Дж., Бэббидж А., Никол С., Хант М., Аслетт М.А. и др. (январь 2012 г.). Хоффманн К.Ф. (ред.). «Систематически улучшенный высококачественный геном и транскриптом человеческой двуустки Schistosoma mansoni» . PLOS Забытые тропические болезни . 6 (1): e1455. дои : 10.1371/journal.pntd.0001455 . ПМЦ 3254664 . ПМИД 22253936 .
- ^ Бильгарц Т., фон Зибольд КТ (1852 г.). «Вклад в литературу о гельминтах, [поражающих] человека…». Журнал научной зоологии (на немецком языке). 4 :53–76. См.: «2. Distomum Haematobium Bilh. », стр. 59–62.
- ^ Билхарц Т. (1856 г.). «Distomum haematobium и его связь с некоторыми патологическими изменениями мочевых органов человека » . Венский медицинский еженедельник (на немецком языке). 6 : 49–52, 65–68.
- ^ Хемсбах Дж. Х., Бильрот Т. (1856 г.). Микрогеология: О конкрециях в животных организмах [ Микрогеология: О конкрециях в животных организмах ] (на немецком языке). Берлин, (Германия): Георг Раймер. п. 114. ИСБН 9783112028636 . Из стр. 114: «Бильгарц впервые описал в книге Зибольда и Кёлликера Zeitschr. f. Zoologie 1852. нового человеческого висцерального червя, очень похожего на дистомов и поэтому названного им Distomum haematobium . Название вида очень описательное, название рода Distoma должно быть не останется и должен быть заменен Билгарзией». (Бильгарц впервые описал в журнале фон Зибольда и Кёлликера по [научной] зоологии 1852 года нового кишечного червя человека, [который] очень похож на Distoma и поэтому был назван им Distomum haematobium . Видовое название очень характерно; название рода не может по праву оставаться Distoma [оно] должно быть заменено на Bilharzia.)
- ^ Вейнланд Д.Ф. (1858 г.). Человеческие цестоиды: очерк о ленточных червях человека ... Кембридж, Массачусетс, США: Меткалф и компания. п. 87. См. сноску †.
- ^ См.:
- Самбон LW (1 апреля 1907 г.). «Новые или малоизвестные африканские энтозоа» . Журнал тропической медицины и гигиены . 10 : 117.
- Самбон LW (16 сентября 1907 г.). «Заметки о Schistosoma mansoni» . Журнал тропической медицины и гигиены . 10 : 303–304.
- ^ См.:
- да Силва П. (август 1908 г.). «Вклад в изучение шистосомоза в Баии». Бразилия-Медико (на португальском языке). 22 :281–282.
- да Силва П. (декабрь 1908 г.). «Вклад в изучение шистосомоза в Баии. Шестнадцать наблюдений» [Вклад в изучение шистосомоза в Баии. Шестнадцать наблюдений.]. Бразилия-Медико (на португальском языке). 22 : 441–444.
- да Силва П (1908). «Вклад в изучение шистосомоза. Двадцать наблюдений» [Вклад в изучение шистосомоза в Баии. Двадцать наблюдений.]. Бразилия-Медико (на португальском языке). 22 : 451–454.
- да Силва П (1908). «Шистосомоз в Баии». Архивы паразитологии (на французском языке). 13 : 283–302.
- да Силва П. (1909). «Вклад в изучение шистосомоза в Баии, Бразилия» . Журнал тропической медицины и гигиены . 12 : 159–164.
- ^ «Расшифрованы гены паразитов-убийц» . Новости Би-би-си . 16 июля 2009 года . Проверено 16 июля 2009 г.
- ^ «Паразиты – шистосомоз» . Центры по контролю и профилактике заболеваний . Министерство здравоохранения и социальных служб США. 28 октября 2020 г.
Дальнейшее чтение
[ редактировать ]- Росс А.Г., Сани А.С., Ли Ю, Дэвис ГМ, Уильямс ГМ, Цзян З и др. (апрель 2001 г.). «Шистосомоз в Китайской Народной Республике: перспективы и проблемы XXI века» . Обзоры клинической микробиологии . 14 (2): 270–95. doi : 10.1128/CMR.14.2.270-295.2001 . ПМЦ 88974 . ПМИД 11292639 .
Внешние ссылки
[ редактировать ]- Британский департамент международного контроля над шистосомозом
- Страница Всемирной организации здравоохранения о шистосомозе
- Лаборатория шистосом Кембриджского университета
- Обзор паразитов шистосом, биология, изображение жизненного цикла на MetaPathogen
| Базы данных органов управления : Национальные |
|---|